| I discovered this having lived in Cape Town for about five years at the time. | Я обнаружила это, когда прожила в Кейптауне около пяти лет. |
| I remember the house we lived in in Ilorin, in Stadium Road. | Я помню дом, в котором мы жили в Ллорине, на Стадиум Роад. |
| I have lived in and around St Louis my entire life. | Я провёл в Сент-Луисе и его окрестностях всю свою жизнь. |
| I had lived on earth long enough to know what the sky looks like. | Я прожил на Земле достаточно, чтобы знать, как выглядит небо. |
| He was pansexual, where Madison didn’t speak to a woman expect for once until he was 42 years old, and then married Dolley and lived happily ever after for 40 years. | Он был пансексуалом, а Мэдисон в жизни и с женщиной-то не говорил до тех пор, пока ему не исполнилось 42 года, и тогда он женился на Долли и жил с ней долго и счастливо следующие 40 лет. |
| So we can look at those to tell us what lived and grew on the landscape and to understand how these rivers were interacting with their floodplains. | Эта информация позволит нам узнать об обитателях и растениях тех времён, а также понять, как реки вели себя в поймах. |
| In seventh grade, I had a best friend named Jenny who lived on the Southwest Side of Chicago. | В седьмом классе у меня была лучшая подруга, её звали Дженни, она жила в районе Саутвест, Чикаго. |
| And then if they wanted to call my grandparents who lived in another city, they needed to book something called a trunk call, and then wait again, for hours or even days. | И если они хотели позвонить родственникам в другой город, нужно было заказать междугородние переговоры и затем ждать часами или даже днями. |
She lived in an isolated community in the forest that had gone for 100 years without any health care until last year when a nurse trained her neighbors to become community health workers. | Она жила в изолированной лесной общине, которая сотни лет обходилась без медицинской помощи, пока в прошлом году медсестра не обучила членов общины, дав им статус медработников. |
| He lived in an area where the roads were sparse and there had been massive shortages of health workers. | Он жил в местности, где почти не было дорог и не хватало медицинских работников. |
| And that is they added a gene that makes the insect glow under UV light so that when they released it they could follow exactly how far it went how long it lived and all of the kinds of data for a good scientific study. | Это был ген, который заставляет насекомое светиться под лучами ультрафиолета, и когда они выпустили его, можно было отслеживать, куда оно полетело, сколько прожило, и получать всевозможные данные для научного исследования. |
I lived in denial for a long time, and the denial was maybe because of the things that had happened, dropping out of school and being fired from my job.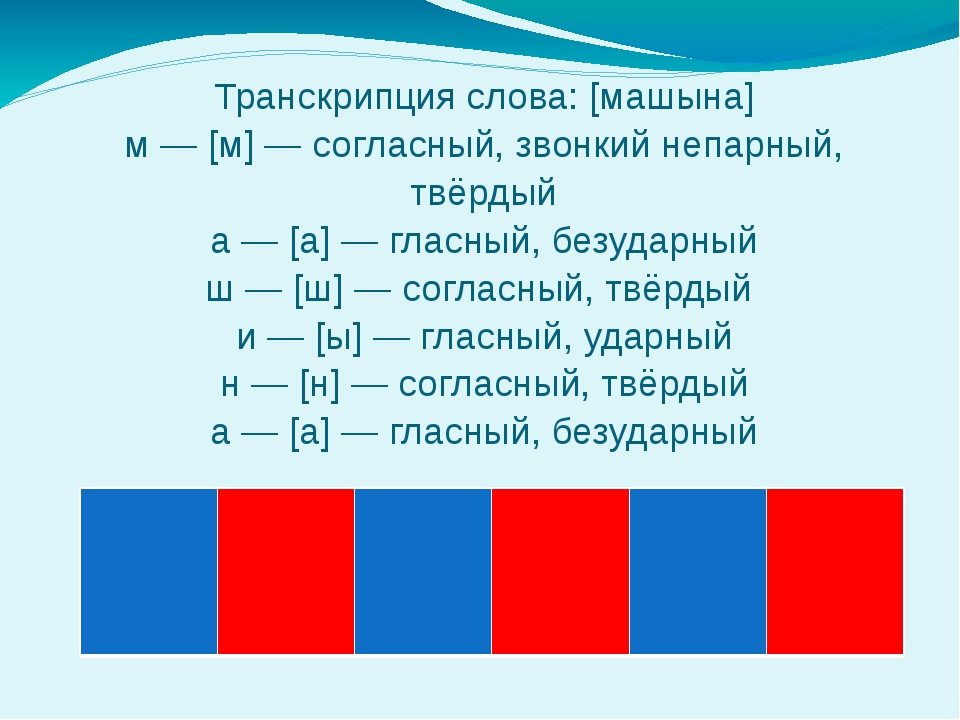 | В течение длительного времени я не хотела с этим смиряться, и, возможно, причиной тому было то, что со мной произошло — исключение из университета и увольнение с работы. |
| George Orwell, one of the greatest writers who ever lived, experienced poverty firsthand in the 1920s. | Джордж Оруэлл, один из величайших писателей всех времён, узнал о нищете на собственном опыте в 1920-х. |
| I believe in a future where the point of education is not to prepare you for another useless job but for a life well-lived. | Я верю в будущее, где цель образования — подготовить вас не к очередной бесполезной работе, а к тому, чтобы вы хорошо прожили жизнь. |
| The most underreported fact of Harriet Tubman is that she lived to be 93 years old, and she didn’t live just an ordinary life; uh-uh. | Наименее известный факт о Гарриет Табмен — это то, что она дожила до 93 лет, и нет, это не была обычная жизнь. |
| We lived with Paul’s illness for 22 months. | Мы прожили с болезнью Пола около 22 месяцев. |
| It’s entirely possible that many of the whales swimming in the Arctic today, especially long-lived species like the bowhead whale that the Inuits say can live two human lives — it’s possible that these whales were alive in 1956, when Jacques Cousteau made his film. | Вполне возможно, что многие из плавающих сегодня в Арктике китов, особенно долгоживущих видов, таких как гренландский кит, которые, как говорят инуиты, могут прожить две человеческие жизни, возможно, что эти киты были живы в 1956 году, когда Жак Кусто снимал свой фильм. |
| I was raised in a home where the two traditions lived together more or less in harmony. | Я вырос в доме, где две традиции жили вместе довольно гармонично, стоит сказать. |
She grew up wishing they lived on a farm, so given the opportunity to adopt local farm animals, she enthusiastically brings them in. | В детстве она мечтала жить на ферме, и когда представилась возможность, она с радостью приютила нескольких животных. |
| The people who lived in the myriads of towns and villages and cities on the Eastern front, who watched their neighbors be rounded up — men, women, children, young, old — and be marched to the outskirts of the town to be shot and left dead in ditches. | Люди, жившие во многих городах, деревнях, местечках на Восточном фронте, те, кто видел, как их соседей — мужчин и женщин, молодых и стариков — собирали и выводили на окраину города, где расстреливали и бросали в канаву. |
| Or the Poles, who lived in towns and villages around the death camps, who watched day after day as the trains went in filled with people and came out empty. | Ошибаются поляки, жившие по соседству с концлагерями и ежедневно видевшие, как переполненные людьми поезда прибывали и возвращались пустыми. |
He lowered himself, he lived his entire human existence practicing the real, concrete language of love. | Который спустился и прожил целую человеческую жизнь, следуя языку любви. |
| He lived in the time of the Soviet Union. | Он жил в Советском Союзе. |
| Okoloma lived on my street and looked after me like a big brother. | Мы с Околомой жили на одной улице, он присматривал за мной как старший брат. |
| So in the literal way, men rule the world, and this made sense a thousand years ago because human beings lived then in a world in which physical strength was the most important attribute for survival. | Так что мужчины буквально правят миром, и это имело смысл тысячу лет назад, потому что люди жили в мире, где физическая сила была необходима для выживания. |
| We lived in a very liberal town. | Мы жили в очень либеральном городе. |
My data sample was actually pretty limited and skewed, so probably it was because of that, I thought, I lived in a bubble, and I didn’t have enough chances to see outside of it. | Доступные мне данные были достаточно ограниченными и искажёнными, и я подумала, что, возможно, поэтому я жила как будто в пузыре, без возможности выглянуть наружу. |
| It looked like we had enough information this time, and many more chances to see outside the closed circle we lived in. | Казалось, будто тогда у нас было достаточно информации и намного больше возможностей посмотреть вне замкнутого пространства, где мы жили. |
| Matter of fact, if we lived in the screen world, we would have a population crisis on our hands. | По сути, если бы мы жили в мире с экрана, то переживали бы демографический кризис. |
| If you go back to the first nations, so, thousands of years ago, the people who lived along the Yellow River in China — it was many, many different tribes and they all depended on the river for survival and for prosperity. | Посмотрим на первые народы, жившие, скажем, тысячи лет назад вдоль реки Хуанхэ в Китае, — то было множество разных племён, существование и процветание которых зависели от реки. |
| But for millions of years, Homo sapiens and our hominid ancestors lived in small communities of a few dozen individuals. | Но миллионы лет Homo sapiens и наши гоминидные предки жили небольшими сообществами, насчитывающими несколько десятков человек. |
| She had lived through bombs being dropped in her town. | Она пережила падающие на город бомбы. |
| She had lived through explosions. | Она пережила взрывы. |
| So they lived on Mars time for a month and had these great adventures, like going bowling in the middle of the night or going to the beach. | То есть целый марсианский месяц был насыщен приключениями, как то: игра в боулинг среди ночи или отдых на пляже. |
| They thought 56 out of every 100 Japanese people lived in rural areas. | Они думали, что 56 % японцев живёт в сельской местности. |
However, I believe that the most important thing we do as archaeologists is acknowledge that past people existed and lived lives worth learning about. | Я считаю, что самое главное, в работе археолога — это признание существования людей и их жизни, о которой нам стоит узнать. |
| Lisht is a royal site, there would have been thousands of people buried there who lived and worked at the court of Pharaoh. | Лишт — территория царей, там были захоронены тысячи людей, которые жили и работали при дворе фараона. |
| Tell her the truth, how you lived in spite of crooked odds. | Скажи ей правду, как выжила, несмотря на жалкие шансы. |
| In fact, everyone at school knew the block where we lived, because it was where people came to buy weed and other drugs. | На самом деле в школе каждый знал квартал, в котором мы жили, именно туда люди приходили, чтобы купить траву и другие наркотики. |
| She worried that the violence we witnessed would overtake our lives, that our poverty meant that the neighbors with whom we lived and shared space would harm us. | Она боялась, что насилие, которое мы видели, захлестнёт нашу жизнь, что наша нищета означала, что соседи, с которыми мы жили и делили пространство, нас обидят. |
| We’ve lived in cities for a very long time, but most of us didn’t live in cities. | Мы уже очень долго живём в городах, но большинство из нас живут не в городе. |
| In 1980, 80 percent of China’s population lived below the extreme poverty level, below the level of having $1,90 per person per day. | В 1980 году 80 % населения Китая жило ниже уровня крайней бедности, получая в день меньше, чем 1,9 доллара на человека. |
| By the year 2000, only 20 percent of China’s population lived below the extreme poverty level — a remarkable feat, admittedly, with some costs in civil liberties that would be tough to accept in the Western world. | К 2000 году только 20 % населения Китая жило ниже уровня крайней бедности — выдающееся достижение, стоившее им, однако, ряда гражданских свобод, что было бы весьма трудно принять в западном мире. |
Well, our ancestors lived immersed in conflict, and they fought right where they lived.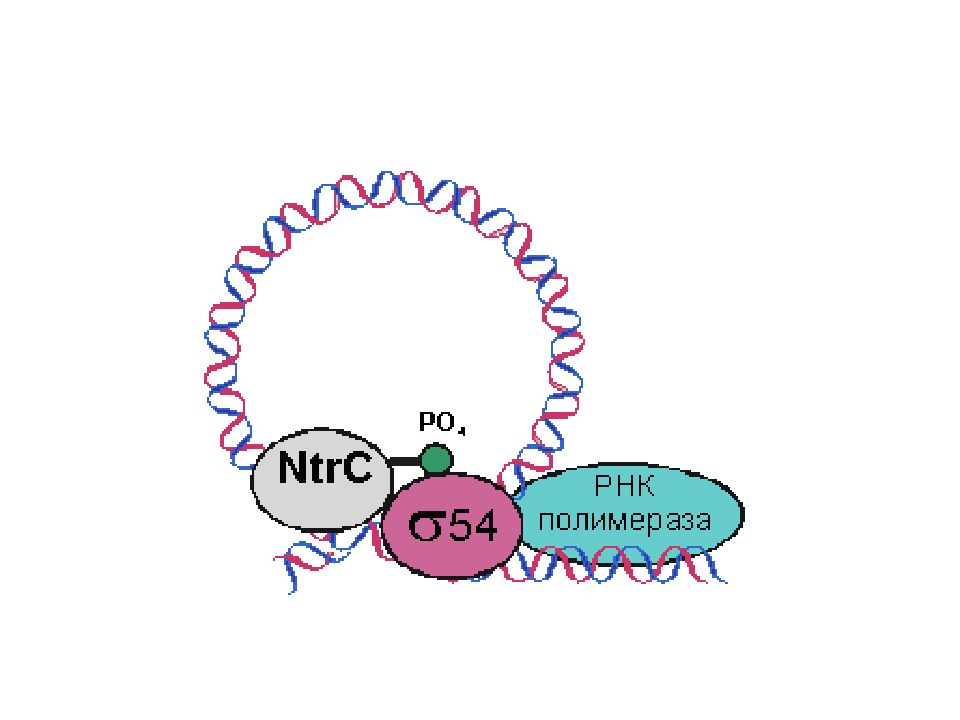 | Наши предки жили в постоянных конфликтах и боролись там же, где жили. |
| Because of this lack of wealth, we lived in a neighborhood that lacked wealth, and henceforth a school system that lacked wealth. | Не имея много денег, мы жили в небогатом районе и я училась в небогатой школе. |
| Nothing to Lose is a work made in collaboration with performers of size who drew from their lived experiences to create a work as varied and authentic as we all are. | Нечего терять создано при сотрудничестве актёров, использовавших свой жизненный опыт, чтобы создать произведение с теми же разнообразием и достоверностью, что есть в нас. |
| But for a long time, I lived for death. | Но долгое время я жил во имя смерти. |
| Why did he die and I lived? | Почему он умер, а я продолжал жить? |
| Everyone lived happily ever after, well, not so fast. | Казалось бы, что сказочке конец, но не тут-то было. |
| So say I lived in a village with the first five rows of this audience, and we all knew one another, and say I wanted to borrow money. | Допустим, я живу в одной деревне с пятью первыми рядами этой аудитории, мы все друг друга знаем, и я хочу занять деньги. |
| He lived out the remainder of his days isolated and tight-lipped. | Остаток своих дней он провёл в молчании и уединении. |
| Pismire lived in a shack near the old wood quarry. | Писмайр жил в хижине поблизости от старого деревянного карьера. |
| What a strange creature lived in that regrown flesh. | Что же за странное создание живет в этом регенерированном теле! |
| Ishaq knew which officials lived in the nice homes. | Он отлично знал чиновников, живших в тех великолепных домах. |
| If I am not mistaken, you’ve lived in Russia for two years. | Если я не ошибаюсь, вы живете в России уже два года. |
| Tricky question, because I’ve lived in a lot of places. | Сложный вопрос, поскольку я жил в разных местах. |
| We lived there, My parents lived there for about ten years, but then, the last couple of that I was, well, I spent some of it in a college in Utah, some of it in Prague. | Мы прожили там, Мои родители прожили там примерно десять лет, но затем, последние два года я был, ну, какое-то время я провел в колледже в Юте, какое-то время — в Праге. |
| And I lived last in Boston, and I’m gonna go back to Washington to work, when I return home. | А я в последнее время жил в Бостоне, и когда вернусь домой, собираюсь поехать работать в Вашингтон. |
| And then they came over, and lived in Idaho and Utah back, when that was totally, basically, breaking new soil, I mean, it was very little there at that time. | Затем они вернулись жить в Айдахо и Юту в то время, когда там, фактически, все только зарождалось, я имею в виду то, что в то время там мало чего было. |
| True, they lived in another epoch, from 1860s to 1890s, I believe. | Правда, они жили в другую эпоху, кажется, в 1860-1890-е годы. |
| Другие результаты | |
Срочно!!!Транскрипция предложения »Жили-были старик со старухой»
СРОЧНО СОР ПО РУССКОМУ ЯЗЫКУ ПОЖАЛУЙСТА
какие различия вы видите в том, как происходит изучение иностранного и русского языка? ПОЛНЫЙ ДОКЛАД!!!
Кто пишит стихи тоже дайте свой дискорд я хочу объменяться стихами и почитать их
В меня много кто не верил, и не верит до сих пор… главное стараться и верить в себя и свои силы! где надо поставить запятые?
В меня много кто не верил и не верит до сих пор…НО главное стараться и верить в себя и свои силы! где надо ставить запятые??? помогите пожалуйста
Помагите пажалуста! Мы привыкли считать, что человек – венец творения. Стоя на верхней ступени эволюции, он приспособился использовать при
… родные ресурсы для своих целей, и вот пещерный охотник, который недавно ставил капкан для мамонта, уже исследует космос. Но чем шире размах – тем больше требуется ресурсов. Со временем человечество стало поручать рутинную и тяжелую работу компьютерным алгоритмам. Сегодня применение роботов в современном мире уже никого не удивляет.На плечи механических друзей ложится множество разнообразных задач. Медицина, банковское обслуживание, промышленность, даже развлечения – основные области применения роботов. Однако с каждым годом появляется все больше работы, которая по зубам искусственному интеллекту. Например,космороботы.B) С чем ассоциируется у вас понятие о робототехнике? Согласитесь, воображение рисует нечто, человекоподобное, с механическими руками и ногами, либо, паукообразное, а ещё, обязательно представляется знаменитая собака-робот. Одним словом, представление о роботах у многих достаточно узкое и однобокое.На самом деле, в современном мире, роботы- довольно востребованы. Их используют в абсолютно различных сферах жизни, о которых многие могут даже не догадываться.Здравоохранение — одна из самых прогрессивных сфер, в которой применяется труд роботов.
Но чем шире размах – тем больше требуется ресурсов. Со временем человечество стало поручать рутинную и тяжелую работу компьютерным алгоритмам. Сегодня применение роботов в современном мире уже никого не удивляет.На плечи механических друзей ложится множество разнообразных задач. Медицина, банковское обслуживание, промышленность, даже развлечения – основные области применения роботов. Однако с каждым годом появляется все больше работы, которая по зубам искусственному интеллекту. Например,космороботы.B) С чем ассоциируется у вас понятие о робототехнике? Согласитесь, воображение рисует нечто, человекоподобное, с механическими руками и ногами, либо, паукообразное, а ещё, обязательно представляется знаменитая собака-робот. Одним словом, представление о роботах у многих достаточно узкое и однобокое.На самом деле, в современном мире, роботы- довольно востребованы. Их используют в абсолютно различных сферах жизни, о которых многие могут даже не догадываться.Здравоохранение — одна из самых прогрессивных сфер, в которой применяется труд роботов. В настоящее время активно развивается роботизированная хирургия. Благодаря кибернетическим технологиям человек может вернуть утраченную часть тела.В медицине достигнут большой прорыв с тех пор, как стали использоваться бионические протезы, которыми человек может управлять при помощи собственной нервной системы.1.Укажите общие и отличительн признаки содержания текста А и текста В, перефразируя предложения.Общие признаки:Отличительные признаки:
В настоящее время активно развивается роботизированная хирургия. Благодаря кибернетическим технологиям человек может вернуть утраченную часть тела.В медицине достигнут большой прорыв с тех пор, как стали использоваться бионические протезы, которыми человек может управлять при помощи собственной нервной системы.1.Укажите общие и отличительн признаки содержания текста А и текста В, перефразируя предложения.Общие признаки:Отличительные признаки:
ПОМОГИТЕ ОЧЕНЬ НАДО
.303. Прочитайте соотношения идеальной фигуры человека, которые установил древнегрееский скульптор Поликлет. Проверьте соотношения на себе. Насколько
… древние нормыприемлемы сегодня?Расстояние от локтя до кончиков пальцев составляет одну пятую роста, от локтядо подмышечной ямкиОдну восьмую роста, от верхней части груди до макушкисоставляет одну шестую, а до корней волос —одну седьмую роста. Наибольшая ши-Восьмая часть роста.Длина лица равна одной десятой длины тела, а длина стопы одной седьмойРасстояние от носа до кончика подбородка и от бровей до корней волос одинаково исоставляет одну треть лица, такой же является длина уха.
Русский язык 1 урок смотрим ролик, пишем конспект в тетради , высылаем мне фото https://youtu.be/ZYKPu2E1_XM
.303. Прочитайте соотношения идеальной фигуры человека, которые установил древнегрееский скульптор Поликлет. Проверьте соотношения на себе. Насколько … древние нормыприемлемы сегодня?Расстояние от локтя до кончиков пальцев составляет одну пятую роста, от локтядо подмышечной ямкиОдну восьмую роста, от верхней части груди до макушкисоставляет одну шестую, а до корней волос —одну седьмую роста. Наибольшая ши-Восьмая часть роста.Длина лица равна одной десятой длины тела, а длина стопы одной седьмойРасстояние от носа до кончика подбородка и от бровей до корней волос одинаково исоставляет одну треть лица, такой же является длина уха.
Lived: перевод, значение, синонимы, произношение, транскрипция, антонимы, примеры предложений | Английско-русский переводчик онлайн
Sergeant told me that the dog lived with Nelson.
Сержант сказал мне, что собака жила с Нельсоном.
We lived in peace, and then you came and laid waste to everything and everyone we loved.
Мы жили в мире, а потом пришел ты и опустошил все и всех, кого мы любили.I lived in Japan for 2 years, but now I’m living in California.
Я прожил в Японии 2 года, а сейчас живу в Калифорнии.December: The Greatest Man Who Ever Lived.
Декабрь: Самый великий человек, который когда-либо жил.See, my cousin Loc Dog lived right across the street.
Видите ли, мой двоюродный брат Лок Дог жил прямо через дорогу.He probably felt as did Lot, another righteous man who lived after the Flood.
Вероятно, он чувствовал то же, что и лот, еще один праведник, живший после потопа.These were the 11 sons that were born to Jacob when he lived with Rachel’s father Laʹban.
Это были 11 сыновей, которые родились у Иакова, когда он жил с отцом Рахили Лаваном. One such law had to do with the disposal of human waste, which had to be properly buried away from the camp so that the area where people lived would not be polluted.
So the monks, map-makers of the Middle Ages, saw the world they lived in.”.
Так монахи, составители карт средневековья, увидели мир, в котором жили”.He only who has lived with the beautiful can die beautifully.
Только тот, кто жил с прекрасным, может умереть красиво.I didn’t know you used to do that when you lived in Australia.
Я не знал, что ты это делал, когда жил в Австралии.While she had lived many years with her husband, they had only the traditional consensual marriage.
Хотя она прожила много лет со своим мужем, у них был только традиционный брак по обоюдному согласию.A Christian father with four teen-agers lived in an area apparently void of recreational opportunities.
Отец-христианин с четырьмя подростками жил в районе, по-видимому, лишенном возможностей для отдыха.
Tom told me he thought that Mary still lived in Boston.
Том сказал мне, что, по его мнению, Мэри все еще живет в Бостоне.I totally lived up to the Joey name.
Я полностью соответствовал имени Джоуи.Prior to the Flood, many humans lived for centuries.
До Потопа многие люди жили веками.Tom is the one who told Mary where John lived.
Том сказал Мэри, где живет Джон.Tom told me he thought that Mary still lived in Australia.
Том сказал мне, что он думал, что Мэри все еще живет в Австралии.Long ago the four nations lived together in harmony.
Давным-давно четыре народа жили вместе в гармонии.Civilians where I lived really had to learn how to protect themselves.
Гражданские там, где я жил, действительно должны были научиться защищать себя. Τhe victims names have yet to be released but they were believed to have lived here, Cendrillon Ηouse.
He has lived in Kabylie, Algeria, so he also speaks Berber.
Все говорят на берберском в Кабиле, Алжир.What are we, indeed, what is our character, if not the concentration of the things which we have lived since our birth.
В самом деле, что мы такое, каков наш характер, если не сосредоточение всего того, чем мы жили с самого рождения?For the vast majority of human history, pretty much everyone lived on the equivalent of one dollar per day, and not much changed.
На протяжении большей части человеческой истории почти все жили на сумму, эквивалентную одному доллару в день, и мало что изменилось.In 1956, I left Bethel to marry Etty, a pioneer I had become acquainted with when she came from the Netherlands to visit her sister who then lived in London.
В 1956 году я покинул Бетель, чтобы жениться на Этти, пионерке, с которой я познакомился, когда она приехала из Нидерландов навестить свою сестру, которая тогда жила в Лондоне.
We lived close to a cemetery, and whenever a funeral procession approached our house, I woke up my child and held it close until the procession had passed.
Мы жили недалеко от кладбища, и всякий раз, когда похоронная процессия приближалась к нашему дому, я будила своего ребенка и прижимала его к себе, пока процессия не проходила мимо.For the first few years of my life, my family lived near Leipzig, East Germany, not far from the Czech and Polish borders.
Первые несколько лет моей жизни моя семья жила недалеко от Лейпцига, Восточная Германия, недалеко от чешской и польской границ.He lived in a typical Japanese-style house.
Он жил в типичном доме в японском стиле.Any joy and happiness would be short-lived if the dead came back to an earth that was filled with strife, bloodshed, pollution, and violence—as is the situation today.
Любая радость и счастье были бы недолговечны, если бы мертвые вернулись на землю, которая была наполнена борьбой, кровопролитием, загрязнением и насилием-как это происходит сегодня.On the east side of the city lived a retired army major who harbored a general dislike for Jehovah’s Witnesses, although, much to his dismay, his own son was a Witness.
В восточной части города жил отставной армейский майор, который питал общую неприязнь к Свидетелям Иеговы, хотя, к его большому разочарованию, свидетелем был его собственный сын.How we lived our lives after the war was as varied as each man.
То, как мы прожили свою жизнь после войны, было таким же разнообразным, как и каждый мужчина.The slave’s owner was Philemon, also a Christian, who lived in Asia Minor.
Рабовладельцем был Филимон, тоже христианин, живший в Малой Азии.I like the first apartment where I lived on my own.
Мне нравится первая квартира, в которой я жил один.After that, Katrina lived in a house bought by the Rothsburgher corporation.
После этого Катрина жила в доме, купленном корпорацией Ротсбургеров.We lived in a prison with brass bands playing on the balconies.
Мы жили в тюрьме, на балконах играли духовые оркестры.Even as a little kid, I knew we lived in the shadow of financial and emotional collapse.
Даже будучи маленьким ребенком, я знал, что мы живем в тени финансового и эмоционального краха.King Solomon got absolutely nowhere with a young girl with whom he had fallen in love —and he was one of the wealthiest, wisest men who ever lived!
У царя Соломона ничего не вышло с молодой девушкой, в которую он влюбился, —а ведь он был одним из самых богатых и мудрых людей на свете!I lived in Kabylie, Algeria, for two years and I learned Berber there.
Я жил в Кабили, Алжир, два года и выучил там берберский.Well, until recently, Ellen lived with us.
Ну, до недавнего времени Эллен жила с нами.Bonica lived these parallel lives.
Боника прожила эти параллельные жизни.With a proper diet and exercise, he should have lived a long life.
При правильной диете и физических упражнениях он должен был прожить долгую жизнь.After his wife died, he lived for quite a few more years.
После смерти жены он прожил еще несколько лет.I lived with my father and mother and sister in a cottage not far from here.Until one day my father died.
Я жил с отцом, матерью и сестрой в коттедже недалеко отсюда. До одного дня мой отец умер.Everything that lives, lived, will live, carbon.
Все, что живет, живет, будет жить, углерод.Tom likes Boston better than any other place where he’s lived.
Том любит Бостон больше, чем любое другое место, где он жил.And what they found was that people who lived to 90 or 100 were more likely to have daf- 2 mutations — that is, changes in the gene that encodes the receptor for IGF- 1.
И они обнаружили, что люди, дожившие до 90 или 100 лет, чаще имели мутации daf — 2, то есть изменения в гене, кодирующем рецептор IGF — 1.The girl hid in the forest for some months… where she lived like an animal.
Девушка несколько месяцев пряталась в лесу … где жила как животное.Somebody who lived here had literary tastes after my own heart.
Кто-то из живущих здесь имел литературные пристрастия мне по душе.A woman who lived just a few blocks away had made the call from a cellular phone.
Женщина, которая жила всего в нескольких кварталах отсюда, позвонила по сотовому телефону.Sami and Layla haven’t lived together for six years.
Сами и Лейла не жили вместе шесть лет.The police found out where the criminal lived.
Полиция выяснила, где проживал преступник.He is the bravest soldier that ever lived.
Он самый храбрый солдат, который когда-либо жил.There on the top of a high mountain they built a small town of their own and lived in peace.
Там на вершине высокой горы они построили собственный городок и жили мирно.We will have lived here for ten years at the end of this month.
В конце этого месяца мы проживем здесь десять лет.We have lived in this town since 1960.
Мы жили в этом городе с 1960 года.We have lived in Tokyo for six years.
Мы живем в Токио шесть лет.We have lived in the U.S. for ten years now.
Мы живем в США уже десять лет.We have lived in this town for five years.
Мы живем в этом городе пять лет.My uncle has lived in Paris for ten years.
Мой дядя живет в Париже десять лет.My grandfather lived to be ninety.
Мой дедушка дожил до девяноста.My grandfather lived a long life.
Мой дед прожил долгую жизнь.I had lived in Osaka for eighteen years when I moved to Tokyo.
Я прожил в Осаке восемнадцать лет, когда переехал в Токио.I lived abroad for ten years.
Я жил за границей десять лет.I have lived here since 1990.
Я живу здесь с 1990 года.I have lived in Sasayama for three years.
Я живу в Сасаяме три года.I once lived in Rome.
Когда-то я жил в Риме.I have lived here a little over 60 years.
Я живу здесь чуть больше 60 лет.I have lived in this village for the past ten years.
Я живу в этой деревне последние десять лет.I’ve lived in Kobe before.
Я раньше жил в Кобе.I have not lived in Sanda since last year.
Я не живу в Санде с прошлого года.I have lived in Kobe since last year.
Я живу в Кобе с прошлого года.I lived in Sanda City last year.
В прошлом году я жил в городе Санда.I have lived in Kobe since yesterday.
Со вчерашнего дня живу в Кобе.I lived in Japan three years ago.
Я жил в Японии три года назад.I’ve lived here since I was a boy.
Я живу здесь с детства.I had lived in Sendai for ten years before I visited Matsushima.
Я жил в Сендае десять лет, прежде чем приехал в Мацусиму.I have lived here for a long time.
Я живу здесь очень давно.I asked where she lived.
Я спросил, где она живет.We have lived in Osaka six years.
Мы живем в Осаке шесть лет.Определение, фонетический (звуко-буквенный) разбор и разбор слова по составу
На данной странице представлено лексическое значение слова «жила», а также сделан звуко-буквенный разбор и разбор слова по составу с транскрипцией и ударениями.Оглавление:
- Значение слова
- Звуко-буквенный разбор
- Разбор по составу
Значение слова
1.
ЖИЛА1, ы, ж.
1. Обиходное название кровеносных сосудов, сухожилий. Жилы на лбу надулись. Рвать жилы (много и напряжённо работать; прост.).
2. Геологическое тело (в 4 знач.), образовавшееся в результате заполнения трещины горной породой, а также сама горная порода в такой трещине. Золотоносная ж. Напасть на золотую жилу (также перен.: о большой и неожиданной удаче).
• Тянуть жилы из кого (разг.) мучить, изводить кого-н. непосильными занятиями, чем-н. нудным, томительным.
| уменьш. жилка, и, ж.
| прил. жильный, ая, ое (спец.). Жильная струна (из жилы). Жильные горные породы. Жильное золото.
2.
ЖИЛА2, ы, м. и ж. (прост. презр.). Скупой, прижимистый человек, скряга.
Фонетический (звуко-буквенный) разбор
жила́
жила — слово из 2 слогов: жи-ла. Ударение падает на 2-й слог.
Транскрипция слова: [жыла]
ж — [ж] — согласный, звонкий парный, твёрдый (непарный, всегда произноится твёрдо), шипящий
и — [ы] — гласный, безударный
л — [л] — согласный, звонкий непарный, сонорный (всегда звонкий), твёрдый (парный)
а — [а] — гласный, ударный
В слове 4 буквы и 4 звука.
При разборе слова используются правила:
- Гласная и после букв ж, ш, ц означает звук [ы]
Цветовая схема: жила
Разбор слова «жила» по составу
жила
Части слова «жила»: жил/а
Состав слова:
жил — корень,
а — окончание,
жил — основа слова.
Транскрипция песни Yellow Submarine — The Beatles
Транскрипция
ин зэ тaун уэр ай уаз борн
лайвд э мэн ху сейлд ту си
энд хи тoулд ас ав хиз лайф
ин зэ лэнд ав ˈсабмэˌринз
сoу уи сейлд ап ту зэ сан
тил уи фaунд зэ си ав грин
энд уи лайвд биˈнис зэ уэйвз
ин ˈaуэр ˈйелoу ˈсабмэˌрин
уи ол лив ин э ˈйелoу ˈсабмэˌрин
ˈйелoу ˈсабмэˌрин, ˈйелoу ˈсабмэˌрин
уи ол лив ин э ˈйелoу ˈсабмэˌрин
ˈйелoу ˈсабмэˌрин, ˈйелoу ˈсабмэˌрин
энд ˈaуэр френдз ар ол эˈборд
ˈмени мор ав зем лив некст дор
энд зэ бэнд биˈгинз ту плей
уи ол лив ин э ˈйелoу ˈсабмэˌрин
ˈйелoу ˈсабмэˌрин, ˈйелoу ˈсабмэˌрин
уи ол лив ин э ˈйелoу ˈсабмэˌрин
ˈйелoу ˈсабмэˌрин, ˈйелoу ˈсабмэˌрин
фул спид эˈхед, ˈмистэр. ˈбoутсуэйн, фул спид эˈхед!
фул спид ит из, ˈсарджэнт!
кат зэ ˈкейбэл, драп зэ ˈкейбэл!
ай-ай, сёр, ай-ай!
ˈкэптэн, ˈкэптэн!
эз уи лив э лайф ав из (э лайф ав из)
ˈевриˌуан ав ас (ˈевриˌуан ав ас) хэз ол уи нид (хэз ол уи нид)
скай ав блу (скай ав блу) энд си ав грин (си ав грин)
ин ˈaуэр ˈйелoу (ин ˈaуэр ˈйелoу) ˈсабмэˌрин (ˈсабмэˌрин, а-ха)
уи ол лив ин э ˈйелoу ˈсабмэˌрин
ˈйелoу ˈсабмэˌрин, ˈйелoу ˈсабмэˌрин
уи ол лив ин э ˈйелoу ˈсабмэˌрин
ˈйелoу ˈсабмэˌрин, ˈйелoу ˈсабмэˌрин
уи ол лив ин э ˈйелoу ˈсабмэˌрин
ˈйелoу ˈсабмэˌрин, ˈйелoу ˈсабмэˌрин
уи ол лив ин э ˈйелoу ˈсабмэˌрин
ˈйелoу ˈсабмэˌрин, ˈйелoу ˈсабмэˌрин
Оригинал
In the town where I was bornLived a man who sailed to sea
And he told us of his life
In the land of submarines
So we sailed up to the sun
‘Til we found the sea of green
And we lived beneath the waves
In our yellow submarine
We all live in a yellow submarine
Yellow submarine, yellow submarine
We all live in a yellow submarine
Yellow submarine, yellow submarine
And our friends are all aboard
Many more of them live next door
And the band begins to play
We all live in a yellow submarine
Yellow submarine, yellow submarine
We all live in a yellow submarine
Yellow submarine, yellow submarine
Full speed ahead, Mr. Boatswain, full speed ahead!
Full speed it is, Sergeant!
Cut the cable, drop the cable!
Aye-aye, sir, aye-aye!
Captain, Captain!
As we live a life of ease (a life of ease)
Everyone of us (everyone of us) has all we need (has all we need)
Sky of blue (sky of blue) and sea of green (sea of green)
In our yellow (in our yellow) submarine (submarine, ah-ha)
We all live in a yellow submarine
Yellow submarine, yellow submarine
We all live in a yellow submarine
Yellow submarine, yellow submarine
We all live in a yellow submarine
Yellow submarine, yellow submarine
We all live in a yellow submarine
Yellow submarine, yellow submarine
MemorySecrets.ru
Тема: Тексты на английском языке
Язык: английский
Транскрипция: есть
Перевод: есть
Для прочтения этого текста на английском языке вам понадобится знание слов на тему «Животные. Animals». Для того чтобы слова было учить интересно и увлекательно воспользуйтесь нашими интерактивными карточками «Животные на английском в картинках».
Текст на английском языке с переводом «Маша и ее животные – Masha and her animals»
Once upon the time there lived Masha. She lived on the farm and had a lot of animals. Animals of different species could be found everywhere both in the house and outdoors. Domestic animals, namely cats and dogs were allowed to enter the house. Farm animals, such as cows, sheep and ducks were not permitted to come in.
Masha liked taking care of all animals that lived on the farm but primarily she adored her pets (domestic animals). She had two dogs, three cats, one parrot and some insects. Her dad and mom didn’t approve of her keeping insects as pets because they were afraid of her life. The spider was her favorite insect and she played with it most of her time. It’s well-known that spiders may bite, that’s why parents thought it to be able to injure their daughter. Nevertheless, a girl was very brave and was scared of nothing because she knew for sure that her spider wasn’t poisonous, it was domesticated.
Apart from domestic animals, farm animals and insects there were reptiles on her farm as well. It were chameleons, lizards and snakes. Masha also liked reptiles but was cautious about them because she was extremely smart and was aware of the grave danger which snakes represented.
Маша и ее животные
Жила-была Маша. Жила она на ферме, и было у нее много животных. Животных разных видов можно было встретить повсюду — как внутри дома, так и за его пределами. Домашним животным, а именно — кошкам и собакам, разрешали входить в дом. Сельскохозяйственным животным же, таким как петухам, коровам, овечкам и уткам, было запрещено входить внутрь дома.
Маше нравилось ухаживать за всеми животными, которые жили на ферме, но в первую очередь она обожала своих домашних питомцев. У нее было две собаки, три кошки, один попугай и несколько насекомых. Ее папа и мама не одобряли то, что она держала насекомых в качестве питомцев, потому что они боялись за ее жизнь. Паук был ее любимым насекомым, и она играла с ним большую часть своего времени. Хорошо известно то, что пауки кусаются, поэтому родители думали, что паук может навредить их дочери. Тем не менее, девочка была очень храброй и не боялась ничего, потому что она знала наверняка, что ее паук не ядовитый, а ручной.
Помимо домашних животных, сельскохозяйственных и насекомых, на ферме жили и пресмыкающиеся (рептилии). Это были хамелеоны, ящерицы и змеи. Маша тоже любила рептилий, однако была очень осторожна с ними, потому что была чрезвычайно умной девочкой и хорошо знала о серьезной опасности, которую несли змеи.
Срочно!!!Транскрипция предложения »Жили-были старик со старухой»
Вольфганг амадей моцарт. джузеппе верди. иоганн штраус. иоганн себастьян бах людвиг ван бетховен. петр ильич чайковский. сергей васильевич рахманинов. ранц петер шуберт
Русский язык постоянно пополняется новыми словами. Основной путь
обогащения словарного состава — образование слов различными способами от иных слов или основ.Слово, от которого образованно другое слово, называется
производящим, а образованное от него слово производным.При последовательном образовании слов друг от друга получается
словообразовательная цепочка однокоренных слов.
Слова могут образовываться с помощью приставок, суффиксов,
а также сложением нескольких основ или слов.
Также есть морфемные способы словообразования .
Суффиксальный — суффикс присоединяется к основе производящего слова.
Приставочный — приставка присоединяется к целому производящему слову.
Приставочно-суффиксальный — приставка и суффикс одновременно присоединяются к основе производящего слова.
Бессуффиксный — этим способом образуются имена существительные:
1) от прилагательных, в которых отбрасывается окончание,либо окончание и суффикс
2)от глаголов, отсечением от начальной формы суффикса и окончания
В процессе образования слов может происходить:
1)чередование звуков;
2)усечение конечных суффиксов или их частей;
3)одновременно чередование и усечение:
Слова могут образовываться сложением основ, с помощью соединительных гласных или без них
зуб корень, ы- окончание, зубаз- зуб — корень, ах-0 окончание (это формы одного слова) зубной- зуб-корень, н-суффикс, ой- окончание. Это новое слово по лотношению к предыдущим. огонь — огненный огонь — оскова равная корню. огненный огн — корень, енн-суффикс, ый окончание. это разные слова.
А где текст?) Блин, правда! Где текст?
Фактор транскрипции ответа митохондриального развернутого белка ATFS-1 способствует долголетию долгоживущего митохондриального мутанта за счет активации путей ответа на стресс | BMC Biology
Митохондриальный ответ развернутого белка активируется у долгоживущих митохондриальных мутантов
Чтобы изучить роль mitoUPR в продолжительности жизни митохондриальных мутантов clk-1 , isp-1 и nuo -6 , мы сначала попытались подтвердить, что mitoUPR активируется у этих мутантов [6, 12, 23].Для этого мы скрестили митохондриальные мутантные штаммы с репортерным штаммом для целевого гена mitoUPR hsp-6 ( Phsp-6 :: GFP ) [2] . Мы обнаружили, что все черви clk-1 , isp-1 и nuo-6 демонстрируют повышенную флуоресценцию по сравнению с червями дикого типа (рис. 1а), что указывает на активацию mitoUPR. Мы обнаружили, что флуоресценция репортера mitoUPR оставалась значительно увеличенной до 5 дня взрослой жизни (рис. 1b). Поскольку GFP-меченные белки могут иметь период полужизни в несколько дней, возможно, что флуоресценция репортерного штамма mitoUPR во взрослом возрасте является результатом активности mitoUPR во время развития.Чтобы подтвердить, что mitoUPR остается активированным во взрослом возрасте, мы изучили данные RNAseq для мутантов clk-1 , isp-1 и nuo-6 на стадии молодого взрослого человека [24]. Мы обнаружили, что мРНК hsp-6 значительно увеличена во всех трех штаммах и коррелирует с уровнями мРНК atfs-1 (дополнительный файл 1: рисунок S1).
Рис. 1ATFS-1-зависимая активация ответа митохондриального развернутого белка (mitoUPR) у долгоживущих митохондриальных мутантов. a Для мониторинга активации mitoUPR митохондриальные мутанты clk-1 , isp-1 и nuo-6 были скрещены с штаммом транскрипционного репортера Phsp-6 :: GFP . Все три штамма показали повышенную экспрессию GFP, что указывает на активацию mitoUPR. b Динамика во взрослом возрасте показывает, что активация mitoUPR сохраняется в раннем взрослом возрасте. c , d Сходным образом все три митохондриальных мутанта демонстрируют активацию mitoUPR уже на 1-й и 2-й день после вылупления. e Активация репортера mitoUPR зависит от фактора транскрипции ATFS-1, так как atfs-1 РНКи предотвращает экспрессию GFP у червей nuo-6; Phsp-6 :: GFP червей . Нокдаун atfs-1 был инициирован на родительской стадии L4, и флуоресценция была исследована у молодых взрослых потомков. Планки погрешностей указывают на SEM. * p <0,05, ** p <0,01, *** p <0,001. См. Дополнительный файл 5: Таблица S5 для получения подробной информации о N и репликах
Затем мы попытались определить, активируется ли mitoUPR также во время личиночного развития.Мы исследовали червей через 1 и 2 дня после вылупления и в обоих случаях обнаружили, что mitoUPR активируется у червей clk-1 , isp-1 и nuo-6 (рис. 1c, d). Наконец, мы хотели определить, зависит ли активация mitoUPR от ATFS-1. Соответственно, мы обработали clk-1; Phsp-6 :: GFP , isp-1; Phsp-6 :: GFP и nuo-6; Phsp-6 :: GFP червей с помощью РНКи, нацеленной на atfs- 1 и обнаружил, что сбивание atfs-1 предотвращает индукцию mitoUPR (рис.1e, дополнительный файл 1: рисунок S2). В совокупности эти результаты демонстрируют, что долгоживущие митохондриальные мутантные штаммы обнаруживают ATFS-1-зависимую активацию mitoUPR в процессе развития и в раннем взрослом возрасте.
Поскольку повышенных уровней АФК достаточно для активации mitoUPR [5, 25] и было показано, что уровни АФК повышены у долгоживущих митохондриальных мутантов [26, 27], мы затем попытались определить степень, в которой активация mitoUPR зависит от повышенных АФК.Для этого мы обработали червей nuo-6 двумя разными антиоксидантами, 10 мМ витамина С и 10 мМ N-ацетилцистеина, которые, как ранее было показано, сокращают продолжительность жизни долгоживущих митохондриальных мутантов [26, 28] . Используя флуоресценцию репортера Phsp-6 :: GFP в качестве измерения активности mitoUPR, мы обнаружили, что лечение антиоксидантами не влияло на активацию mitoUPR (дополнительный файл 1: рисунок S3). Это говорит о том, что повышение ROS не требуется для мутации nuo-6 для активации mitoUPR.Однако также возможно, что лечение антиоксидантами не привело к достаточному снижению уровней ROS или не уменьшило нужную популяцию ROS, чтобы наблюдать эффект на активацию mitoUPR.
Ответ митохондриального развернутого белка необходим для выживания и продолжительности жизни долгоживущих митохондриальных мутантов
Для выяснения роли mitoUPR в продолжительности жизни clk-1 , isp-1 и nuo-6 , мы исследовали, как снижение экспрессии atfs-1 влияет на их продолжительность жизни.Мы использовали три разные парадигмы возрастающей серьезности: atfs-1 РНКи, начинающаяся в экспериментальном поколении L4, atfs-1 РНКи, начинающаяся в родительском поколении L4, и делеционная мутация atfs-1 (gk3094) .
В экспериментальной парадигме L4 RNAi червям позволяют развиваться на пластинах NGM до стадии L4 развития, когда они переносятся на atfs-1 RNAi пластин. Используя эту парадигму, мы обнаружили, что atfs-1 РНКи вызывает небольшое уменьшение продолжительности жизни clk-1 (рис.2а), но не повлияли на продолжительность жизни червей isp-1 (рис. 2b) или nuo-6 (рис. 2c). На 2 день взросления эта парадигма привела к значительному снижению уровней atfs-1 , а также целевого гена hsp-6 atfs-1 (рис. 2d), но не повлияла на экспрессию цитозольного Ответ развернутого белка нацелен на hsp-16.11 или hsp-16.2 (которые, как мы обнаружили, значительно увеличиваются при мутации atfs-1 — см. рис.6).
Рис. 2ATFS-1 необходим для выживания и долгожительства долгоживущих митохондриальных мутантов. Уровни atfs-1 были снижены с использованием трех парадигм возрастающей серьезности: РНКи, начиная с экспериментального поколения L4 (левый столбец), РНКи, начиная с родительского поколения L4 (средний столбец) и делеционной мутации (правый столбец). Уменьшение экспрессии atfs-1 , начиная с стадии разработки L4, практически не повлияло на продолжительность жизни clk-1 ( a ), isp-1 ( b ) или nuo. -6 ( c ) червей, несмотря на снижение экспрессии atfs-1 и atfs-1 целевого гена hsp-6 ( d ).Снижение экспрессии atfs-1 , начиная с родительского поколения, приводило к остановке развития у червей clk-1 ( e ) и isp-1 ( f ), а у червей nuo-6 выросли до зрелого возраста и показали уменьшенную продолжительность жизни ( г, ). Это лечение снизило уровни atfs-1 и hsp-6 и привело к усилению регуляции hsp-16.11 и hsp-16.2 ( h ). Удаление atfs-1 привело к остановке развития у червей clk-1 ( i ) и isp-1 ( j ).Напротив, nuo-6; atfs-1 червей жизнеспособны, а делеция atfs-1 полностью вернула продолжительность жизни nuo-6 к дикому типу ( k ). Делеция atfs-1 снизила экспрессию гена atfs-1 и hsp-6 и повысила уровни hsp-16.11 и hsp-16.2 мРНК ( l ). Указывается значение между красной и фиолетовой линиями. Планки погрешностей указывают на SEM. * p <0,05, ** p <0.01, *** р <0,001. Необработанные данные о сроке службы см. В дополнительном файле 4: Таблица S4. См. Дополнительный файл 5: Таблица S5 для получения подробной информации о N и репликах
В парадигме родительского L4 черви переносятся на планшеты atfs-1 RNAi на стадии L4. Когда эти черви становятся беременными имаго, их переносят на вторую пластину atfs-1 РНКи и дают возможность откладывать яйца в течение 24 часов. Затем измеряется продолжительность жизни потомства F1, когда черви достигают зрелого возраста. В рамках этой парадигмы мы обнаружили, что только черви дикого типа и nuo-6 способны развиваться до взрослого состояния, в то время как черви clk-1 и isp-1 задерживаются во время личиночного развития (рис.2д, е; Дополнительный файл 1: Рисунки S4, S5), как было замечено ранее [6]. Мы обнаружили, что сбивание atfs-1 , начиная со стадии L4 в родительском поколении, значительно снижает продолжительность жизни nuo-6 , но не влияет на продолжительность жизни дикого типа (рис. 2g). Эта парадигма значительно снизила уровни экспрессии atfs-1 и hsp-6 до такой же степени, как мы наблюдали в экспериментальной парадигме L4. В отличие от экспериментальной парадигмы L4, сбивание atfs-1 , начиная с родительского поколения, приводило к значительному увеличению экспрессии hsp-16.11 и hsp-16.2 , которые участвуют в реакции цитозольного развернутого белка (также известной как реакция теплового шока) (рис. 2h). Это говорит о том, что степень протеотоксического стресса может быть выше, когда atfs-1 сбивается в течение всей жизни.
Наконец, мы исследовали влияние делеционной мутации atfs-1 на продолжительность жизни митохондриальных мутантов. Мы обнаружили, что двойные мутанты clk-1; atfs-1 дают мало потомства и что потомство останавливается на стадии развития L1 или L2 (рис.2i). Сходным образом мы обнаружили, что isp-1; atfs-1 двойных мутантов арестовываются на стадии L2 или L3 (рис. 2j). Напротив, двойные мутанты nuo-6; atfs-1 оказались жизнеспособными и фертильными. Изучение продолжительности жизни этих двойных мутантов показало, что делеция atfs-1 полностью вернула продолжительность жизни nuo-6 к дикому типу, но не повлияла на продолжительность жизни дикого типа (рис. 2k). Делеция atfs-1 снижала экспрессию atfs-1 и hsp-6 и увеличивала экспрессию hsp-16.11 и hsp-16.2 (рис. 2л). В совокупности эти результаты показывают, что atfs-1 требуется для развития червей clk-1 и isp-1 до взрослого состояния и для продолжительности жизни червей nuo-6 . Потеря atfs-1 не снижает продолжительность жизни дикого типа, а у митохондриальных мутантов atfs-1 не требуется во взрослом возрасте для нормальной продолжительности жизни.
Поскольку мы наблюдали эффект atfs-1 РНКи на продолжительность жизни мутантов nuo-6 , мы решили исследовать другие компоненты mitoUPR, включая UBL-5, DVE-1 и HAF-1.Мы использовали родительскую парадигму L4, в которой мы наблюдали эффект atfs-1 РНКи у червей nuo-6 . В этих условиях мы обнаружили, что уничтожение dve-1 или ubl-5 предотвратило развитие червей nuo-6 до взрослого состояния (дополнительный файл 1: Рисунок S6).
Поскольку nuo-6 черви могли развиться до стадии L4 на ubl-5 РНКи, мы сравнили выживаемость этих червей с червями, выращенными на пустых векторных РНКи. В соответствии с нашими наблюдениями с atfs-1 RNAi, мы обнаружили, что выживаемость L4 nuo-6 червей на ubl-5 RNAi была заметно снижена по сравнению с EV RNAi (дополнительный файл 1: рисунок S7A).На haf-1 RNAi, nuo-6 черви развились до зрелого возраста и показали аналогичную продолжительность жизни червям, выращенным на EV RNAi (дополнительный файл 1: рисунок S7B). Это предполагает, что механизм, с помощью которого нарушение nuo-6 активирует mitoUPR, не требует HAF-1. Как и в случае с результатами наших исследований продолжительности жизни, мы обнаружили, что haf-1 РНКи имели другой эффект на активацию ответа цитозольного развернутого белка по сравнению с другими компонентами mitoUPR. В то время как нокдаун atfs-1 , ubl-5 или dve-1 приводил к повышенной экспрессии генов-мишеней ответа цитозольного развернутого белка, hsp-16.11 и hsp-16.12 , haf-1 РНКи не оказали значительного влияния на экспрессию любого гена (дополнительный файл 1: Рисунок S8). Опять же, это предполагает, что нарушение haf-1 в митохондриях влияет на mitoUPR иначе, чем нарушение ядерных факторов atfs-1 , dve-1 и ubl-5.
Поскольку удаление гена митохондриальной супероксиддисмутазы sod-2 [29] и умеренное ингибирование митохондриальной функции были предложены для увеличения продолжительности жизни за счет повышения уровней митохондриального супероксида [26], мы также исследовали роль митохондриального UPR. в долговечности дерн-2 червей.Как и в случае долгоживущих митохондриальных мутантов, мы обнаружили, что mitoUPR активируется у мутантов sod-2 , как было измерено с использованием репортерных штаммов Phsp-6 :: GFP (дополнительный файл 1: рисунок S9A). Однако мы обнаружили, что подавление экспрессии atfs-1 не препятствовало развитию червей sod-2 и не влияло на их продолжительность жизни (дополнительный файл 1: Рисунок S9B, C). Это указывает на то, что механизмы, лежащие в основе долгожительства у мутантов sod-2 , по крайней мере частично, отличаются от долгоживущих митохондриальных мутантов.
Потеря экспрессии ATFS-1 во время развития или во взрослом возрасте сама по себе не сокращает продолжительность жизни
nuo-6Тот факт, что сбивание atfs-1 во взрослом возрасте практически не повлияло на продолжительность жизни, предполагает возможность того, что ATFS-1 влияет на продолжительность жизни во время развития. Соответственно, мы стремились определить, будет ли сбивание уровней atfs-1 только во время разработки достаточно для уменьшения продолжительности жизни. Для этого мы использовали подход, при котором червей выращивали на atfs-1 РНКи во время развития и затем переносили на РНКи против дайсера ( dcr-1 ) во взрослом возрасте [11, 12, 30].Путем нокдауна дайсера, который является важным компонентом аппарата РНКи, следует предотвратить ингибирование экспрессии atfs-1 , позволяя восстановить уровней atfs-1 . Черви выращивали на пустом векторе или atfs-1 РНКи, начиная с стадии развития L4 родительского поколения, а затем переносили либо на планшеты dcr-1 RNAi (для восстановления экспрессии atfs-1, ) или atfs-1. Планшеты РНКи (для продолжения нокдауна atfs-1 ) во взрослом возрасте (рис.3а).
Рис. 3Потеря ATFS-1 во время развития или во взрослом возрасте недостаточна для уменьшения продолжительности жизни nuo-6 . a Чтобы изучить временные требования для роли ATFS-1 в продолжительности жизни nuo-6 и активации mitoUPR, nuo-6 червей обрабатывали RNAi против atfs-1 только для разработки ( atfs-1 ➔ dcr-1 ), только для взрослых (EV ➔ atfs-1 ) или как для развития, так и для взросления ( atfs-1 ➔ atfs-1 ).Количественная ОТ-ПЦР в реальном времени была использована для подтверждения нокдауна atfs-1 , dcr-1 и целевого гена ATFS-1 hsp-6. Обработка dcr-1 РНКи в зрелом возрасте эффективно снижала экспрессию dcr-1 ( b ) . Уровни мРНК atfs-1 были снижены на atfs-1 РНКи только во взрослом возрасте или в период развития и во взрослом возрасте ( c ). Перенос РНКи atfs-1 в РНКи dcr-1 приводил к восстановлению уровней мРНК atfs-1 по направлению к контролю EV.Точно так же уровни целевого гена atfs-1 hsp-6 были снижены на atfs-1 РНКи только во взрослом возрасте или в период развития и во взрослом возрасте ( d ). Перенос РНКи atfs-1 в РНКи dcr-1 приводил к полному восстановлению экспрессии hsp-6 . e Сбивание уровней atfs-1 только во взрослом возрасте не влияет на продолжительность жизни nuo-6 . f Сбивание atfs-1 только во время разработки не приводит к значительному сокращению срока службы nuo-6 .Напротив, atfs-1 РНКи, примененные как во время развития, так и во взрослом возрасте, снижали продолжительность жизни nuo-6 . Планки погрешностей указывают на SEM. ** p <0,01, *** p <0,001. Необработанные данные о сроке службы см. В дополнительном файле 4: Таблица S4. См. Дополнительный файл 5: Таблица S5 для получения подробной информации о N и репликах
Поскольку atfs-1 абсолютно необходим в червях clk-1 и isp-1 для развития до взрослого возраста, мы завершили эти эксперименты, и все остальные эксперименты на червях nuo-6 .Мы использовали количественную ОТ-ПЦР в реальном времени на 2-й день взросления, чтобы исследовать уровни dcr-1 , atfs-1 и hsp-6. Мы обнаружили, что перенос на планшеты dcr-1 РНКи во взрослом возрасте эффективно снижает экспрессию dcr-1 (рис. 3b). Уровни atfs-1 эффективно подавлялись atfs-1 РНКи и значительно повышались при переносе на dcr-1 РНКи (рис. 3c). Важно отметить, что обработка atfs-1 РНКи во взрослом возрасте значительно снижала экспрессию hsp-6 , тогда как hsp-6 экспрессия у червей, выращенных на atfs-1 РНКи во время развития и перенесенных на dcr-1 РНКи взросление (парадигма atfs-1 ➔ dcr-1 ) было эквивалентно червям, которые не лечились atfs-1 РНКи (парадигма EV ➔ dcr-1 ) (рис.3d).
Показав, что уровней atfs-1 были сбиты именно во время разработки, мы затем исследовали результирующее влияние на продолжительность жизни. Мы обнаружили, что подавление экспрессии atfs-1 и во взрослом возрасте (парадигма EV ➔ atfs-1 ) не снижает продолжительность жизни червей nuo-6 и (рис. 3e). Точно так же только уменьшение экспрессии atfs-1 во время развития (парадигма atfs-1 ➔ dcr-1 ) было недостаточным для уменьшения продолжительности жизни nuo-6 (рис.3е). Обратите внимание, что продолжительность жизни червей nuo-6 , обработанных парадигмой atfs-1 ➔ dcr-1 , значительно увеличилась по сравнению с червями, обработанными парадигмой atfs-1 ➔ atfs-1 или atfs-1 ➔ парадигма EV (дополнительный файл 1: рисунок S10), предоставляя дополнительные доказательства того, что перенос dcr-1 РНКи был эффективен при восстановлении экспрессии atfs-1 (продолжение нокдауна atfs-1 в atfs-1 ➔ Парадигма электромобилей приводит к сокращению срока службы).Таким образом, для того, чтобы нокдаун atfs-1 уменьшил продолжительность жизни nuo-6 , уровни atfs-1 должны были быть уменьшены как во время развития, так и во взрослом возрасте.
Потеря ATFS-1 влияет на физиологические показатели и митохондриальную функцию червей
nuo-6Чтобы начать исследовать механизм, с помощью которого потеря atfs-1 сокращает продолжительность жизни червей nuo-6 , мы сравнили физиологические показатели и функция митохондрий у червей nuo-6, и nuo-6; atfs-1 .Мы обнаружили, что мутация atfs-1 увеличивала как эмбриональную летальность, так и задержку развития, особенно у червей nuo-6 , но не у червей дикого типа (рис. 4a, b). Из тех червей, которые действительно развиваются до взрослого состояния, мутантов atfs-1 развиваются медленнее, чем черви дикого типа, а черви nuo-6; atfs-1 достигают зрелости быстрее, чем черви nuo-6 (рис. 4в). Потеря atfs-1 приводит к уменьшению размера расплода, но не приводит к дальнейшему уменьшению и без того низкого размера выводка червей nuo-6 (рис.4г). Точно так же мутанты atfs-1 имеют более низкую скорость обмолота, чем черви дикого типа, но мутация не усугубляет дефицит обмолота, присутствующий у мутантов nuo-6 (рис. 4e). Потеря atfs-1 приводит к более медленной скорости дефекации и уменьшению длины тела как у червей дикого типа, так и у червей nuo-6 (рис. 4f, g).
Рис. 4Потеря atfs-1 изменяет физиологические показатели митохондриального мутанта nuo-6 . a nuo-6 черви имеют повышенную эмбриональную летальность по сравнению с червями дикого типа, а летальность еще больше увеличивается за счет делеции atfs-1 . b В то время как все черви nuo-6 и развиваются до взрослого состояния после вылупления, часть червей nuo-6; atfs-1 задерживается во время личиночного развития. c Удаление atfs-1 замедляет развитие у червей дикого типа, но ускоряет время развития у червей nuo-6 . d Потеря atfs-1 уменьшает размер расплода у червей дикого типа, но не приводит к дальнейшему уменьшению небольшого размера расплода у червей nuo-6 . e Делеция atfs-1 снижает скорость передвижения у червей дикого типа, но не усугубляет медленное движение червей nuo-6 . Потеря atfs-1 вызывает медленную скорость дефекации ( f ), уменьшение длины тела ( г ) и снижение потребления кислорода ( h ) как у червей дикого типа, так и у червей nuo-6 , но существенно не влияет на уровень АТФ ( i ). Планки погрешностей указывают на SEM. * p <0.05, ** p <0,01, *** p <0,001. См. Дополнительный файл 5: Таблица S5 для получения подробной информации о N и репликах
При изучении функции митохондрий мы определили скорость окислительного фосфорилирования путем измерения потребления кислорода на одного червя с помощью анализатора внеклеточного потока. Мы обнаружили, что потребление кислорода на одного червя снижается за счет потери atfs-1 как у червей дикого типа, так и у мутантов nuo-6 (рис. 4h). Чтобы определить, приводит ли скорость потребления кислорода к изменению уровней АТФ, мы использовали набор на основе люцигенина для измерения уровней АТФ.Мы обнаружили, что мутация atfs-1 не снижает уровни АТФ у червей дикого типа или nuo-6 (рис. 4i). Чтобы понять взаимосвязь между потреблением кислорода и уровнями АТФ, мы исследовали влияние мутации atfs-1 на уровни АФК у червей дикого типа и nuo-6 . В соответствии со сниженными уровнями потребления кислорода уровни ROS, обнаруженные с помощью флуоресценции дихлорфлуоресцеина (DCF), были значительно снижены при делеции atfs-1 (дополнительный файл 1: Рисунок S11).В совокупности эти результаты показывают, что нарушение atfs-1 отрицательно влияет на физиологические показатели червей и функцию митохондрий, и это может, по крайней мере, частично объяснить пагубное влияние мутации atfs-1 на продолжительность жизни nuo-6 и способность clk-1 и isp-1 червей, чтобы развиться до взрослого состояния.
Потеря ATFS-1 снижает устойчивость к множественным стрессам
Поскольку способность противостоять множественным стрессам связана с долгим сроком службы, мы затем исследовали влияние мутации atfs-1 на стрессоустойчивость в nuo-6 черви.Чувствительность к окислительному стрессу оценивалась путем воздействия на червей 2 мМ параквата, соединения, которое увеличивает уровень супероксида, прежде всего, в митохондриях. Мы обнаружили, что черви nuo-6 обладают повышенной выживаемостью по сравнению с червями дикого типа и что мутация atfs-1 полностью возвращает их устойчивость к стрессу к дикому типу (рис. 5a). atfs-1 мутанты также показали небольшое снижение устойчивости к окислительному стрессу по сравнению с червями дикого типа.
Фиг.5Потеря atfs-1 снижает повышенную устойчивость к множественным стрессам у долгоживущих червей nuo-6 и снижает стрессоустойчивость у червей дикого типа. a Чувствительность к окислительному стрессу оценивалась путем воздействия на червей планшетов, содержащих 2 мМ параквата (PQ). nuo-6 Черви показали повышенную устойчивость к окислительному стрессу, который полностью снимался удалением atfs-1. b Чувствительность к осмотическому стрессу оценивали путем воздействия на червей 550 мМ NaCl и количественной оценки выживаемости через 48 часов. nuo-6 Черви демонстрируют повышенную выживаемость по сравнению с червями дикого типа. Потеря atfs-1 привела к снижению выживаемости как у червей дикого типа, так и у червей nuo-6 . c Для измерения чувствительности к тепловому стрессу червей инкубировали при 37 ° C, а выживание отслеживали ежечасно. nuo-6 Черви демонстрируют повышенную выживаемость по сравнению с червями дикого типа. В обоих случаях выживаемость при тепловом стрессе снизилась на атс-1. d Удаление atfs-1 вызывает повышенную чувствительность к аноксии как у червей дикого типа, так и у червей nuo-6 .Планки погрешностей указывают на SEM. На панелях A и C значение p указывает на значимость между nuo-6 и nuo-6; atfs-1 червей. ** p <0,01, *** p <0,001. См. Дополнительный файл 5: Таблица S5 для получения подробной информации о N и репликах
. Чувствительность к осмотическому стрессу количественно определяли путем переноса червей на чашки, содержащие 550 мМ NaCl (в 11 раз больше их нормальной концентрации соли), и изучения выживаемости через 2 дня. И снова мы обнаружили, что черви nuo-6 обладают повышенной устойчивостью к осмотическому стрессу, которая снижается в присутствии мутации atfs-1 , а мутанты atfs-1 имеют меньшую выживаемость по сравнению с червями дикого типа ( Инжир.5б).
В тесте теплового стресса черви подвергаются тепловому стрессу 37 ° C, и их выживаемость отслеживается ежечасно. nuo-6 Черви показали умеренно повышенную устойчивость к тепловому стрессу (рис. 5c). Потеря atfs-1 снизила выживаемость как nuo-6, , так и червей дикого типа во время теплового стресса (рис. 5c).
Наконец, мы исследовали выживаемость после 2 дней аноксии с однодневным восстановлением. Мы обнаружили, что nuo-6 червей имели эквивалентную выживаемость дикому типу, и что в обоих случаях выживаемость была снижена мутацией atfs-1 (рис.5г). В совокупности эти результаты показывают, что ATFS-1 важен для выживания при множественных формах стресса. Снижение стрессоустойчивости у двойного мутанта nuo-6; atfs-1 может, по крайней мере, частично объяснить снижение продолжительности жизни по сравнению с мутантами nuo-6 .
nuo-6 черви демонстрируют зависимые от ATFS-1 изменения в экспрессии генов, участвующих в стрессовой реакции и метаболизмеДля дальнейшего изучения механизма, с помощью которого потеря atfs-1 сокращает продолжительность жизни nuo-6 червей мы сравнили экспрессию генов в мутантах nuo-6 и nuo-6; atfs-1 .Мы предположили, что опосредованные ATFS-1 изменения в экспрессии генов способствуют долгой жизни червей nuo-6 . Соответственно, мы исследовали экспрессию генов, участвующих в стрессовой реакции или метаболизме, чтобы идентифицировать гены, которые модулируются у червей nuo-6 и восстанавливаются до дикого типа за счет потери atfs-1. В отличие от ATFS-1-зависимой активации mitoUPR ( hsp-6 ), ни ответ белка эндоплазматического ретикулума ( hsp-4 ), ни ответ цитозольного белка ( hsp-16.2 ) активируется у червей nuo-6 (рис. 6a – c).
Рис. 6ATFS-1 опосредует специфические транскрипционные изменения у червей nuo-6 . Количественная ОТ-ПЦР в реальном времени использовалась для сравнения экспрессии выбранных генов стрессовой реакции и метаболизма у червей nuo-6 и nuo-6; atfs-1 . a nuo-6 черви показали пониженную экспрессию hsp-4 , который является маркером ответа на развернутый белок эндоплазматического ретикулума. b Экспрессия hsp-6 была увеличена у червей nuo-6 , и это увеличение было полностью предотвращено потерей atfs-1. c Удаление atfs-1 привело к повышенной экспрессии hsp-16.2 , который является маркером ответа цитозольного развернутого белка. d nuo-6 черви демонстрировали повышенную экспрессию целевого гена SKN-1 / NRF2 gst-4 , что предотвращалось делецией atfs-1 . e Экспрессия гена-мишени HIF-1 nhr-57 повышается у червей nuo-6 и восстанавливается до дикого типа с помощью делеции atfs-1 . Гены-мишени DAF-16 / FOXO mtl-1 ( f ), sodh-1 ( g ) и sod-3 ( h ) активируются в nuo-6 . worms и для этого требуется atfs-1 . Точно так же fmo-2 ( i ) и cdr-2 ( j ), которые участвуют в детоксикации ксенобиотиков, демонстрируют ATFS-1-зависимое увеличение экспрессии в мутантах nuo-6 . k — n Выбранные гены, участвующие в метаболизме ( ldh-1 , aldo-1 , gpd-2 , acs-2 ), демонстрируют ATFS-1-зависимое увеличение экспрессии в nuo. -6 червей. Планки погрешностей указывают на SEM. * p <0,05, ** p <0,01, *** p <0,001. См. Дополнительный файл 5: Таблица S5 для получения подробной информации о N и репликах
Интересно, что делеция atfs-1 привела к заметной активации цитозольного UPR, что измерялось увеличением hsp-16.2 выражение. Предварительное изучение этого результата показало, что активация hsp-16.2 (и hsp-16.11 ) у мутантов atfs-1 зависит от фактора теплового шока HSF-1, но не зависит от фактора транскрипции FOXO DAF-16 ( Дополнительный файл 1: Рисунок S12) [31]. Для дальнейшего изучения этой взаимосвязи мы создали двойных мутантов atfs-1; hsf-1 . Мы обнаружили, что двойные мутанты atfs-1; hsf-1 вредны для здоровья, на что указывает высокая стерильность и низкий размер расплода.Это предполагает, что активация цитозольного UPR у мутантов atfs-1 может быть компенсаторным ответом на нарушение mitoUPR.
Изучение других путей стрессовой реакции показало, что стрессовый ответ, опосредованный SKN-1 ( gst-4 ), опосредованный HIF-1 ответ гипоксии ( nhr-57 ) и опосредованный DAF-16 стресс response ( mtl-1 , sodh-1 , sod-3 ) все активированы в червях nuo-6 (рис.6д – ч; Дополнительный файл 1: Рисунок S13), который может способствовать повышенной устойчивости к стрессу, которую мы наблюдали у мутантов nuo-6 . В каждом случае потеря atfs-1 уменьшала степень активации каждого стрессового ответа, предполагая, что активация mitoUPR может привести к активации других путей стрессовой реакции. Обратите внимание, что мы и другие также наблюдали повышенную регуляцию HIF-1 target nhr-57 в clk-1 и isp-1 мутантов [27, 32, 33].Поскольку уровни АФК повышены у мутантов nuo-6 [26], а повышенных АФК достаточно для активации пути HIF-1 [25], вероятно, что АФК ответственны за активацию HIF-1 в nuo-6 . черви.
Мы также исследовали уровни экспрессии fmo-2 , флавинсодержащей моноксигеназы, участвующей в детоксикации ксенобиотиков, которая, как недавно было показано, модулирует продолжительность жизни [34], и cdr-2 , глутатион-S-трансферазы, также участвующей в детоксикация ксенобиотиков и участие в продолжительности жизни мутантов isp-1 [35].Как и в случае с другими генами стрессовой реакции, мы наблюдали повышенную экспрессию у мутантов nuo-6 , которая была значительно снижена при потере atfs-1 (рис. 6i, j). Обратите внимание, что другие антиоксидантные ферменты, включая супероксиддисмутазы ( sod-1 , sod-2 , sod-4 , sod-5 ), каталазы ( ctl-1 , ctl-2 , ctl-3 ), и пероксиредоксины ( prdx-2 , prdx-6 ), за исключением prdx-3 , не пострадали от потери atfs-1 в диком типе или nuo -6 червей (дополнительный файл 1: Рисунок S14).
Поскольку было показано, что мутанты nuo-6 изменяют экспрессию метаболических генов [36], мы исследовали роль ATFS-1 в экспрессии избранных метаболических генов. Мы выбрали гены, которые, как мы ранее обнаружили, были увеличены у червей nuo-6 с использованием RNAseq, включая лактатдегидрогеназу ( ldh-1 ), фруктозобифосфатальдолазу ( альдо-1 ), глицеральдегид-3-фосфат-gpd ( gpd). -2 ) и жирной ацил-КоА синтетазы ( acs-2 ) [37].Мы обнаружили, что все эти гены показали повышенную экспрессию у червей nuo-6 и , которая зависит от atfs-1 (рис. 6k – n).
Специфические гены стрессового ответа и метаболизма, регулируемые ATFS-1, необходимы для продолжительности жизни
nuo-6Чтобы изучить роль специфических изменений экспрессии генов, индуцированных mitoUPR, в продолжительной продолжительности жизни мутантов nuo-6 мы снизили экспрессию генов, которые, как мы обнаружили, имеют повышенную регуляцию у мутантов nuo-6 и , зависимым от ATFS-1 образом, и измерили продолжительность жизни.Мы уменьшили экспрессию gst-4 , mtl-1 , sodh-1 , fmo-2 , cdr-2 , ldh-1 , aldo-1 и acs. -2 с использованием РНКи, начиная со стадии L4 родительского поколения. Для hsp-6 мы начали обработку РНКи на стадии L4 экспериментального поколения, поскольку hsp-6 РНКи могут вызывать эмбриональную летальность [38]. Для sod-3 мы использовали генетическую мутацию, а не RNAi, чтобы избежать возможности сбить его паралог sod-2 , что может усложнить интерпретацию результата, поскольку потеря sod-2 увеличивает продолжительность жизни [29 ].Мы также использовали генетическую мутацию для hif-1 , чтобы мы могли сравнить результаты с предыдущими результатами для мутантов clk-1 и isp-1 [33]. Обратите внимание, что наблюдались противоречивые результаты в отношении продолжительности жизни мутанта hif-1 [39, 40]. Мы использовали условия, в которых известно, что мутант hif-1 имеет увеличенную продолжительность жизни [41], так что любое наблюдаемое уменьшение продолжительности жизни nuo-6 в результате мутации hif-1 было бы легче интерпретировать.
Мы обнаружили, что снижение экспрессии hsp-6 , gst-4 , sodh-1 , cdr-2 и ldh-1 не влияло на выживаемость nuo-6 . или черви дикого типа (рис. 7a, b, e, h, i). Напротив, мы обнаружили, что нокдаун mtl-1 , fmo-2 или aldo-1 значительно сокращает продолжительность жизни nuo-6 , но не влияет на продолжительность жизни дикого типа (рис. 7d, g. , к). Аналогичным образом, двойные мутанты nuo-6; sod-3 и nuo-6; hif-1 показали уменьшенную продолжительность жизни по сравнению с мутантами nuo-6 (рис.7в, е). В совокупности это демонстрирует, что изменения в экспрессии генов стрессовой реакции ( hif-1 , mtl-1 , fmo-2 , sod-3 ) и генов, участвующих в метаболизме ( aldo-1 ) которые индуцируются ATFS-1, способствуют продолжительности жизни митохондриальных мутантов nuo-6 и .
Рис. 7Гены, регулируемые ATFS-1, участвующие в стрессоустойчивости и метаболизме, необходимы для долгой жизни мутантов nuo-6 .Чтобы изучить вклад генов, которые демонстрируют ATFS-1-зависимую активацию в мутантах nuo-6 , мы снизили экспрессию этих генов у червей nuo-6 . Мы обнаружили, что уменьшение экспрессии hsp-6 ( a ), gst-4 ( b ), sodh-1 ( e ), cdr-2 ( h ) , ldh-1 ( i ) или acs-2 ( k ) с использованием RNAi не уменьшали продолжительность жизни червей nuo-6 .Напротив, мутация hif-1 ( c ), mtl-1 RNAi ( d ), sod-3 мутация ( f ), fmo-2 RNAi ( g ) , и aldo-1 RNAi ( j ) значительно сокращали продолжительность жизни nuo-6 . Это указывает на то, что изменения в экспрессии генов, вызванные активацией mitoUPR, способствуют увеличению продолжительности жизни червей nuo-6 . Указывается значение между красной и фиолетовой линиями.Необработанные данные о сроке службы см. В дополнительном файле 4: Таблица S4. См. Дополнительный файл 5: Таблица S5 для получения подробной информации о N и репликах
Поскольку мутация в hif-1 полностью отменила увеличение продолжительности жизни, присутствующее у червей nuo-6 , так что nuo-6; hif-1 двойные мутанты живут короче, чем черви дикого типа (рис. 7c), мы решили дополнительно изучить роль hif-1 в продолжительности жизни nuo-6 . Для этого мы использовали RNAseq для сравнения экспрессии генов в мутантах nuo-6 и nuo-6; hif-1 , чтобы идентифицировать гены, которые модулируются у червей nuo-6 и восстанавливаются до дикого типа с помощью мутация hif-1 , поскольку эти гены могут способствовать долгой жизни червей nuo-6 .Используя этот подход, мы обнаружили множество генов с повышенной или пониженной регуляцией у червей nuo-6 и , которые зависят от HIF-1 (дополнительный файл 1: Рисунок S15). Хотя углубленное изучение этих целей выходит за рамки нашего текущего исследования, наши первоначальные анализы с обогащением определили гены, участвующие в различных типах метаболизма, как высшие категории активируемых генов, которые реверсируются мутацией hif-1 , в то время как транспорт Гены / traffic обогащены среди генов с пониженной регуляцией (дополнительный файл 1: рисунки S16, S17).
ATFS-1 влияет на экспрессию генов-мишеней DAF-16
Поскольку мы наблюдали ATFS-1-зависимую активацию трех генов-мишеней DAF-16 у мутантов nuo-6 , мы решили продолжить изучение роли ATFS -1 при активации генов-мишеней DAF-16. Для этого мы исследовали влияние делеции atfs-1 на ядерную локализацию DAF-16 с использованием трансляционного репортерного штамма DAF-16 ( zIs356 [Pdaf-16 :: daf-16: GFP] ) [ 42]. Мы обнаружили, что у мутантов с делецией atfs-1 ядерная локализация DAF-16 в ответ на тепловой стресс 35 ° C была значительно замедлена (дополнительный файл 1: рисунок S18), что позволяет предположить, что ATFS-1 может способствовать ядерной локализации DAF. -16.
Поскольку потеря ATFS-1 повлияла на ядерную локализацию DAF-16, мы затем задались вопросом, в какой степени активация ATFS-1 вызывает модуляцию генов-мишеней DAF-16. Ранее мы показали, что изменения транскрипции, присутствующие в мутантах nuo-6 , имеют значительное перекрытие с мутантами daf-2 и значительное обогащение генов-мишеней DAF-16, и что DAF-16 необходим для продолжительной жизни . nuo-6 [37]. Чтобы более точно оценить роль ATFS-1 в модуляции генов-мишеней DAF-16, мы использовали RNAseq для изучения экспрессии генов в двух мутантах atfs-1 с усилением функции.Эти два мутанта, atfs-1 (et15) и atfs-1 (et17) , имеют точечные мутации, влияющие на митохондриальную целевую последовательность, что приводит к цитоплазматическому накоплению ATFS-1, позволяя ATFS-1 проникать в ядро и активировать его транскрипционные мишени [43].
Мы обнаружили, что гены с повышенной и пониженной регуляцией в мутантах atfs-1 с усилением функции демонстрировали значительную степень перекрытия с мутантами daf-2 (e1370) (рис. 8a, дополнительный файл 1: таблица S1). ).Кроме того, многие из генов-мишеней DAF-16 с максимальной активностью и подавлением, определенные с помощью мета-анализа данных экспрессии daf-16 [44], аналогичным образом модулировались в мутантах atfs-1 с усилением функции (рис. . 8b). Хотя для изучения молекулярных механизмов потребуются дальнейшие исследования, эти результаты предполагают, что ATFS-1 может влиять на гены-мишени DAF-16, по крайней мере, частично через ядерную локализацию DAF-16.
Рис. 8Изменения экспрессии генов при усилении функции мутанты atfs-1 обнаруживают значительное перекрытие с мутантами daf-2 и обогащение генов-мишеней DAF-16.Диаграммы a Венна показывают перекрытие в дифференциально экспрессируемых генах между двумя мутантами atfs-1 по усилению функции ( et15 и et17 ) и daf-2 (e1370). Изменения в экспрессии генов в atfs-1 ( et17) почти полностью перекрываются с изменениями в atfs-1 (et-15). Проценты указывают процент всех генов, которые по-разному экспрессируются в одном из трех мутантов. b Тепловые карты, показывающие экспрессию генов-мишеней DAF-16 в мутантах с усилением функции atfs-1 по сравнению с daf-2. Показанные гены являются главными генами-мишенями DAF-16, определенными метаанализом данных экспрессии генов, выполненным Tepper et al., 2013. Показаны только гены, которые показали значительное изменение экспрессии у мутантов daf-2 . atfs-1 -мутанты с усилением функции демонстрируют повышенную регуляцию многих из лучших генов с повышенной активностью DAF-16 и подавление многих из основных генов с пониженной регуляцией DAF-16. Однако некоторые гены демонстрируют противоположный паттерн регуляции.
ATFS-1-зависимые изменения транскрипции у
nuo-6 мутантовПоскольку мы идентифицировали множественные ATFS-1-зависимые изменения в экспрессии генов у червей nuo-6 , которые способствовали долголетия, мы решили использовать RNAseq, чтобы получить более полное представление о том, какие изменения транскрипции у мутантов nuo-6 зависят от ATFS-1 (см. дополнительный файл 2: Таблица S2 для дифференциально экспрессируемых генов).Мы идентифицировали многие гены, которые были активированы или подавлены у червей nuo-6 и восстановлены до дикого типа с помощью делеции atfs-1 (дополнительный файл 1: Рисунок S19). Хотя для оценки вклада этих генов в долголетие nuo-6 и потребуются дальнейшие исследования, наш первоначальный анализ обогащения показывает, что гены, участвующие в метаболизме липидов, являются наиболее обогащенным классом активированных генов, который зависит от ATFS-1, тогда как гены В метаболизме РНК участвует наиболее обогащенный класс генов с пониженной регуляцией, который зависит от ATFS-1 (дополнительный файл 1: рисунки S20, S21).Соответствуют нашим результатам, показывающим, что роль ATFS-1 заключается в устойчивости к стрессу, устойчивость к стрессу была одним из терминов GO, определенных для ATFS-1-зависимых усиленных генов у мутантов nuo-6 (дополнительный файл 3: таблица S3).
Показав, что черви nuo-6 проявляют активацию mitoUPR и большое количество ATFS-1-зависимых изменений в экспрессии генов, мы сравнили экспрессию генов мутантов nuo-6 с atfs-1 мутанты с усилением функции, atfs-1 (et15) и atfs-1 (et17) .Мы обнаружили, что существует весьма значительная степень перекрытия между мутантами nuo-6 и мутантами atfs-1 с усилением функции (рис. 9). Мы также наблюдали весьма значительную степень перекрытия между мутантами clk-1 и isp-1 и мутантами atfs-1 с усилением функции (дополнительный файл 1: рисунки S22, S23). В совокупности это предполагает, что большая часть изменений транскрипции у долгоживущих митохондриальных мутантов вызывается активацией ATFS-1.
Рис. 9atfs-1 -мутанты с усилением функции демонстрируют очень значимое перекрытие изменений экспрессии генов с мутантами nuo-6 . Диаграммы Венна a указывают на перекрытие в дифференциально экспрессируемых генах между двумя мутантами atfs-1 с усилением функции ( et15 и et17 ) и nuo-6 (qm200) червями . Проценты указывают процент всех генов, которые по-разному экспрессируются в одном из трех мутантов. b Как для генов с повышенной, так и с пониженной регуляцией степень перекрытия очень значима
Последовательности паузы облегчают переход в долгоживущие состояния паузы за счет снижения скорости транскрипции РНК-полимеразы
Реагенты
Все ДНК-модифицирующие ферменты были приобретены в Новой Англии Биолаборатории. Олигонуклеотиды были приобретены у IDT. Нуклеотидтрифосфаты были приобретены у Thermo Scientific, а стандартные соли и компоненты буфера были приобретены у Sigma Aldrich.Карбоксилированные гранулы полистирола 1 мкм были приобретены в Bangs Labs. Бактериальные штаммы для экспрессии белков и размножения плазмид были приобретены у EMD Millipore.
Плазмиды и шаблоны ДНК
Плазмиды pIA1127 (для экспрессии сигма 70), pIA1234 (для экспрессии сортированной РНК-полимеразы) и pIA2–6 (используемые в качестве матрицы для изготовления ручек ДНК) были подарком Ирины Арцимович (Огайо). Государственный университет, кафедра микробиологии). Матрица (8XHis) была получена из плазмиды, содержащей промотор T7A1, кассету без U размером 29 п.н., спейсерную область размером ~ 1 т.п.н., восемь повторов, содержащих паузу ‘his’, и, наконец, последовательность терминатора rrnB T1 13 .Плазмида для экспрессии сортазы была подарком Дэвида Лю (Гарвардский университет, факультет химии и химической биологии).
Олигонуклеотиды, использованные в этом исследовании
Для получения ручек размером 1,5 т.п.н. из плазмиды pIA2-6 были использованы следующие олигонуклеотиды:
Фор-биотин: 5 ‘/ 5Biosg / GAAAGTCCGGCATCTCAATCCC 3’
G ‘-B GAGACC GATGTGGCTTCGGTCCCTTC 3 ‘
В Rev-BsaI подчеркнутые основания обозначают сайт узнавания BsaI, который после расщепления образует 5′-выступ ACCG.
Рукоятка была подготовлена с помощью ПЦР и очищена стандартной очисткой ПЦР, обработана BsaI-HF (5 единиц на мкг ДНК в течение 2 ч при 37 ° C с последующей тепловой инактивацией в течение 20 мин при 65 ° C) и очищена с помощью очистки с помощью ПЦР.
Для модификации поверхности гранул были заказаны следующие олигонуклеотиды, очищенные с помощью ВЭЖХ и использованные в полученном виде:
Амин гранул: 5 ‘/ 5AmMC6 / TTAATTCATTGCGTTCTGTACACG 3’
Bead CGGTT: 5 ‘/ 5Phos / CGGAGCGT000: 5’ / 5Phos / CGGAGCT000 5 ‘/ 5Phos / ACCGCGTGTACAGAACGCAATGAATT 3’
Получение ДНК-матрицы
Для приготовления ДНК-матрицы плазмиды 8XHis рестриктировали BsaI-HF (8 единиц на мкг ДНК в течение 2 часов при 37 ° C) и обрабатывали параллельно щелочная фосфатаза креветок (0.4 единицы на мкг ДНК) для создания линейной ДНК с четкими дефосфорилированными 5′-выступами. Ферменты деактивировали нагреванием в течение 20 минут при 65 ° C, и ДНК немедленно обрабатывали 3′-5′-экзополимеразой Кленова (1 единица на мкг ДНК) и 0,1 мМ ddATP (для создания матрицы вспомогательной силы) или 0,1 мМ. mM ddCTP (для создания шаблона противостоящей силы). Реакция протекала в течение 30 мин при 37 ° C с последующей тепловой инактивацией в течение 20 мин при 75 ° C. Затем образец пять раз экстрагировали фенол-хлороформом и один раз хлороформом, осаждали этанолом и восстанавливали в Трис 10 мМ pH = 8,0.1 мМ ЭДТА. Чистота ДНК составила ~ 88% с помощью электрофореза в агарозном геле.
Сочетание бусинок с олигонуклеотидами
Для получения двухцепочечного олиго для связывания олигонуклеотид Bead Amine был гибридизован с олиго CGGT на бусах или олиго Bead ACCG с образованием двухцепочечного олиго, содержащего фосфорилированный 5′-выступ. Отжиг выполняли путем нагревания смеси олигонуклеотидов в воде (0,25 мМ каждый) в соотношении 1: 1 до 95 ° C в течение 10 мин с последующим охлаждением до комнатной температуры на стенде.Это привело к получению двухцепочечных олигонуклеотидов, несущих аминогруппу на одном конце и либо 5′-выступающий выступ CGGT (для экспериментов с вспомогательной силой), либо 5′-выступающий выступ ACCG (для экспериментов с противодействующей силой) на другом конце.
Гранулы карбоксилированного полистирола диаметром 1 мкм (Bangs Labs) были присоединены к полученным двухцепочечным олигонуклеотидам следующим образом: 10 мкл 10% (вес / объем) гранул промыли 4 раза 200 мкл буфера для связывания (MES 0,1 M pH = 4.7, 150 мМ NaCl, 5% ДМСО) и диспергировали в 20 мкл буфера для связывания.Все центрифугирования проходили в течение 5 мин при 4500 g. Добавляли 10 мкл 20 мкМ двухцепочечного олиго и 6 мкл 2 М 1-этил-3- (3-диметиламинопропил) карбодиимида (EDC) с последующим энергичным встряхиванием в течение 2 часов при комнатной температуре. В этот момент добавляли еще 10 мкл 2 М EDC с последующим встряхиванием в течение ночи при комнатной температуре.
Затем оставшийся активный EDC гасили добавлением 2,5 мкл 1 М глицина, и шарики промывали 5 раз буфером для хранения (Трис 20 мМ pH = 8, 1 мМ EDTA, 0.05% Tween 20, 5 мМ азид натрия) с 3-минутной обработкой ультразвуком между промывками. В конце гранулы диспергировали до концентрации 1% (мас. / Об.) И хранили при 4 ° C.
Пассивация гранул
Гранулы пассивировали 6-кратным разведением в ТЕ (Трис 20 мМ, pH = 8, 1 мМ ЭДТА) и добавлением β-казеина до концентрации 1 мг / мл. Гранулы встряхивали в течение 10 мин, один раз промывали ТЕ, диспергировали до концентрации 0,2% в ТЕ и хранили при 4 ° C до эксперимента.
Приготовление сигма 70
Плазмида pIA1127 была трансформирована в бактерии Rosetta2.Бактерии выращивали в 2 литрах среды 2YT с добавлением 1% глюкозы, NPS (25 мМ (NH 4 ) 2 SO 4 , 50 мМ KH 2 PO 4 , 50 мМ Na 2 HPO 4 ), 1 мМ сульфата магния, 34 мкг / мл хлорамфеникола и 50 мкг / мл канамицина. Культуру выращивали при 37 ° C до OD600 0,5, переносили до 17 ° C и добавляли IPTG до 0,1 мМ. Индукция продолжалась 16 ч.
Для очистки бактерии диспергировали в 80 мл буфера A25 (Трис 20 мМ, pH = 7.5, 0,5 М NaCl, 10% глицерин, 25 мМ имидазол, 2 мМ бета-меркаптоэтанол) с добавлением 0,1 мг / мл лизоцима и ингибиторов протеаз (Roche). Бактерии лизировали с помощью французского пресса, лизат осветляли центрифугированием и фильтрацией и загружали в колонку с Ni-NTA объемом 5 мл. Колонку промывали 20 мл буфера A25 и 20 мл A50 (A25 + 50 мМ имидазол), и сигма 70 с гист-меткой элюировали в A300 (A25 + 300 мМ имидазол). TEV-протеазу (выраженную в BL21 и очищенную с использованием смолы Ni-NTA с последующей эксклюзионной хроматографией 40 ) добавляли в молярном соотношении 1:40, и реакция продолжалась в течение ночи при 4 ° C, подвергаясь диализу против A50.Затем образец снова пропускали через колонку Ni-NTA. Проточный поток, содержащий сигму 70 без гистага, собирали, концентрировали в два раза и дополнительно очищали гель-фильтрацией на колонке с сефакрилом S300, уравновешенной буфером B (Трис 20 мМ pH = 7,5, 0,5 М NaCl, 10% глицерин, 1 мМ ЭДТА, 1 мМ ДТТ). Протокол давал ~ 35 мг чистой сигма 70. Аликвоты мгновенно замораживали в жидком азоте и хранили при -80 ° C.
Получение сортируемого холофермента РНК-полимеразы
Плазмиду pIA1234 трансформировали в бактерии Rosetta2.Sortag-RNAP экспрессировали с использованием того же протокола, что и сигма 70, за исключением того, что вместо канамицина использовали ампициллин.
Для очистки использовалась модифицированная версия опубликованного протокола 41 . Клетки диспергировали в 75 мл буфера для лизиса (Трис 50 мМ pH = 6,9, 0,5 М NaCl, 5% глицерин) с добавлением 0,1 мг / мл ингибиторов лизоцима и протеаз и лизировали во французской прессе. Лизат центрифугировали и фильтровали, и добавляли имидазол до 20 мМ. Образец загружали в колонку Ni-NTA объемом 5 мл.Колонку промывали 30 мл буфера для лизиса + 20 мМ имидазола, и ядерный фермент RNAP с гист-меткой элюировали буфером для лизиса + 250 мМ имидазол.
Для образования холофермента образец инкубировали с 2-кратным избытком очищенного сигма 70 в течение ночи на льду. Образец разбавляли в десять раз буфером B0 (50 мМ Трис pH = 6,9, 5% глицерин, 0,5 мМ EDTA, 1 мМ DTT) и наносили на колонку с гепарином 5 мл. Чтобы избежать перегрузки колонки, образец был разделен на три части, которые загружались отдельно.Для элюирования белка использовали градиент от 50 мМ до 1 М NaCl. Холофермент RNAP был четко отделен от избытка сигма 70. Образец диализовали против буфера B50 (50 мМ Tris pH = 6,9, 5% глицерин, 50 мМ NaCl, 0,5 мМ EDTA, 1 мМ DTT), а затем очищали в 1 мл monoQ с использованием градиента NaCl от 50 мМ до 1 М (опять же, образец был разделен на три порции, загруженные отдельно). Чистые фракции объединяли, диализовали против буфера для хранения (20 мМ Трис pH = 7,5, 200 мМ KCl, 0,2 мМ EDTA, 0,2 мМ DTT, 5% глицерин), разделяли на аликвоты, быстро замораживали и хранили при -80 ° C.
Биотинилирование sortag-RNAP
Мы получили пептид, содержащий N-концевую GGG-метку с модифицированным биотином остатком лизина (Genscript): GGGGDGDY {Lys (биотин)}. 100 мкл 9,6 мкМ sortag-RNAP реагировали с 200-кратным избытком биотинилированного пептида в 200 мкл буфера для связывания (Tris 50 мМ pH = 7,5, 5 мМ CaCl 2 ), содержащего 2 мкМ сортазы (экспрессированной в бактериях BL21 и очищенной с использованием смолы Ni-NTA с последующей эксклюзионной хроматографией 42 ). Реакция продолжалась 60 мин.На этом этапе добавляли имидазол до 25 мМ и NaCl до 350 мМ, и образец пропускали через 70 мкл гранул Ni-NTA для удаления сортазы и непрореагировавшего RNAP. Пептид удаляли диализом в буфер для хранения, и биотинилированный RNAP хранили в буфере для хранения при -80 ° C.
Получение GreB
Ген GreB клонировали в вектор pET путем клонирования, не зависящего от лигирования (Addgene # 29653). Плазмиду трансформировали в клетки Rosetta 2, бактерии выращивали в 1 литре среды 2YT с добавлением 1% глюкозы, NPS (25 мМ (NH 4 ) 2 SO 4 , 50 мМ KH 2 PO 4 , 50 мМ Na 2 HPO 4 ), 1 мМ сульфата магния, 34 мкг / мл хлорамфеникола и 50 мкг / мл канамицина.Культуру выращивали при 37 ° C до OD600 0,6, добавляли IPTG до 0,5 мМ и трансформацию продолжали в течение 4 часов при 37 ° C. Затем бактерии центрифугировали и диспергировали в 40 мл буфера для лизиса (Трис 100 мМ pH = 7,9, 25 мМ имидазол, 1 М NaCl, 2 мМ бета-меркаптоэтанол) с добавлением 1 мМ PMSF и 0,2 мг / мл лизозима. Бактерии лизировали обработкой ультразвуком, раствор центрифугировали и фильтровали.
Образец загружали в колонку Ni-NTA объемом 2 мл, промывали 12 мл буфера для лизиса, затем 12 мл буфера для лизиса с 50 мМ имидазолом и, наконец, элюировали буфером для лизиса с 300 мМ имидазолом.Добавляли протеазу TEV в молярном соотношении 1:10, и образец инкубировали в течение ночи при 4 ° C при диализе против буфера для лизиса. Образец снова пропускали через гранулы Ni-NTA объемом 1 мл, концентрировали до <3 мл и загружали в колонку для гель-фильтрации с сефакрилом S100, уравновешенную трис 25 мМ pH = 8, 1 М NaCl, 1 мМ EDTA, 1 мМ DTT. Фракции, содержащие чистый GreB, объединяли и концентрировали до ~ 50 мкМ; глицерин был добавлен до 50%; и белок мгновенно замораживали жидким азотом и хранили при -80 ° C.
При проведении экспериментов с GreB белок сначала диализовали в HEPES 25 мМ pH = 8, 1 M KCl, 1 мМ DTT и 1 мМ EDTA, чтобы его можно было смешать с экспериментальным буфером в предварительно рассчитанных соотношениях, чтобы поддерживать буферный состав.
Все белки имели чистоту> 95% на основании SDS – PAGE. Активность и паузу Holo-RNAP подтверждали с использованием короткой матрицы, содержащей промотор T7A1, кассету без U из 29 пар оснований и расположенный ниже «his» сайт. Активность GreB тестировали путем восстановления комплекса элонгации с обратным прослеживанием длиной 2 п.н., собранного с использованием олигонуклеотида РНК с двумя несовпадающими основаниями на 3′-конце 25 .
Получение остановленных комплексов и сборки связки
Остановленные комплексы были получены путем инкубации 2 нМ ДНК с 10 нМ РНКП в TB20 (Трис 20 мМ pH = 8, 20 мМ NaCl, 20 мМ DTT, 10 мМ MgCl 2 , 20 мкг / мл казеина) в течение 20 мин при 37 ° C.
Остановленные комплексы затем лигировали с шариками при соотношении 1 фмоль остановленного комплекса к 2 мкг шариков в ТВ20 в присутствии 0,1 мМ АТФ и 0,4 единиц ДНК-лигазы Т4 в течение 60 мин при комнатной температуре. Для ручек ДНК размером 1,5 т.п.н. 1 фмоль ручки лигировали с 3 мкг шариками.
После лигирования к шарикам добавляли гепарин до 0,4 мг / мл. К шарикам, прикрепленным к ручке ДНК, добавляли 200-кратный избыток нейтравидина и инкубировали с шариками в течение 10 мин перед разбавлением экспериментальным буфером. Для остановленных комплексных шариков шарики инкубировали в течение 10 мин с добавленным гепарином перед разбавлением экспериментальным буфером.
Конструкция прибора
Эксперименты проводились на установке оптического пинцета с разделением времени, модифицированной по сравнению с конструкцией Комстока и др. 43,44 . В этой конфигурации лазер с длиной волны 1064 нм проходит через акустооптический дефлектор, при этом положение лазера между двумя ловушками меняется каждые 5 мкс. Положение шариков относительно ловушек было измерено с помощью интерферометрии в задней фокальной плоскости 45 .
Экспериментальная установка
Экспериментальный буфер представлял собой: HEPES 50 мМ pH = 8, 130 мМ KCl, 4 мМ MgCl 2 , 0,1 мМ DTT, 0,1 мМ EDTA, 20 мкг / мл гепарина и 10 мМ NaN 3 ( добавлен в качестве поглотителя синглетного кислорода для снижения степени фотоповреждения ( 46 ).Концентрации NTP составляли 1 мМ UTP, 1 мМ GTP, 0,5 мМ АТФ и 0,25 мМ CTP 18 . При добавлении к раствору NTP концентрации GreB и РНКазы A составляли 0,87 мкМ и 0,1 мг / мл соответственно. Для проведения эксперимента использовалась установка с ламинарным потоком 21 . Основной канал камеры был образован потоком, выходящим из резервуара, содержащего буфер, и потоком из второго резервуара, содержащего раствор NTP. Два потока образуют в камере хорошо разделенные области. Гранулы, содержащие ручку ДНК, загружали в боковой канал, соединенный со стороной NTP через тонкий капилляр, в то время как шарики, содержащие остановленный комплекс, загружали в аналогичный канал, соединенный со стороной буфера.Каждый эксперимент состоял из следующих шагов:
1. Захват шариков ручки ДНК на стороне NTP.
2. Захват застывшей сложной бусины на стороне буфера.
3. Потирайте бусинки друг о друга со стороны буфера до тех пор, пока не сформируется трос (если вообще).
4. Если привязка сформирована и имеет ожидаемую длину, перемещение пары на сторону NTP для перезапуска транскрипции.
Наши эксперименты проводились с использованием активного силового зажима, который постоянно перемещал одну из ловушек для поддержания постоянной средней силы.Это позволяло поддерживать силу в пределах 0,1 пН в пределах каждой трассы. Однако мы обнаружили, что небольшие изменения в калибровке могут возникать из-за того, что калибровка и измерение активности не выполнялись в одном и том же месте в камере, возможно, из-за изменений толщины стекла или изменения показателя преломления из-за разный состав буфера в канале NTP. Поэтому после разрыва троса мы выполнили дополнительную калибровку в канале NTP и использовали ее для расчета сил.Очевидно, что обратная связь все же должна была осуществляться с использованием параметров калибровки, измеренных в буферном канале. Это привело к небольшому отклонению измеряемой силы от привязи к привязи, которая редко превышала 0,5 пН. Для каждой трассы сбор данных прекращался после разрыва привязки (преждевременно или из-за разрыва связи) или после очень долгих (> 200 с) пауз.
Доступность данных
Данные, подтверждающие выводы этой рукописи, доступны у соответствующего автора по разумному запросу.
Визуализация динамики факторов транскрипции в живых клетках | Журнал клеточной биологии
Примерно 2 м (приблизительно шесть миллиардов пар оснований) линейной ДНК должны быть упакованы в ядро каждой диплоидной клетки человека (~ 5–10 мкм в диаметре). При такой же плотности упаковки можно поместить цепь ДНК, превышающую длину окружности Земли в 6000 раз внутри куриного яйца. Эффективная упаковка ДНК в ядро клетки частично достигается за счет наматывания ДНК вокруг гистонов в нуклеосомы.В настоящее время у нас есть кристаллические структуры нуклеосомы (Luger et al., 1997). На основании структурных исследований in vitro (Schalch et al., 2005; Woodcock and Ghosh, 2010; Song et al., 2014) было предложено, что нуклеосомные полимеры могут собираться в 30-нм волокна. Недавно был разработан новый краситель DNA EM для детального изображения организации волокон хроматина с высоким разрешением в отдельных фиксированных клетках. Эти исследования выявили очень неупорядоченные структуры, и волокна размером ~ 5-24 нм были обнаружены in situ (Ou et al., 2017), указывая тем самым, что 30-нм волокно, вероятно, не является доминирующей формой хроматина в ядре.Другой результат этого исследования предполагает, что трехмерные субядерные домены собраны с разной плотностью хроматина, и эта особенность, вероятно, определяет глобальную доступность и активность ДНК в ядре.
В настоящее время большие субядерные компартменты, такие как гетерохроматин, ядерные спеклы, тельца Кахаля, кластеры гистоновых локусов и нуклеоиды, могут быть визуализированы в живых клетках с помощью световой микроскопии (Mao et al., 2011). Однако из-за высокой плотности упаковки нативных волокон хроматина и отсутствия неинвазивных инструментов для мечения специфических локусов генов, структурная информация, лежащая в основе коммуникации энхансер-промотор в клетке, остается недоступной для традиционной структурной биологии или световой микроскопии.Текущая информация о потенциальных структурах, управляющих коммуникацией энхансер-промотор, в основном получена с помощью непрямых методов, таких как анализы на основе захвата конформации хромосомы (3C) (de Wit and de Laat, 2012). В частности, химически сшитые волокна хроматина перевариваются рестрикционными ферментами, и частота контактов между фрагментами хроматина определяется на основе реакций лигирования с близостью, проводимых со смесями больших популяций клеток. В последнее десятилетие обширные полногеномные исследования (4C, 5C и Hi-C) предоставили модель сворачивания хроматина, топологических доменов и потенциальной трехмерной организации генома (Dekker et al., 2013; Деккер, Мирный, 2016). Двумя основными особенностями, извлеченными из этих исследований, являются: (1) как было предложено первоначально из самых ранних исследований энхансеров (Su et al., 1991), энхансеры и промоторы, вероятно, связываются друг с другом через петли хроматина, и (2) геном организован. в топологически ассоциированные домены (TAD), которые могут разделять активность локальных генов. Однако данные визуализации и геномики иногда показывают значительные расхождения (Belmont, 2014; Williamson et al., 2014; Boettiger et al., 2016; Wang et al., 2016b), предполагая, что анализы на основе 3C могут иметь ограниченную способность фиксировать истинные трехмерные структуры и физическую близость элементов в отдельных живых клетках. Существует также дополнительная возможность того, что химическое сшивание может привести к некоторым непредвиденным последствиям, которые нелегко предвидеть или интерпретировать. Например, недавние исследования SMT показывают, что в живых клетках Sox2 взаимодействует с митотическими хромосомами для создания закладок (Deluz et al., 2016; Teves et al., 2016), тогда как наиболее поразительно то, что химическое перекрестное связывание приводит к исключению Sox2 из митотические хромосомы (Teves et al., 2016). Кроме того, недавнее исследование CTCF и когезина с SMT показало, что образование петли хроматина может быть высокодинамичным (~ мин) и регулируемым событием в клетке (Hansen et al., 2017a). Результаты таких визуализационных исследований с использованием живых клеток показывают, что частота взаимодействия, обнаруженная с помощью анализов 3C, с большей вероятностью отражает сложную свертку физической близости, частоты петель ДНК, локальной среды хроматина и клеточной гетерогенности (Fudenberg and Imakaev, 2017). В соответствии с этим представлением, эксперименты с Hi-C одной ячейкой показывают, что стохастические кластеры контактов могут происходить через границы TAD в отдельных ячейках, но усредняются в TAD в ансамблевых анализах (Flyamer et al., 2017), предполагая, что TAD могут быть результатом вычислительной нормализации событий перекрестного связывания в миллионах ячеек.
Несмотря на некоторую возможность для дискуссий, данные визуализации живых клеток сходятся с геномными исследованиями присутствия топологических структур в ядре, которые, вероятно, также формируют локальные регуляторные активности генов. Какова взаимная связь между этими топологическими структурами и пространственным распределением различных ТФ в ядре? Путем визуализации долгоживущих событий связывания Sox2 в живых клетках с помощью решеточной световой микроскопии стало возможным систематически картировать организацию 3D-энхансера Sox2 в ES клетках.Было обнаружено, что стабильные сайты связывания Sox2 (наиболее вероятно, энхансеры) образуют пространственно ограниченные кластеры в ядре живых ES клеток (Liu et al., 2014). Эти кластеры энхансеров пространственно отделены от гетерохроматических областей, но перекрываются с подмножеством кластеров, обогащенных Pol II (Fig. 3 A). Кроме того, эксперименты с SMT показали, что внутри кластеров энхансеров Sox2 демонстрирует значительно более высокие скорости прямой ассоциации (k на ), тем самым увеличивая локальные концентрации TF и обеспечивая быстрое повторное связывание TF с участками открытого хроматина (рис.3 Б). В соответствии с этим ранним наблюдением SMT-эксперименты подтвердили, что TF также образует кластеры в клетках дрожжей и Drosophila и что кластеризация TF, вероятно, регулирует транскрипционный выход из нижележащих генов (Mir et al., 2017; Wollman et al., 2017). Кроме того, подробный анализ долгоживущих событий связывания Sox2 на уровне одной молекулы показал, что Sox2 прыгает между кластерными сайтами связывания в пространственно ограниченных субядерных регионах (Liu et al., 2018). Эти результаты подтверждают существование определенных топологических структур и концентраторов TF в ядре, которые могут формировать локальную динамику поиска мишеней для TF и потенциально точно настраивать скорости сборки комплекса TF на цис-регуляторных элементах.
Какие силы удерживают вместе такие центры цис-регулирующих элементов? С самого раннего клонирования и характеристики классических последовательностей ТФ возникла загадочная особенность: открытие простых повторяющихся, в значительной степени неструктурированных аминокислотных мотивов (например, кислотных повторов, богатых глицином и пролином), которые служат в качестве связанных доменов активации (AD). с ДНК-связывающими доменами (Courey and Tjian, 1988). Более поздние данные свидетельствуют о том, что такие простые повторяющиеся аминокислотные мотивы, теперь называемые доменами низкой сложности (LCD), широко распространены в различных регуляторных белках, включая многие классические TF и другие белки, такие как Fused in саркома (FUS ), TAF15 и белок саркомы Юинга (EWS; Kwon et al., 2013) и могут участвовать в формировании динамических фазово-разделенных компартментов в живых клетках (Patel et al., 2015; Shin, Brangwynne, 2017). Мы предполагаем, что кластеризация энхансеров in vivo или «образование концентраторов», наблюдаемое в исследованиях живых клеток, открывает возможность того, что высокие локальные концентрации TF могут быть результатом образования кластеров ЖКД, по крайней мере временно. Т.о., возможно, что эти энхансерные кластеры могут служить поливалентными стыковочными сайтами для динамического рекрутирования TF посредством слабых межбелковых взаимодействий, потенциально управляемых с помощью LCD-содержащих белков.В свою очередь, также возможно, что такие слабые межбелковые взаимодействия усиливают и вносят вклад в установление кластеризации цис-регуляторных элементов. В самом деле, LCDs в AD Sp1 и Foxp2 могут динамически взаимодействовать с массивами PolyQ на поверхности агрегатов мутантного хантингтина в живых клетках (Li et al., 2016). Сходным образом мы можем представить, что таким центрам слабых мультивалентных белок-белковых взаимодействий внутри энхансерных кластеров могут способствовать более сильные последовательности-специфичные транзакции белок-ДНК, которые вместе влияют на локальную среду хроматина, чтобы регулировать активность генов.Интересно, что структурный анализ in vitro восстановленного Mig1 указывает на то, что кластеры TF в ядре дрожжей стабилизируются за счет взаимодействий между LCDs, подчеркивая критическую роль LCD в обеспечении образования кластеров TF (Fig. 3 C). В соответствии с повышенной скоростью прямой ассоциации для Sox2 в кластере энхансеров предполагается, что образование этих кластеров TF в дрожжах сокращает время поиска промотора за счет переноса между сегментами при стабилизации экспрессии гена (рис. 3D; Wollman et al., 2017). Интересная теоретическая работа также предполагает, что в присутствии TF ДНК может подвергаться фазовому разделению (Le Treut et al., 2016). В соответствии с этой моделью концентратора TF, другие хорошо описанные разделенные по фазе структуры, известные как ядерные тельца (например, ядерные спеклы, белковые тельца промиелоцитарного лейкоза и тельца Кахаля), как полагают, способны влиять на организацию генома путем секвестирования генов-мишеней в специализированных микросредах. (Brown et al., 2008; Ching et al., 2013; Wang et al., 2016а). Моделирование геномных данных на основе клеточной популяции также предполагает кластеризацию и организацию цис-регуляторных элементов более высокого порядка в ядре (Canals-Hamann et al., 2013; Hnisz et al., 2013, 2017; Dai et al., 2016; Beagrie et al., 2017). Однако эти исследования и гипотетические модели не предоставляют информацию о пространственном аспекте кластеров на уровне одной клетки, одной молекулы. Несмотря на появляющиеся доказательства кластеризации TF, важно отметить, что взаимосвязь между организацией генома, позиционированием генов и активностью генов четко не установлена.С одной стороны, было показано, что расположение гена в ядре может быть потенциально критическим для опосредования активности генов. Например, на про-В-клетках и мышечных клетках было показано, что активация или репрессия конкретных генов связана с транслокацией генов в новые места (Kosak et al., 2002; Moen et al., 2004). С другой стороны, геномные данные предполагают, что области, контактирующие с GR-регулируемыми генами, не особенно обогащены сайтами нацеливания GR или какой-либо функциональной группой генов (Hakim et al., 2011), предполагая, что функциональная организация генома скорее всего не отвечает на кластерное связывание индивидуальных TF. Контактные области, однако, обогащены сайтами гиперчувствительности к DNaseI, что указывает на то, что ядро предварительно организовано в конформацию, благоприятную для быстрого репрограммирования транскрипции, и эта организация, вероятно, управляется сайтами хроматина, доступными для различных регуляторных факторов.
Пространственно-скоординированная динамическая транскрипция генов в живой ткани гипофиза
Рецензенты считают работу интересной, в частности, то, что пространственные отношения клеток в ткани могут привести к общей основе для экспрессии.Однако они считают, что есть неясные анализы и текст неоднозначен по частям. Один рецензент «нашел работу несовершенной с точки зрения количественного и теоретического анализа». Другая мысль, что «основным вкладом этой статьи может быть взаимосвязь между этим стохастическим процессом и интегрированным производством из ткани», но это не было четко развито. В обоих случаях потребуются некоторые обширные исправления, в частности, более строгий текст и анализ, прежде чем его можно будет рассматривать дальше.
Мы внесли серию подробных изменений в документ в ответ на эти комментарии и включили новый окончательный рисунок, новый дополнительный рисунок и исправления к исходному рисунку 2 и рисунку 3, чтобы попытаться представить данные более четко и ключевые понятия более лаконично. Мы считаем, что эти изменения значительно улучшили рукопись, и надеемся, что теперь она будет принята для публикации в eLife . Наши подробные ответы перечислены ниже.
Рецензент № 1:
1) Авторы анализируют данные временной флуоресценции, используя теоретическую стохастическую модель, чтобы извлечь параметры активности генов.Однако достоверность модели не подвергается сомнению при проверке ее способности успешно воспроизводить экспериментальные данные. Нигде мы не видим график с прямым сравнением экспериментальной наблюдаемой и ее предсказанного значения на основе модели. Таким образом, у нас есть процесс «только вперед», когда экспериментальные данные вводятся в алгоритм для получения оценочных параметров без критической обратной связи от модели к эксперименту. Это резко снижает ценность теоретического анализа.
Рецензент цитирует статью Halpern et al.как образец для этого вида количественного анализа. Мы согласны с тем, что эта статья представляет собой современную передовую область, и цитируем ее в нашей рукописи. Halpern et al. использовать сложный smFISH, что означает, что динамика транскрипции рассчитывается на основе одного зафиксированного снимка активности. Напротив, наш подход отслеживает динамику транскрипции в отдельных клетках в реальном времени в течение 40-часовых периодов. Подход, который мы использовали, является значительным развитием работы, впервые опубликованной в двух новаторских статьях 2011 года, в которых использовался количественный анализ динамики отдельных ячеек (Suter et al.in Science, и наша предыдущая статья Харпера по биологии PLoS и др.). Ключевой новизной данной работы является анализ кинетики транскрипционного взрыва в зависимости от пространственной близости клеток в интактной живой ткани. При этом используются основные достижения в статистическом анализе транскрипции с несколькими состояниями, основанные на разработке и применении теоретических инструментов, описанных в нашей статье по биостатистике (Hey et al. 2015).
Рецензент, возможно, не оценил, что разработка нашего статистического анализа в основном опиралась на итеративное тестирование модели на синтетических и реальных данных.Наш подход на сегодняшний день является единственным способом сравнения моделирования с истинным лежащим в основе биологическим процессом (при условии, что предполагаемые переключатели действительно являются переключателями в транскрипции). Мы подтвердили, что модель в разумных пределах соответствует экспериментальным данным, итеративно протестированы с помощью рекурсивных остатков. Эти остатки были рассчитаны как для ткани P1.5, так и для ткани взрослого, которые были представлены в статье биостатистики, и мы включаем их для вашего рассмотрения. Эти остатки представляют собой способ сравнения прогноза модели (основанного на оценках) и наблюдаемых данных.Подробная информация об использовании таких остатков содержится в Приложении G статьи по биостатистике (см. Дополнительные материалы, доступные на сайте Biostatistics в Интернете), и никаких отклонений от допущений модели обнаружено не было, что указывает на то, что SSM как в LNA, так и в BDA хорошо соответствует данным .
Мы добавили к рукописи следующее предложение, касающееся этих пунктов: «Соответствие модели было проверено путем вычисления рекурсивных остатков как способа сравнения прогноза модели и наблюдаемых данных (информацию см. В Приложении G к ( 20)).Они не показали отклонения от допущений модели, указывая на то, что SSM хорошо соответствует данным ».
2) В связи с этим неясно, что было извлечено из модели, чего еще не было в экспериментальных данных. В частности, временные и пространственные корреляции в экспрессии и то, как они меняются в процессе развития, очевидны из прямого анализа данных, и не было очевидно, что модель проливает дополнительный свет на эти результаты.
Это критический момент.Экспериментальные данные, на которые ссылается обозреватель, являются необработанными данными экспрессии флуоресцентного белка. Сигнал флуоресценции зависит от множества процессов, включая транскрипцию, трансляцию и деградацию белков и мРНК. Чтобы исследовать корреляцию между клетками, критически важно иметь возможность точно измерить и статистически оценить переключение нескольких состояний в скорости транскрипции в отрыве от других процессов. Обратный расчет скорости транскрипции, который мы предприняли, позволяет нам оценить корреляцию между клетками с точки зрения процесса транскрипции, оценивая корреляцию во временах переключения и направлениях переключения.Подход к моделированию также показал наличие более коротких периодов «включения» в незрелой ткани гипофиза и что сходные распределения скорости транскрипции присутствовали на всех стадиях развития гипофиза. Эти идеи важны для понимания того, как изменения в ответах отдельных клеток приводят к изменениям общего выхода тканевой системы. Важность моделирования динамики транскрипции и выводы, полученные в результате такого анализа, включая подход к моделированию, использованный в нашем исследовании, обсуждались во втором параграфе «Обсуждение».Что касается информативных выводов относительно общего биологического поведения, мы добавили некоторый дополнительный текст к Обсуждению в параграфе шесть и теперь включаем новую имитационную модель и гипотезу относительно природы общего транскрипционного выхода из эндокринной ткани (обрисовано в общих чертах на Рисунке 9).
3) Помимо теоретической модели, количественный анализ данных сам по себе в ряде случаев был весьма некорректным. Примеры:
i) «активность флуоресценции показала явное отклонение от процесса белого шума, что указывает на пульсирующее поведение транскрипции».Почему следует ожидать «белого шума»? И почему отклонение от белого шума указывает на наличие импульсов? Это очень непонятно.
ii) Корреляции между ячейками на Рисунке 2B-ii показывают увеличение на больших расстояниях. Вероятно, это артефакт, поскольку его можно увидеть и в рандомизированном контроле, но наличие этой тенденции подозрительно — может ли это указывать на ошибку в вычислении корреляции, например в нормализации на количество пар ячеек?
Пример i).Рецензент прав, критикуя предложение «Автокорреляционный анализ не выявил регулярный однородный период, но активность флуоресценции показала явное отклонение от процесса белого шума, указывающее на пульсирующее транскрипционное поведение (рис. 1D) с определенной количественной статистической структурой. «Мы согласны с тем, что это вводило в заблуждение, поскольку это, кажется, предполагает, что, поскольку процесс не был процессом белого шума, он, следовательно, должен отражать пульсирующее транскрипционное поведение. Мы должны были сказать:
«Автокорреляционный анализ не выявил регулярного однородного периода, а активность флуоресценции показала явное отклонение от процесса белого шума (рис. 1D).Кроме того, анализ с использованием модели стохастического переключения (SSM) выявил определенную поддающуюся количественной оценке статистическую структуру, указывающую на пульсирующее поведение транскрипции, совместимое с процессом импульсного телеграфа, как описано выше ».
Это предложение было соответствующим образом изменено в рукописи в подзаголовке «Паттерны транскрипционной активности гена пролактина во взрослой ткани гипофиза».
Пример ii) Судья улавливает важный момент, который мы проверили дальше. Корреляции между ячейками на рисунке 2B, на которые ссылается рецензент, находятся внутри 90% доверительного интервала рандомизированного контроля и, следовательно, не являются значимыми.Это дополнительно подчеркивается данными в части i) рисунка, из которых ясно, что количество пар в ячейках выше 70 очень мало. Мы решили эту проблему, заново построив график данных с использованием выбранных интервалов, чтобы все они имели одинаковое количество пар ячеек. Это ясно показывает значительное снижение корреляции для расстояний менее примерно 40 мкм, и мы видим, что последние два интервала уверенно попадают в 90% доверительный интервал рандомизированного контроля. Эта новая информация теперь включена в рисунок 3 — дополнение к рисунку 1.
Рецензент № 2:
1) Моя основная критика касается характера развития, выхода генов и пространственной координации. Авторы представляют серию наблюдений по всем трем, но я полагаю, что было бы более логичным с биологической точки зрения интегрировать эти наблюдения для описания общего выхода пролактина из ткани. Увеличивается ли общий объем производства на этапах разработки? Если да, то является ли этот результат результатом большего количества ячеек (т.е. большей плотности) или изменений в пульсации? Если клетки пульсируют синхронно, будет ли общий вывод также отображать пульсирующую продукцию железой? Заманчиво сделать вывод из данных авторов, что клетки имеют своего рода фиксированную цепь регуляции пролактина, но по мере созревания железы большее количество клеток приводит к более высоким уровням продукции, и этот вывод становится динамически синхронизированным посредством межклеточной коммуникации.Это правильно? Или я неправильно понимаю данные? Здесь количественное моделирование было бы чрезвычайно ценным, возможно, позволяя экстраполировать от срезов ткани к функции in vivo.
Мы согласны с тем, что нам следует более четко предложить биологическое значение и контекст работы. Мы попытались решить этот вопрос, соединив различные нити в новых абзацах раздела «Обсуждение», абзац шесть. Мы надеемся, что они значительно улучшат статью и прояснят in vivo актуальность модели транскрипции с несколькими состояниями, которую мы предлагаем.Мы также включаем схематическую диаграмму вместе с новыми данными из имитационной модели, чтобы помочь отреагировать на этот момент (рисунок 9).
[Примечание редакции: до принятия были запрошены дополнительные исправления, как описано ниже.]
В этой редакции Фезерстоуна и др. Авторы внесли в рукопись некоторые текстовые и организационные изменения, чтобы лучше передать биологический смысл. Тем не менее, я все еще считаю, что рукописи не хватает привлекательности для широкой аудитории или даже для более узких читателей по экспрессии генов.На мой взгляд, этого недостаточно для того, чтобы по-новому взглянуть на развивающийся гипофиз. Хотя меня сильно привлекает подход, разработанный авторами, он, кажется, является продолжением их предыдущей работы в этой области, опубликованной в серии статей, в которых наблюдаются пульсации, изменения в процессе разработки и рефрактерный период (Harper, JCS, 2010; Featherstone, JCS, 2011; Harper, PLoS Bio, 2011). Здесь они уточнили свою модель основной динамики, используя метод статистического вывода.Они приходят к выводу, что динамика транскрипции не подчиняется простому телеграфному процессу, но не выходит за рамки этой феноменологии и не анализирует функциональные последствия.
Настоящим достижением в этой статье является наблюдаемая пространственная связь между отдельными клетками и открытие, что эта пространственная связь изменяется в процессе развития. Это связывание, которое происходит в масштабе ~ 30 мкМ, нарушается ограниченным расщеплением трипсином. Это открытие действительно интересно, но поднимается ли он до уровня самодостаточной истории? И нельзя ли представить это более убедительно? В общем, считаю, что работа технически завершена, но мое общее впечатление таково, что они не описали четко значительный прогресс в нашем биологическом понимании.
Один рецензент усомнился в значимости нашего исследования для широкой биологической аудитории. Мы считаем, что эта статья не только представляет важные достижения как в теоретическом, так и в экспериментальном подходе, но также является первым исследованием с достаточным разрешением и пропускной способностью для оценки влияния тканевой среды на пространственную координацию экспрессии генов. В целом, статья представляет собой полный интегрированный и количественный анализ контроля динамики транскрипции, как временной, так и пространственной, в процессе развития в ключевой ткани млекопитающих.Никакие предыдущие исследования не достигли этого.
Основные изменения, которые мы внесли, следующие:
Мы переписали аннотацию, чтобы сделать основные идеи статьи более понятными.
Мы упростили и оптимизировали Введение, чтобы более четко представить основные справочные вопросы. В ответ на незначительные комментарии рефери мы также удалили субъективные комментарии о значимости или важности.
Мы не изменили цифры, кроме незначительных корректировок маркировки, но мы попытались уточнить описание результатов, показанных на рисунках, чтобы сделать ключевые сообщения более доступными для широкого круга читателей.Изменения в разделе «Результаты» особенно касаются обработки результатов, показанных на рисунках 2, 4 и 7, где мы попытались уточнить представление в основном тексте. Мы также немного изменили условные обозначения фигур (особенно рисунок 7) и скорректировали маркировку рисунков (рисунок 2, рисунок 5, рисунок 7, рисунок 8) для большей ясности.
Изменения в обсуждении были внесены в последние два абзаца, чтобы подчеркнуть ключевые новые результаты, которые мы попытались резюмировать более кратко.Вот эти выводы:
1) Моделирование динамики транскрипции с несколькими состояниями показывает, что импульсы транскрипции не являются просто бинарными по своей природе, и мы характеризуем поведение переключения в интактной ткани.
2) И длительность импульсов, и координация динамики транскрипции различаются между незрелой и взрослой тканями гипофиза.
3) Структура ткани влияет на клеточное поведение и обеспечивает локальную координацию динамики транскрипции. Наша работа не только с трипсином, но и с блокаторами щелевых соединений указывает на то, что в этом поведении участвует передача сигналов щелевых соединений.
https://doi.org/10.7554/eLife.08494.026Семинар BMSE: «Неравновесная регуляция доступности хроматина и транскрипции в процессе развития» | Биомолекулярная наука и инженерия
Спикер
Проф. Эрнан Гарсия (Калифорнийский университет в Беркли)Дата и место
Среда, 16 октября 2019 г., 11:001601 Elings Hall
Аннотация
Открытой проблемой в биологии развития является предсказание паттернов экспрессии генов на основе знания пространственно-временной динамики концентрации входных факторов транскрипции и их связывания с регуляторной ДНК.После успехов у бактерий большинство описаний регуляции транскрипции в развитии было построено в предположении равновесия, когда энергия не расходуется во время связывания фактора транскрипции или взаимодействия с транскрипционным аппаратом. Однако появляется все больше свидетельств того, что такие процессы, как расход АТФ на модификацию хроматина, очень короткое время связывания факторов транскрипции с ДНК и динамический характер общего транскрипционного аппарата, способствуют нарушению равновесия системы.Чтобы подвергнуть это широко распространенное предположение о равновесии строгой проверке, мы используем широко изученное формирование ступенчатого паттерна экспрессии гена горбуна, опосредованного активатором Bicoid и пионерским фактором транскрипции Zelda в раннем эмбрионе плодовой мушки Drosophila. Используя комбинацию количественной визуализации отдельных клеток вживую, теоретических моделей и компьютерного моделирования, мы демонстрируем, что никакие процессы равновесия не могут повторить, как Bicoid и Zelda диктуют выражение горбунов.Вместо этого мы показываем, что регуляция горбачей может быть описана только моделями, в которых энергия расходуется на регуляцию транскрипции, что, как мы предполагаем, происходит из процесса обеспечения доступности хроматина для факторов транскрипции. Наши результаты предполагают, что для достижения предсказательного и количественного понимания принятия клеточных решений в процессе развития необходимо идентифицировать и охарактеризовать шаги, которые работают вне равновесия в транскрипционном каскаде, где расходуется энергия.
границ | Выживание долгоживущих плазматических клеток (LLPC): собираем головоломку
Введение
Плазматические клетки (ПК) представляют собой важное звено гуморального иммунитета как главную линию защиты от инфекции и повторного инфицирования. Как основные продуценты циркулирующего иммуноглобулина (Ig), эти клетки обеспечивают жизненно важный прочный и защитный иммунитет против множества патогенов. Продольный анализ титров антиген-специфических антител от вакцинированных людей показывает, что прогнозируемый период полувыведения титра кори составляет 3014 лет (1).Это свидетельство долговечной защиты, которую может обеспечить ПК. В отличие от других подмножеств иммунных клеток, таких как Т-клетки или В-клетки, сложность различных подмножеств ПК только начинает пониматься. Генерация плазматических клеток происходит в основном при Т-клеточной дифференцировке В-лимфоцитов в ПК в реакциях зародышевого центра (2, 3). Текущая парадигма предполагает, что в результате этих взаимодействий развиваются два основных типа ПК: короткоживущие плазматические клетки (SLPC) и долгоживущие плазматические клетки (LLPC) (4, 5).Подмножество LLPC, которое, как типично считается, перемещается в костный мозг (BM) и находится в нем, является подмножеством, которое обеспечивает долгосрочную и устойчивую продукцию антител, которая сохраняется в течение десятилетий до всей жизни человека (6-8). Реакции зародышевого центра посредством соматической гипермутации и рекомбинации с переключением классов позволяют отобрать ПК, продуцирующие высокоаффинные антитела, которые, как предполагается, являются основными предшественниками LLPC (9, 10). Однако существует относительно мало понимания движущей силы, по которой эти LLPC могут стать долгоживущими.
Специализированные ниши для LLPC
Были проведены обширные исследования биологии PC в целом, от того, как они генерируются и задействованы ключевые программы транскрипции, до их способности перемещаться в различные органы (11-15). Дальнейшие исследования выявили клеточные и молекулярные компоненты различных органоспецифических ниш, занятых PC (16-18). Однако существует относительно мало понимания того, что отличает способность LLPC выживать от SLPC.LLPC не являются долгоживущими по своей природе, поскольку их выживание критически зависит от способности получать доступ и использовать фиксированное количество специфических ниш, способствующих выживанию, в костном мозге, вторичных лимфоидных органах, слизистой оболочке и участках воспаления (5, 7, 19). –24). BM традиционно считается основным органом резиденции LLPC. Он обеспечивает динамическую инфраструктуру, поддающуюся формированию сложной микросреды, и позволяет создавать специфические ниши клеточного типа легче, чем другие менее пластичные органы (5, 6).
Более поздняя работа пролила свет на тот факт, что LLPC не только находятся в пределах BM. Около 80% ПК человека расположены в лимфоидной ткани, ассоциированной с кишечником (GALT), и продуцируют в основном IgA (25). Это обеспечивает толерантность к комменсальным бактериям в нашем кишечнике, а также обеспечивает защиту от неблагоприятных микробных и пищевых антигенов. Первоначально считалось, что постоянная активация В-клеток в слизистой оболочке снабжает пул IgA-продуцирующего ПК в кишечнике (26, 27). Однако новые исследования показали, что ПК в кишечнике также может сохраняться в течение длительных периодов времени.Антитела, специфичные к Escherichia coli , были обнаружены через 112 дней после воздействия (28). Исследование биоптатов кишечника, хранящихся в культуре, содержало непролиферирующий IgA-продуцирующий ПК в течение> 4 недель (29). Другое исследование показало, что через 9 месяцев после иммунизации как IgA-индуцирующим антигеном (холерный токсин), так и Т-зависимым антигеном (Ova) антиген-специфический ПК может быть обнаружен в собственной пластинке (LP), но также и в BM ( 21). Это предполагает, что ниши выживания, присутствующие в кишечнике, могут быть подобны нишам в BM и что PC слизистой оболочки могут использовать эти ниши таким же образом, как PC BM, а также вносить вклад в пул LLPC BM (21).
Традиционно считается, что большинство BM LLPC секретируют антитела IgG, а те, которые находятся в участках слизистой оболочки, таких как кишечник, продуцируют антитела IgA, однако около 40% BM PC также продуцируют IgA (30). Также сообщалось, что в BM существуют долгоживущие низкоаффинные IgM, продуцирующие ПК, которые, по-видимому, занимают разные ниши (20). Кроме того, аллергическая сенсибилизация овальбумином генерирует IgG, IgA и IgE, секретирующие ПК, которые могут обнаруживаться в костном мозге в течение длительного периода (31).Следовательно, данные свидетельствуют о том, что долговечность ПК в первую очередь определяется не изотипом его антител, а, скорее, природой патогена, характеристиками начальной активации В-клеток, вызванной патогеном, и нишей, в которой находится результирующий ПК. Эта широкая структура предполагает, что основная роль SLPC заключается в защите от частых (и менее тяжелых) эндемических инфекций, которые поддерживаются повторяющейся и антиген-зависимой реактивацией B-клеток; тогда как LLPC обеспечивает устойчивый защитный иммунитет против редких (но более тяжелых) эпидемических инфекций антиген-независимым образом.Также было высказано предположение, что ранние сигналы активации B-клеток / плазмобластов определяют, входят ли PC и отвечают на сигналы выживания в нише PC (32-34). Поскольку пространство, доступное LLPC, ограничено, долговечность LLPC требует как доступа к нише выживания, так и способности реагировать на уникальные сигналы ниши в пользу выживания.
LLPC и B-клетки памяти — не одно и то же
Первоначально предполагалось, что устойчивые ответы антител являются результатом постоянного пополнения пула SLPC путем непрерывной повторной стимуляции В-клеток памяти (35).Тем не менее, наблюдения, что трансплантация костного мозга вызвала аллергию у людей, не страдающих аллергией, посредством передачи аллерген-специфической продукции IgE, подтвердили идею о том, что эти ПК были долгоживущими из-за отсутствия антигенной повторной стимуляции у этих пациентов (36, 37). Дальнейшие исследования показали in vivo , что некоторые субпопуляции ПК действительно являются долгоживущими и что отсутствие антигена плюс истощение В-клеток памяти из-за облучения не отменяет способность этой субпопуляции LLPC продолжать продуцировать высокоаффинные антиген-специфические антитела. (38, 39).Другие исследования показали, что длительное терапевтическое истощение общего пула В-клеток не повлияло на количество антиген-специфических PC PC или титры антител в моделях вакцинированных мышей (40), а также не повлияло на титры антител против детских вакцин у людей (41). В исследованиях на людях титры вакцин-специфических антител поддерживались после лечения моноклональными антителами против CD20 (которые нацелены на В-клетки, но не на ПК), несмотря на явное истощение пула В-клеток памяти (41). Устойчивая аплазия B-клеток, вызванная CD19 CAR-T-клетками (которые также нацелены на B-клетки, но не PC) у взрослых и детей, у пациентов с острым лимфоцитарным лейкозом также не влияла на титры антиген-специфических антител сыворотки вакцины или количество PC в BM или подвздошной кишке. и толстая кишка (42).Исследование динамики ПК из биопсии пересаженной двенадцатиперстной кишки показало, что через 1 год после трансплантации CD38 + ПК от донора все еще можно идентифицировать. Дальнейшая характеристика идентифицировала CD19 — и CD19 + PC, присутствующие в этих биопсиях, где подмножество CD19 — PC представляло стабильную популяцию с потенциально долгой продолжительностью жизни (24). Наконец, характеристика подмножеств PC человека продемонстрировала, что CD19 – PC в BM были преимущественно секретирующими IgG со зрелым фенотипом и оставались оседающими в BM после системной вакцинации (43).Это демонстрирует, что LLPC действительно отличается от других подмножеств B-клеток, однако ключевые факторы, которые отличают эти клетки, только недавно начали быть охарактеризованы.
Уникальный транскрипционный профиль
Во время дифференцировки В-клеток в ПК подавляется регуляция многих генов, включая те, которые участвуют в презентации антигена и функции BCR. Одновременно происходит активация линии PC, определяющей фактор транскрипции PRDM1 (BLIMP-1) и генов, участвующих в трансляции белка и развернутом ответе белка ( XBP-1 ) (11, 44).Способ отличить это подмножество LLPC лежит через их транскрипционный профиль. Недавнее исследование ПК человека установило уникальный профиль, который различает ранний ПК, ПК в циркулирующей крови и долгоживущий BM-резидентный CD138 + ПК (14). Сигнатура, которая отличает LLPC, содержит значительное количество генов с пониженной регуляцией и только несколько генов с повышенной регуляцией. Эта последняя группа включает гены антиапоптоза Mcl-1 и ZNF667 , гены, связанные со стрессом ER, включая EROILB и MANF , транспорт катионов ATPase ATP12A и TFBS и играют роль в удержании гематопоэтических клеток-предшественников в BM.Многие из этих генов связаны с центральной функцией PC по выработке значительных количеств белка в форме иммуноглобулина — хотя тот факт, что эта функция не является уникальной для LLPC, предполагает, что может быть дополнительная недооцененная биология этих генов в LLPC.
Идентификация профиля транскрипции, уникального для LLPC, сталкивается со значительным экспериментальным препятствием, а именно с возможностью идентифицировать чистую популяцию LLPC для анализа. Анатомическое расположение традиционно было способом, которым SLPC и LLPC отличались друг от друга из-за отсутствия отдельных фенотипических маркеров, различающих эти подмножества.Однако, как отмечалось выше, недавнее сложное фенотипическое профилирование сделало все более очевидным, что LLPC не единственные ПК, присутствующие в BM. Недавняя работа, посвященная популяциям предшественников PC, определила, что несколько волн трафика могут происходить после воздействия антигена и дифференцировки PC, в результате чего другие PC, помимо LLPC, находятся в BM (34). Кроме того, недавно было показано, что поверхностные маркеры, которые традиционно использовались для идентификации LLPC, по-разному окрашивают различные подмножества ПК — например, анализ ПК в компартменте BM показал, что существует различная экспрессия поверхностных маркеров, таких как CD38 и CD19 (43, 45).Эти фенотипические различия четко связаны с функциональными различиями, такими как то, продуцируют ли эти ПК антитела с высоким или низким сродством (46).
Из-за недавнего увеличения гетерогенности субпопуляции PC, анализ общей популяции PC PC по сравнению с PC из других компартментов может не выявить специфические факторы и взаимодействия, ответственные за усиление и поддержание сигнатуры транскрипции, необходимой для субпопуляции LLPC. Таким образом, один из подходов к определению ключевых транскрипционных сетей, уникальных для LLPC, состоит в том, чтобы сначала идентифицировать вышестоящие рецепторы и пути передачи сигналов, которые однозначно поддерживают выживание и продолжительность жизни LLPC.
Внутренние сигналы для выживания и функционирования LLPC
BCMA
Теперь мы знаем, что существуют внутренние сигналы, уникально используемые LLPC, которые регулируют их долгосрочное выживание и функцию (Таблица 1). Хорошо известно, что такие факторы, как лиганд, индуцирующий пролиферацию (APRIL), интерлейкин-6 (IL-6), фактор активации В-клеток (BAFF), CD44 и CXCL12, могут поддерживать выживаемость как SLPC, так и LLPC в различных органах (4 , 12, 29, 47–49). Одним из рецепторов как BAFF, так и APRIL является антиген созревания В-клеток (BCMA), который в высокой степени регулируется на PC (50).Генетический нокаут Tnfrsf17 (который кодирует BCMA) привел к значительной потере секретирующих антитела клеток (ASC) в BM по сравнению с контролем дикого типа (Wt) через 6-8 недель после иммунизации. Однако ответы зародышевого центра и ранние антиген-специфические сывороточные титры IgM и IgG были нормальными, что указывает на то, что потеря BCMA в первую очередь повлияла на подмножество LLPC (50). Дополнительные исследования показали, что выживаемость В-клеток памяти мышей не зависит от ВСМА, а ВСМА индуцируется только в В-клетках человека, преданных дифференцировке ПК (47, 51).Это говорит о том, что зависимость от BCMA для выживания специфична для компартмента ПК. Кроме того, исследования множественной миеломы (ММ), резидентной опухоли LLPC с переключаемым классом Ig, показали, что BCMA значительно способствует выживаемости и пролиферации ММ (52). Терапевтическое воздействие на BCMA в MM показало высокую селективность в отношении популяций MM и PC (53), что подтверждает роль рецептора, ограниченную PC. Активация BCMA с помощью BAFF увеличивает экспрессию антиапоптотической молекулы Mcl-1, указывая на механистическую основу, с помощью которой ось BAFF / BCMA способствует долгосрочному выживанию (50).Увеличение экспрессии гена Mcl-1 в BM PC наблюдалось у мышей Wt по сравнению с BM PC у мышей BCMA — / — , где как мРНК Mcl-1, так и экспрессия белка были значительно ниже в нокаутах (54). Кроме того, потеря Mcl-1 вызывает значительное снижение антиген-специфических титров через 21 день после иммунизации, что согласуется с влиянием на подмножество LLPC (54). Как и BCMA, Mcl-1 также является критическим фактором для выживаемости MM (55, 56), что согласуется с более широкой концепцией, согласно которой большая часть биологии миеломы на самом деле является биологией нормального LLPC (57).
Таблица 1 . Резюме внутренних факторов, поддерживающих выживаемость LLPC.
STAT3
Значение IL-6 для функции ПК и выживаемости широко изучено (49, 58). IL-10 и IL-21 также участвуют в созревании и выживании PC человека (59–61). IL-21 преимущественно продуцируется Т-фолликулярными клетками-помощниками (Tfh) (62), которые участвуют во многих фазах гуморального иммунного ответа от активации В-клеток до созревания и дифференцировки В-клеток памяти в ПК (63).Исследование миндалин, селезенки, лимфатических узлов и ПК КМ показало, что ПК КМ имеет очень низкую экспрессию IL-21R и что IL-21 только усиливает секрецию Ig из вторичного лимфоидного ПК, но не ПК КМ (64). Однако более поздняя работа постулировала, что клетки Tfh влияют на индуцированный Ag в крови PC, который может содержать предшественники подмножества LLPC (65). Неизвестно, происходит ли этот эффект непосредственно через IL-21 или влияние IL-21 на субпопуляции B-клеток и активацию рецепторов, таких как IL-6R. Как эти отдельные цитокины способствуют выживанию и функции BM PC, пока неясно; однако все они инициируют передачу сигналов STAT3 (66–68).Активация STAT3 играет важную роль в способности ПК отвечать на эти различные цитокиновые сигналы (68). Нарушение этого пути ингибирует цитокин-зависимое повышение жизнеспособности и секреции IgG. Более того, активация STAT3, по-видимому, является предпосылкой для способности ПК отвечать на APRIL и BAFF. Это указывает на критическую роль STAT3 как ключевого нижестоящего сигнального узла, передающего стимулы в LLPC, что усиливает их выживание и продолжительность жизни (68).
Айолос
Другие факторы, ограниченные лимфоидным происхождением, были изучены на предмет их возможной роли в поддержке подмножества LLPC.Aiolos, часть семейства генов Ikaros ядерных регуляторов, которые действуют в модуляции структуры хроматина, является одним из этих факторов (69). Отсутствие этого гена не влияет на развитие В-клеток или ранний ответ SLPC, но влияет на долгосрочную выживаемость ПК (70). Иммунизация обычно вызывает первую волну низкоаффинных антител, продуцируемых главным образом SLPC, а затем вторую волну более высокоаффинных антител из BM LLPC (10). После делеции Aiolos наблюдается значительное снижение продукции высокоаффинных антител в BM через 120 дней после первичной иммунизации без влияния на ответ низкоаффинных антител (71).Дополнительные исследования показывают, что делеция этого гена не вызывает дефекта самонаведения. Это предполагает, что активация Aiolos необходима для высокоаффинного антиген-специфичного LLPC, в отличие от других факторов, таких как BLIMP-1, которые необходимы для генерации всех PC. Точный механизм еще предстоит понять, но способность Aiolos стимулировать высокоаффинные антитела важна для долгосрочного усиления защитного иммунитета.
CD93
Другой отправной точкой для идентификации специфических рецепторов, участвующих в выживании и долголетии LLPC, являются те рецепторы, которые в большей степени регулируются на BM PC по сравнению со зрелыми B-клетками.Гринли и др. наблюдали, что CD93, который в значительной степени охарактеризован своей ролью в межклеточной адгезии и клиренсе апоптотических клеток, не присутствует на зрелых В-клетках, но сильно активируется во время дифференцировки ПК (72). После иммунизации Т-независимыми (TI) или Т-зависимыми (TD) антигенами CD93 дополнительно активируется в BM PC, а CD93 + CD138 + PC имеют повышенную экспрессию BLIMP-1, XBP-1, IRF4 и продуцируют антитела с более высоким сродством, что позволяет предположить, что этот рецептор идентифицирует субпопуляцию LLPC (73).В соответствии с этим потеря CD93 уменьшала устойчивость долгосрочных титров антител после иммунизации (73). Потеря Blimp-1 снижает экспрессию CD93 только в PC, поскольку CD93 все еще присутствует на низких уровнях в незрелых B-клетках с дефицитом Blimp-1. Эта связь с Blimp-1, который наиболее сильно экспрессируется в LLPC, привела авторов к гипотезе о том, что CD93 необходим для стимулирования зрелого фенотипа LLPC и продукции высокоаффинных антител. Это дополнительно подтверждается недавним исследованием титров кори у вакцинированных школьников, в котором наиболее высокоэкспрессированными генами в группах с высоким ответом были CD93, IL-6 и CXCL12, что указывает на участие CD93 в генерации долгоживущих антител. человек (74).
CD28
После активации В-клетки подавляют фактор транскрипции Рах5, определяющий клон В-клеток (75). Было обнаружено, что Pax5 непосредственно ингибирует экспрессию гена CD28 в B-клетках, а подавление Pax5 приводит к усилению регуляции CD28 как немедленному / раннему событию в дифференцировке B → PC. В этом исследовании потеря CD28 нарушала кратковременную индукцию титров антиген-специфических антител, хотя долгоживущие ответы антител не оценивались. Предыдущие исследования у мышей с глобальным нокаутом CD28 и с нокаутом рецептора CD28 продемонстрировали значительные дефекты в ответах антител (76–79), но это было приписано дефектам в помощи Т-клеток.Последующая работа в нашей лаборатории показала, что CD28 играет важную роль, присущую ПК, в выживании LLPC и поддержании устойчивых титров антиген-специфических антител IgG после вакцинации (80). По сравнению с мышами Wt, невакцинированные мыши CD28 — / — имеют сопоставимое количество ПК в селезенке, но имеют значительно меньшее количество ПК. Исследования in vitro продемонстрировали, что активация CD28 сама по себе (без «сигнала 1», который необходим для Т-клеток) увеличивает выживаемость очищенного PC BM, но не PC селезенки, в ответ на смерть, вызванную сывороточным голоданием. In vivo , потеря экспрессии CD28 специфически в линии B-клеток (B-клетки не экспрессируют CD28, только PC) привела к неспособности поддерживать титры антиген-специфических антител в этих мышиных моделях (80). Работа, проделанная Njau et al. также посмотрел влияние CD28 на ПК. Первоначальные исследования с глобальными мышами CD28 — / — показали значительное увеличение антиген-специфических ответов IgM по сравнению с мышами Wt, но обнаружили значительно более низкий антиген-специфический ответ IgG по сравнению с контрольными мышами Wt через 60 дней после иммунизации, что они приписали дефект в помощи Т-лимфоцитов (81).Однако, в отличие от Rozanski et al. специфическая потеря CD28 в линии B-клеток привела к значительному увеличению титров антиген-специфических антител по сравнению с Wt PC. Основа различий между этими двумя исследованиями неясна, хотя стратегии иммунизации (как антиген, так и, в частности, используемые адъюванты) были разными. Независимо от того, в обоих исследованиях установлено, что CD28 присутствует на ПК и играет роль в влиянии на ответы антител.
Исследования сигнальных путей, исходящих от рецептора CD28 в LLPC, продемонстрировали, что потеря нижестоящей передачи сигналов PLCγ1, опосредованной Vav / Grb2, вызвала потерю BM LLPC, в то время как потеря нижестоящей передачи сигналов PI3K / AKT не имела никакого эффекта (82).Интересно, что хотя мышиный SLPC также экспрессируют CD28, прямая активация рецептора не вызывает последующую активацию PLCγ1 и NFκB, что указывает на сигнальную основу, почему SLPC не реагирует на CD28, и предполагает, что CD28 имеет разные пороги активации в SLPC по сравнению с LLPC. Далее активация CD28 увеличивает экспрессию BLIMP-1, главного регулятора идентичности ПК и продукции антител (82, 83). В соответствии с ролью, которую CD28 играет в нормальном LLPC, мы также обнаружили, что активация CD28 дает основной сигнал, способствующий выживанию / устойчивости к химиотерапии при ММ (84, 85).
Активация CD28 на ММ непосредственно трансдуцирует сигнал, способствующий выживанию, а также сигналы обратно дендритным клеткам (ДК) через лиганды CD28, CD80 и CD86, экспрессируемые на этих ДК (84–86). Эта обратная передача сигнала через CD80 / CD86 активирует экспрессию / активность как IL-6, так и иммуносупрессивного фермента индоламин-2,3-диоксигеназы (IDO) в DC, что способствует дальнейшему выживанию MM. Таким образом, CD28 как напрямую доставляет сигнал, способствующий выживанию, миеломной клетке, так и модулирует DC в нише для дальнейшей поддержки выживания MM.Напротив, блокада in vivo взаимодействия CD28-CD80 / CD86 с CTLA4-Ig (который одобрен FDA для лечения ревматоидного артрита) значительно повысила чувствительность к химиотерапии на мышиной модели ММ (84). Учитывая значительную общую биологию между LLPC и MM, эта двунаправленная сигнальная ось между CD28 и CD80 / CD86 в настоящее время исследуется как ранее неизвестные факторы выживания для LLPC.
Аутофагия
Поскольку LLPC постоянно генерируют и секретируют высокий уровень белка Ig, им требуются уникальные метаболические пути, чтобы справиться с белковой нагрузкой и стрессом ER.Индукция развернутого белкового ответа (UPR) была четко определена как важная для дифференцировки В-клеток в ПК (87). Второй механизм, связанный с адаптацией к этому стрессу, — это аутофагия. Основная функция аутофагии — поддерживать клеточный метаболизм при голодании. У дрожжей, чтобы уравновесить стресс, вызванный экспансией ER, аутофагическая обрезка используется для удаления избытка ER (88). Внутри иммунного компартмента зрелые Т-клетки полагаются на опосредованный аутофагией оборот митохондрий (митофагия), чтобы контролировать внутриклеточное производство активных форм кислорода (89, 90).Аутофагия также имеет решающее значение для выживания LLPC. Поскольку продуцирующие антитела ПК обладают высокой степенью биосинтеза, можно предположить, что аутофагия играет ключевую роль как в обработке / устранении / рециркуляции неправильно свернутых иммуноглобулинов, так и в обеспечении потребностей ПК в топливе и метаболическом субстрате — в частности, поскольку микроокружение костного мозга может столкнуться с серьезными проблемами. переменная доступность питательных веществ из-за изменения уровней кроветворения во время стресса (91, 92). Анализ секретома мезенхимальных стромальных клеток (которые, как известно, являются частью ниши BM) идентифицировал два основных фактора выживания ПК BM, фибронектин (FN-1) и белок YWHAZ, которые, как было показано, подавляют передачу сигналов mTORC1 в ПК BM (17 ).Индукция аутофагии обычно происходит после ингибирования mTOR (мишень рапамицина), эволюционно консервативной протеинкиназы. mTOR является ключевым регулятором роста клеток и реакции на состояние питательных веществ и стрессовые сигналы в клетке, и известно, что он негативно регулирует аутофагию (93). Т.о., эти факторы в нише могут в дальнейшем влиять на способность LLPC активировать аутофагию как механизм выживания. Другие исследования показали, что у мышей, нокаутированных по Atg5 , который играет центральную роль в инициации аутофагии, имеется значительный дефект в постиммунизации высокоаффинного LLPC антигеном TD (94).Хотя все ПК вырабатывают Ig на высоких уровнях, возможно, что LLPC в большей степени зависят от аутофагии в течение своей жизни как для устранения неправильно свернутого белка, так и для механизма рециркуляции для обеспечения метаболических субстратов, чтобы усилить метаболическую пригодность этих клеток во время периодов. недостатка питательных веществ.
Метаболизм
Недавние исследования начали связывать пути производства метаболической энергии, используемые LLPC, с путями, используемыми Т-клетками памяти (которые должны быстро реагировать на активацию, чтобы стать высоко биосинтетическими и размножаться).Костимуляция CD28 в наивных Т-клетках переключает их использование топлива на жирные кислоты и митохондриальное окисление, чтобы поддерживать их метаболические потребности, и было показано, что это необходимо для генерации Т-клеток памяти (95, 96). Костимуляция CD28 также вызывает изменения в митохондриях Т-клеток памяти — удлиненные митохондрии с увеличенной структурой крист связаны с повышенной (резервной) респираторной способностью по сравнению с наивными Т-клетками. Точно так же работа Lam et al. О метаболическом фенотипе LLPC.интересно показало, что LLPC мыши и человека обладают большей максимальной респираторной способностью, чем SLPC, и, хотя они обладают повышенной способностью поглощать глюкозу (через активацию Glut1), они постоянно используют длинноцепочечные жирные кислоты в качестве основного источника топлива — и появляется глюкоза. в первую очередь использоваться для гликозилирования антител (97, 98). Однако в возможно неоптимальных условиях они также способны использовать гликолиз и импорт митохондриального пирувата. Важность этой «альтернативной» программы метаболизма была продемонстрирована обнаружением того факта, что потеря способности импортировать пируват в митохондрии значительно снижает частоту LLPC и отменяет устойчивый долгоживущий ответ антител через 22 недели после вакцинации (97).Аналогичная работа, посвященная IgA-секретирующему ПК в LP, показала более высокую экспрессию связанных с гликолизом метаболитов, чем наивные В-клетки в пейеровских бляшках (PP) (99). Предполагается, что переключение с IgM на IgA требует метаболического сдвига и предпочтительного использования гликолитического пути. Эта группа также отметила, что SLPC селезенки не содержал очень много гликолитических метаболитов, что указывает на то, что и BM, и кишечник являются уникальными микроокружениями, которые могут способствовать этому сдвигу метаболического фенотипа LLPC.
ENPP1
Дальнейшее исследование потенциально уникальных метаболических характеристик LLPC привело к открытию других ранее неизвестных путей. Трансмембранный гликопротеин эктонуклеотидпирофосфатаза / фосфодиэстераза 1 (ENPP1) высоко экспрессируется на ПК, но ее функция была продемонстрирована только недавно. Этот фермент ранее был охарактеризован в связи с его ролью в формировании костей, гомеостазе глюкозы и подавлении сигнального пути инсулина (100).У мышей ENPP1 — / — также обнаружено значительное снижение BM LLPC через 12 недель после иммунизации TD-зависимым антигеном и инфекции C. chabaudi , но не наблюдалось никакого эффекта на ПК в селезенке (101). ENPP1 — / — PC потребляют меньше глюкозы, чем их аналоги Wt, и обнаруживают нарушение гликолиза, что указывает на роль ENPP1 как регулятора глюкозы и метаболического пути, обнаруженного в LLPC (101).
Параллели между мышами и людьми
Актуальность некоторых из вышеперечисленных факторов для поддержки выживания LLPC недавно была подтверждена характеристиками BM PC человека.Эти исследования элегантно продемонстрировали, что гетерогенность BM PC у мышей очевидна и у людей, из которых только часть, по-видимому, является истинной долгоживущей субпопуляцией. В этом исследовании определены 4 подгруппы ПК, обнаруженные в BM: CD19 + CD38 hi CD138 — (подмножество A), CD19 + CD38 hi CD138 + (подмножество B), CD19 — CD38 hi CD138 — (подмножество C) и CD19 — CD38 hi CD138 + (подмножество D) (45).Подгруппа D не пролиферировала по сравнению с тремя другими подгруппами и была CD20-отрицательной, HLA-DR-отрицательной, но положительной по CD28 (45). В этой подгруппе было наибольшее количество IgG-специфических антител против столбняка, кори, эпидемического паротита и гриппа. Интересно, что эта подгруппа также обнаружила отличительную сигнатуру транскриптома РНК для усиления аутофагии. Это открытие предполагает, что те компоненты, которые были обнаружены в исследованиях на мышах, имеют отношение к ПК человека и имеют фундаментальное значение для функции и выживания LLPC.
Внешние сигналы и вклад ниши
General PC Partners: стромальные клетки, мегакариоциты и базофилы
Внутренняя сигнализация ПК и нисходящие ответы, описанные выше, не возникают в вакууме.LLPC критически зависят от сигналов, которые они получают из ниши, поскольку LLPC, культивируемый отдельно in vitro, быстро умирает (102). Исследования ниши LLPC BM идентифицировали несколько различных типов клеток, которые могут передавать сигналы о выживании PC (Figure 1). Первым идентифицированным типом клеток были ретикулярные и мезенхимальные стромальные клетки внутри костного мозга. Они обладают высокой экспрессией CXCL12, лиганда CXCR4, экспрессируемого на ПК, который отвечает за их доставку в BM (18, 103).Однако, хотя первоначальные исследования продемонстрировали, что эти стромальные клетки поддерживали выживаемость PC in vitro , эта выживаемость не была устойчивой — и in vivo , они, по-видимому, не играли существенной роли в поддержке выживаемости LLPC (48, 104). Более того, факторы выживания PC, такие как APRIL, BAFF и IL-6, не секретируются этими клетками. Тем не менее, недавно было показано, что мезенхимальные стромальные клетки BM секретируют фибронектин (FN-1) и другие белки (YWHAZ), которые могут поддерживать выживание LLPC in vitro в сочетании с гипоксией и такими факторами, как APRIL (17).FN-1 связывает интегрины, коллаген и CD138 (высоко экспрессируется на LLPC) и, как предполагается, играет важную роль в связывании ПК с внеклеточным матриксом костного мозга. Исследования ММ также показали, что опосредованное интегрином связывание с фибронектином способствует устойчивости к опосредованному химиотерапией киллингу (105).
Рисунок 1 . Внешние факторы в нише LLPC. PC связываются с DC через взаимодействия CD28: CD80 / CD86 и с Treg. Стромальные клетки, экспрессирующие CXCL12, действуют как сигнал самонаведения для CXCR4 + PC.Также присутствуют мегакариоциты и эозинофилы, которые, как было показано, продуцируют растворимые факторы, такие как APRIL и BAFF, для повышения выживаемости ПК.
Другие исследования идентифицировали мегакариоциты как часть ниши LLPC. Мегакариоциты могут продуцировать APRIL и IL-6, а мыши, не обладающие способностью образовывать мегакариоциты, имеют меньшее количество BM PC по сравнению с мышами Wt (106). Кроме того, после иммунизации овальбумином (Ova) было обнаружено, что Ova-специфические PC находятся в прямом контакте с мегакариоцитами в BM.Однако функциональные исследования постиммунизации ПК в отсутствие мегакариоцитов не показали какого-либо эффекта на поздней стадии антиген-специфических ответов антител. Базофилы — это еще один тип клеток, поддерживающий выживаемость ПК. В in vitro культурах базофилов и PC BM присутствие базофилов поддерживало выживание этих PC и заметно увеличивало продукцию Ig этими клетками (107). Однако функциональные исследования влияния базофилов на антиген-специфические ответы BM PC были оценены только in vitro , поэтому до сих пор не охарактеризовано, как эти клетки могут влиять на LLPC BM в условиях in vivo .Было определено, что эффекты выживания, наблюдаемые у базофилов, наиболее вероятно связаны с их способностью секретировать IL-4 и IL-6. Т.о., мегакариоциты и базофилы могут играть важную роль в установлении ниши LLPC и, возможно, доступа к LLPC, однако остается неясным, как они вносят вклад в долгосрочное выживание и поддержание LLPC.
Эозинофилы
Другим подмножеством клеток, которое, как считается, обеспечивает ПК жизненно важные цитокины, такие как IL-6 и APRIL, являются эозинофилы. Сообщалось, что эозинофилы совместно локализуются с ПК как в костном мозге, так и в лимфоидной ткани, ассоциированной с кишечником (GALT), и что мыши с дефицитом эозинофилов имеют значительно меньше ПК в стабильном состоянии и после воздействия антигена (108, 109).Для IgA-секретирующего ПК эозинофилы необходимы для стимулирования продукции IgA и поддержания иммунного гомеостаза в GALT (110). При исследовании ПК в собственной пластинке было обнаружено, что одна треть ПК расположена рядом с эозинофилами (21). Что касается ниши BM, было обнаружено, что недостаток эозинофилов заставляет LLPC в BM подвергаться апоптозу. После восстановления эозинофилов количество PC временно увеличивалось, что указывает на роль этих клеток в нахождении PC в BM, а также в поддержании популяции PC, когда они там были (108).После активации эозинофил может способствовать дифференцировке В-клеток в ПК, секретирующий IgM, который не зависит от индукции зародышевых центров и образования ПК с высоким сродством, секретирующего IgG (111). Однако более поздние исследования поставили под сомнение, являются ли эозинофилы необходимым компонентом для выживания и функции LLPC, по крайней мере, на мышиных моделях это не было окончательным (112). Следовательно, текущая литература предполагает, что эозинофилы могут играть критическую роль в генерации и транспортировке PC как для участков слизистой оболочки, так и для BM, но их вклад в выживание, зрелость и долгосрочную функцию LLPC остается неустановленным.
Дендритные клетки
Параллельные исследования в этой нише были сосредоточены на клеточных компонентах, которые экспрессируют лиганды, которые могут взаимодействовать с рецепторами на LLPC, которые поддерживают выживание и функцию. Наша работа продемонстрировала ключевую роль CD80 и CD86, природных лигандов для CD28. Отсутствие этих лигандов у мышей с глобальным нокаутом значительно снижает количество LLPC BM и устойчивые антиген-специфические титры (80). Клетки, которые имеют самую высокую экспрессию CD80 и CD86, представляют собой миелоидные (обычные) DC.Другие миелоидные клетки (моноциты, эозинофилы) также могут иметь значительную экспрессию. Наша лаборатория и другие продемонстрировали, что совместная локализация DC и ПК происходит внутри BM. Кроме того, это поддерживает выживание LLPC in vitro (контактным и CD28-зависимым образом) и in vivo (80, 113). Это взаимодействие имеет дополнительное значение, так как DC могут продуцировать значительные количества про-PC цитокина выживания IL-6, и что активация CD80 / CD86 является одним из путей, который индуцирует продукцию DC IL-6 (86).Как отмечалось выше, DC также значительно увеличивает выживаемость MM зависимым от CD28-CD80 / CD86 образом. Это указывает на наличие двунаправленной оси передачи сигналов в нише, чтобы поддерживать LLPC как прямо, так и косвенно посредством модуляции клеточных партнеров, чтобы производить важные факторы выживания.
T Регуляторные ячейки
Другим важным партнером, недавно идентифицированным в нише LLPC BM, являются T-регуляторные (Tregs) клетки, находящиеся в тесной физической ассоциации с CD11c + DC. Было показано, что большое количество Treg совмещается с PC в BM in vivo , и это необходимо для гомеостатической поддержки популяций BM PC (113).Удаление Tregs во время инфекции вызывало значительное снижение количества PC BM. Фенотип этих BM Tregs соответствовал активированной подгруппе с повышенными супрессивными способностями и наиболее точно соответствовал профилю, наблюдаемому в поврежденной мышце. Интересно отметить, что в отличие от Treg-клеток селезенки, которые имеют небольшую экспрессию CTLA-4, эти Treg-клетки BM экспрессируют CTLA-4, который является членом суперсемейства CD28, которое также связывает CD80 и CD86. В отсутствие экспрессии Treg CTLA-4 количество PC BM увеличивалось в 3 раза.Авторы предположили, что CTLA-4 + Tregs вносят вклад в поддержание иммунной ниши для LLPC, частично за счет буферизации этой ниши против системных воспалительных изменений, а также поддерживая гомеостаз LLPC за счет ограничения пула PC (113). Однако неясно, какую роль играет сам CTLA-4 в нише ПК. Возможно, что посредством выражения в Tregs он действует гомеостатическим образом, ограничивая количество ПК, способных передавать трафик в BM и находиться в нем. Однако он также может связываться с рецепторами CD80 / CD86 на нишевых DC, вызывая повышенную регуляцию цитокинов, таких как IL-6, для поддержки выживания ПК.Альтернативно, Treg CTLA-4 может конкурировать и ограничивать связывание LLPC CD28 с этими лигандами на DC, как часть гомеостатической регуляции популяции LLPC. Интересно предположить, что этот клеточный триумвират LLPC, DC и Tregs модулируется динамическими молекулярными взаимодействиями между CD28, CD80 / CD86 и CTLA-4, обеспечивая стабильную нишу, которая критически поддерживает выживание и непрерывную функцию LLPC.
Заключительные замечания
Этот обзор выявил некоторые компоненты, которые образуют сети, которые позволяют выживать и поддерживать LLPC.Долгосрочная генерация защитных антител этой подгруппой имеет решающее значение для устойчивого защитного иммунитета. Понимание этих ключевых молекулярных и клеточных компонентов, которые поддерживают выживание и долголетие LLPC, дает нам инструменты для использования новых стратегий вакцинации, а также терапевтических стратегий против заболеваний, возникающих из этой клетки, таких как MM и аутоиммунные синдромы.
Стало ясно, что существуют отдельные подмножества ПК, определяемые анатомическим расположением, фенотипом, функцией и продолжительностью жизни (Таблица 2).Имеющиеся данные свидетельствуют о том, что ПК, производящие нейтрализующий Ig с высоким сродством (который был бы особенно важен для быстро летальных патогенов, которые обычно вызывают эпидемии), являются ключевыми членами подмножества LLPC и образуются при реакциях зародышевого центра и затем передаются в BM; где они обладают уникальными характеристиками, которые позволяют им разблокировать и занять / использовать нишу LLPC. Теперь ясно, что это значительное упрощение, учитывая гетерогенность популяций BM PC и ниш, присутствующих в других органах, таких как кишечник.Тем не менее, все больше данных демонстрирует, что «традиционные» LLPC, которые производят пожизненные защитные титры антител, требуют, чтобы как внутренние, так и нишевые компоненты ПК были долгоживущими. Эти взаимодействия могут затем привести к нижестоящей внутренней передаче сигналов PC, которая поддерживает способность подмножества LLPC выживать в течение десятилетий в их микросреде; который в BM гипоксичен и переполнен быстро пролиферирующими клетками (особенно реактивными BM, отвечающими на инфекцию), конкурирующими за пространство и питательные вещества, и на участках слизистой оболочки, где есть постоянное воздействие как полезных, так и вредных микробов.Таким образом, ключевые аспекты, лежащие в основе долговечности LLPC, вероятно, усиливают механизмы, позволяющие эффективно адаптироваться к этим стрессам на протяжении всей жизни.
Таблица 2 . Важность внутренних факторов в подмножествах В-клеток и ПК.
Авторские взносы
SL — основной автор и составитель основной части обзора, существенный вклад в концепцию и дизайн. AU внес изменения в содержание рукописи. KL внесла свой вклад в концепцию и дизайн, внесла критические изменения и окончательное утверждение, а также предоставила финансирование.
Финансирование
Эта работа была частично поддержана грантами трех Национальных институтов здравоохранения (NIH): награды R01CA121044 и T32CA085183 от Национального института рака (NCI) и R01AI100157 от Национального института аллергии и инфекционных заболеваний (NIAID).
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Список литературы
2. Smith KG, Light A, OReilly LA, Ang SM, Strasser A, Tarlinton D. Экспрессия трансгена bcl-2 ингибирует апоптоз в зародышевом центре и выявляет различия в отборе B-клеток памяти и антителообразующих клеток костного мозга. J Exp Med . (2000) 191: 475–84. DOI: 10.1084 / jem.191.3.475
PubMed Аннотация | CrossRef Полный текст | Google Scholar
3. Фан Т.Г., Паус Д., Чан Т.Д., Тернер М.Л., Натт С.Л., Бринк Р. В-клетки зародышевого центра с высоким сродством активно отбираются в компартмент плазматических клеток. J Exp Med . (2006) 203: 2419–24. DOI: 10.1084 / jem.20061254
PubMed Аннотация | CrossRef Полный текст | Google Scholar
4. Мозер К., Токойода К., Радбрух А., МакЛеннан И., Манц Р.А. Стромальные ниши, дифференцировка и выживание плазматических клеток. Curr Opin Immunol . (2006) 18: 265–70. DOI: 10.1016 / j.coi.2006.03.004
PubMed Аннотация | CrossRef Полный текст | Google Scholar
5. Sze DM, Toellner KM, de Vinuesa CG, Taylor DR, Ian MacLennan CM.Внутреннее ограничение роста плазмобластов и внешние пределы выживаемости плазматических клеток. J Exp Med . (2000) 192: 813–22. DOI: 10.1084 / jem.192.6.813
PubMed Аннотация | CrossRef Полный текст | Google Scholar
7. Слифка М.К., Матлоубиан М., Ахмед Р. Костный мозг является основным местом долгосрочной выработки антител после острой вирусной инфекции. Дж Вирол . (1995) 69: 1895–902.
PubMed Аннотация | Google Scholar
10. Смит К.Г., Лайт А., Носсал Г.Дж., Тарлинтон Д.М.Степень созревания аффинности различается между клеточными компартментами памяти и образующими антитело клетками в первичном иммунном ответе. EMBO J . (1997) 16: 2996–3006. DOI: 10.1093 / emboj / 16.11.2996
PubMed Аннотация | CrossRef Полный текст | Google Scholar
11. Tarte K, Zhan F, De Vos J, Klein B, Shaughnessy J. Профилирование экспрессии генов плазматических клеток и плазмобластов: к лучшему пониманию поздних стадий дифференцировки B-клеток. Кровь .(2003) 102: 592–600. DOI: 10.1182 / кровь-2002-10-3161
PubMed Аннотация | CrossRef Полный текст | Google Scholar
12. Кассезе Г., Арсе С., Хаузер А.Е., Ленерт К., Моуэс Б., Манц Р.А. Выживание плазматических клеток опосредуется синергическим действием цитокинов и зависимых от адгезии сигналов. Дж Иммунол . (2003) 171: 1684–90. DOI: 10.4049 / jimmunol.171.4.1684
PubMed Аннотация | CrossRef Полный текст | Google Scholar
13. Шапиро-Шелеф М., Линь К.И., МакХейзер-Вильямс Л.Дж., Ляо Дж., МакХейзер-Вильямс М.Г., Каламе К.Blimp-1 необходим для образования плазматических клеток, секретирующих иммуноглобулин, и пре-плазменных В-клеток памяти. Иммунитет . (2003) 19: 607–20. DOI: 10.1016 / S1074-7613 (03) 00267-X
PubMed Аннотация | CrossRef Полный текст | Google Scholar
14. Валор Л.М., Родригес-Байона Б., Рамос-Амайя А.Б., Бриева Дж.А., Кампос-Каро А. Транскрипционное профилирование человеческих in vivo плазматических клеток, генерируемых , выявляет избирательный дисбаланс при моноклональных гаммопатиях. PLoS ONE .(2017) 12: e0183264. DOI: 10.1371 / journal.pone.0183264
PubMed Аннотация | CrossRef Полный текст | Google Scholar
15. Кассамбара А., Рем Т., Мишель Дж., Тьерри Ф., Дирк Х., Карин Т. и др. GenomicScape: простой в использовании веб-инструмент для анализа данных экспрессии генов. Приложение для исследования молекулярных событий при дифференцировке В-клеток в плазматические клетки. PLoS Comput Biol. (2015) 11: e1004077 – e77. DOI: 10.1371 / journal.pcbi.1004077
PubMed Аннотация | CrossRef Полный текст | Google Scholar
16.О’Коннор Б.П., Глисон М.В., Ноэль Р.Дж., Эриксон Л.Д. Взлет и падение долгоживущего гуморального иммунитета: терминальная дифференцировка плазматических клеток при здоровье и болезни. Immunol Ред. . (2003) 194: 61–76. DOI: 10.1034 / j.1600-065X.2003.00055.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
17. Нгуен, округ Колумбия, Гарималла С., Сяо Х., Кю С., Альбизуа I, Галипо Дж. И др. Факторы микроничества костного мозга, поддерживающие выживаемость плазматических клеток человека и секрецию иммуноглобулинов. Нац Коммуна . (2018) 9: 3698. DOI: 10.1038 / s41467-018-05853-7
PubMed Аннотация | CrossRef Полный текст | Google Scholar
19. Эллиард Д.И., Эйвери Д.Т., Фан Т.Г., Хейр Нью-Джерси, Ходжкин П.Д., Танге С.Г. Отобранные антигеном, секретирующие иммуноглобулин клетки сохраняются в селезенке и костном мозге человека. Кровь . (2004) 103: 3805–12. DOI: 10.1182 / кровь-2003-09-3109
PubMed Аннотация | CrossRef Полный текст | Google Scholar
20. Боханнон К., Пауэрс Р., Сатьябхама Л., Цуй А., Типтон С., Михаэли М. и др.Долгоживущие антиген-индуцированные плазматические клетки IgM демонстрируют соматические мутации и вносят свой вклад в долгосрочную защиту. Нац Коммуна . (2016) 7: 12687. DOI: 10.1038 / ncomms12687
CrossRef Полный текст | Google Scholar
21. Лемке А., Крафт М., Рот К., Ридель Р., Ламмердинг Д., Хаузер А. Долгоживущие плазматические клетки образуются в иммунных ответах слизистых оболочек и вносят вклад в пул плазматических клеток костного мозга у мышей. Иммунол слизистой оболочки . (2016) 9: 83–97. DOI: 10,1038 / ми.2015,38
PubMed Аннотация | CrossRef Полный текст | Google Scholar
22. Cassese G, Lindenau S, de Boer B, Arce S, Hauser A, Riemekasten GA. Воспаленные почки мышей NZB / W являются основным местом гомеостаза плазматических клеток. евро J Immunol . (2001) 31: 2726–32. DOI: 10.1002 / 1521-4141 (200109) 31: 9 <2726 :: AID-IMMU2726> 3.0.CO; 2-H
CrossRef Полный текст | Google Scholar
23. Manz RA, Arce S, Cassese G, Hauser A.E, Hiepe F, Radbruch A.Гуморальный иммунитет и долгоживущие плазматические клетки. Curr Opin Immunol . (2002) 14: 517–21. DOI: 10.1016 / S0952-7915 (02) 00356-4
PubMed Аннотация | CrossRef Полный текст | Google Scholar
24. Landsverk OJ, Snir O, Casado RB, Richter L, Mold JE, Jahnsen FL. Плазматические клетки, секретирующие антитела, десятилетиями сохраняются в кишечнике человека. J Exp Med . (2017) 214: 309–17. DOI: 10.1084 / jem.20161590
PubMed Аннотация | CrossRef Полный текст | Google Scholar
25.Brandtzaeg P, Johansen FE. В-клетки слизистой оболочки: фенотипические характеристики, регуляция транскрипции и свойства самонаведения. Immunol Ред. . (2005) 206: 32–63. DOI: 10.1111 / j.0105-2896.2005.00283.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
26. Каваниши Х., Зальцман Л. Е., Стробер В. Механизмы, регулирующие выработку иммуноглобулинов, специфичных для класса IgA, в лимфоидных тканях, ассоциированных с кишечником мыши. I. Т-клетки, полученные из пейерсовских пятен, которые переключают sIgM B-клетки на sIgA B-клетки in vitro.J Exp Med . (1983) 157: 433–50. DOI: 10.1084 / jem.157.2.433
PubMed Аннотация | CrossRef Полный текст | Google Scholar
27. Dullaers M, Li D, Xue Y, Ni L, Gayet I., Morita R, Oh S. Зависимый от Т-клеток механизм индукции плазмобластов, секретирующих человеческий иммуноглобулин A в слизистой оболочке. Иммунитет . (2009) 30: 120–9. DOI: 10.1016 / j.immuni.2008.11.008
PubMed Аннотация | CrossRef Полный текст | Google Scholar
28. Hapfelmeier S, Lawson MA, Slack E, Kirundi JK, Stoel M, Macpherson AJ.Обратимая микробная колонизация мышей, свободных от микробов, выявляет динамику иммунных ответов IgA. Наука . (2010) 328: 1705–9. DOI: 10.1126 / science.1188454
PubMed Аннотация | CrossRef Полный текст | Google Scholar
29. Месин Л., Ди Ниро Р., Томпсон К. М., Лундин К. Э., Соллид Л. М.. Долгоживущие плазматические клетки из биопсий тонкого кишечника человека секретируют иммуноглобулины в течение многих недель in vitro . Дж Иммунол . (2011) 187: 2867–74. DOI: 10.4049 / джиммунол.1003181
PubMed Аннотация | CrossRef Полный текст | Google Scholar
30. Mei HE, Yoshida T, Sime W., Hiepe F, Thiele K, Manz R, Dorner T. Переносимые с кровью плазматические клетки человека в устойчивом состоянии получены из иммунных ответов слизистых оболочек. Кровь . (2009) 113: 2461–69. DOI: 10.1182 / кровь-2008-04-153544
PubMed Аннотация | CrossRef Полный текст | Google Scholar
31. Люгер Э.О., Фокул В., Вегманн М., Абрам М., Тиллак К., Ахатц Г., Ренц Х. Индукция долгоживущих аллергенспецифических плазматических клеток путем заражения аллергеном слизистой оболочки. J Allergy Clin Immunol . (2009) 124: 819–26.e4. DOI: 10.1016 / j.jaci.2009.06.047
PubMed Аннотация | CrossRef Полный текст | Google Scholar
33. Medina F, Segundo C, Campos-Caro A, Gonzalez-Garcia I, Brieva JA. Неоднородность, демонстрируемая человеческими плазматическими клетками миндалин, крови и костного мозга, выявляет ступенчатые стадии увеличения зрелости, но локальные профили экспрессии молекул адгезии. Кровь . (2002) 99: 2154–61. DOI: 10.1182 / blood.V99.6.2154
PubMed Аннотация | CrossRef Полный текст | Google Scholar
34.О’Коннор Б.П., Каскальо М., Ноэль Р.Дж. Короткоживущие и долгоживущие плазматические клетки костного мозга получены из новой популяции предшественников. J Exp Med . (2002) 195: 737–45. DOI: 10.1084 / jem.20011626
PubMed Аннотация | CrossRef Полный текст | Google Scholar
35. Цинкернагель Р.М., Бахманн М.Ф., Кундиг Т.М., Оехен С., Пирше Х., Хенгартнер Х. Об иммунологической памяти. Анну Рев Иммунол . (1996) 14: 333–67. DOI: 10.1146 / annurev.immunol.14.1.333
CrossRef Полный текст | Google Scholar
36.Schuurman HJ, Verdonck LF, Geertzema JG, van der Linden JA, de Gast GC. Монотипические плазматические клетки иммуноглобулина E у реципиента аллогенного трансплантата костного мозга. Гистопатология. (1986) 10: 963–9. DOI: 10.1111 / j.1365-2559.1986.tb02594.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
37. Абеди М.Р., Бакман Л., Перссон Ю., Рингден О. Уровни IgE в сыворотке после трансплантации костного мозга. Пересадка костного мозга . (1989) 4: 255–60.
PubMed Аннотация | Google Scholar
38.Hammarlund E, Thomas A, Amanna IJ, Holden LA, Slayden OD, Slifka MK. Выживание плазматических клеток при отсутствии памяти В-клеток. Нац Коммуна . (2017) 8: 1781. DOI: 10.1038 / s41467-017-01901-w
PubMed Аннотация | CrossRef Полный текст | Google Scholar
40. Ахуджа А., Андерсон М.С., Халил А., Шломчик Дж. Поддержание пула плазматических клеток не зависит от В-клеток памяти. Proc Natl Acad Sci USA . (2008) 105: 4802–7. DOI: 10.1073 / pnas.0800555105
PubMed Аннотация | CrossRef Полный текст | Google Scholar
41.DiLillo DJ, Hamaguchi Y, Ueda Y, Yang K, Uchida J, Haas K, Tedder TF. Поддержание долгоживущих плазматических клеток и серологической памяти, несмотря на истощение зрелых и В-клеток памяти во время иммунотерапии CD20 у мышей. Дж Иммунол . (2007) 180: 361–71. DOI: 10.4049 / jimmunol.180.1.361
PubMed Аннотация | CrossRef Полный текст | Google Scholar
42. Bhoj VG, Arhontoulis D, Wertheim G, Capobianchi J, Callahan CA, Milone M. Сохранение долгоживущих плазматических клеток и гуморальный иммунитет у людей, отвечающих на терапию CD19-направленными CAR Т-клетками. Кровь . (2016) 128: 360–70. DOI: 10.1182 / кровь-2016-01-694356
PubMed Аннотация | CrossRef Полный текст | Google Scholar
43. Mei HE, Wirries I, Frolich D, Brisslert M, Giesecke C, Grun J, Dorner T. Уникальная популяция IgG-экспрессирующих плазматических клеток, лишенных CD19, обогащена костным мозгом человека. Кровь . (2015) 125: 1739–48. DOI: 10.1182 / кровь-2014-02-555169
PubMed Аннотация | CrossRef Полный текст | Google Scholar
44. Андерхилл Г.Х., Джордж Д., Бремер Э.Г., Канзас Г.С.Профили экспрессии генов показывают высокоспециализированную генетическую программу плазматических клеток. Кровь . (2003) 101: 4013–21. DOI: 10.1182 / кровь-2002-08-2673
PubMed Аннотация | CrossRef Полный текст | Google Scholar
45. Халлили Дж. Л., Типтон С. М., Лисвельд Дж., Розенберг А. Ф., Дарс Дж., Ли Ф. Э. Долгоживущие плазматические клетки содержатся в субпопуляции CD19 (-) CD38 (hi) CD138 (+) в костном мозге человека. Иммунитет . (2015) 43: 132–45. DOI: 10.1016 / j.immuni.2015.06.016
PubMed Аннотация | CrossRef Полный текст | Google Scholar
46.Reynolds AE, Kuraoka M, Kelsoe G. Природный IgM продуцируется CD5-плазматическими клетками, которые занимают особую нишу выживания в костном мозге. Дж Иммунол . (2015) 194: 231–42. DOI: 10.4049 / jimmunol.1401203
PubMed Аннотация | CrossRef Полный текст | Google Scholar
47. Бенсон М.Дж., Диллон С.Р., Кастильи Э., Геха Р.С., Сюй С., Ноэль Р.Дж. Передний край: зависимость плазматических клеток и независимость В-клеток памяти от BAFF и APRIL. Дж Иммунол . (2008) 180: 3655–9. DOI: 10.4049 / джиммунол.180.6.3655
PubMed Аннотация | CrossRef Полный текст | Google Scholar
48. Minges Wols HA, Ippolito JA, Yu Z, Palmer JL, White FA, Witte PL. Влияние микросреды и внутреннего программирования на выживаемость плазматических клеток. Инт Иммунол . (2007) 19: 837–46. DOI: 10.1093 / intimm / dxm051
PubMed Аннотация | CrossRef Полный текст | Google Scholar
49. Jourdan M, Cren M, Robert N, Bollore K, Fest T, Duperray C. IL-6 поддерживает образование долгоживущих плазматических клеток человека в сочетании с APRIL или факторами, растворимыми в стромальных клетках. Лейкемия. (2014) 28: 1647–56. DOI: 10.1038 / leu.2014.61
CrossRef Полный текст | Google Scholar
50. О’Коннор Б.П., Раман В.С., Эриксон Л.Д., Кук В.Дж., Уивер Л.К., Ноэль Р.Дж. BCMA необходим для выживания долгоживущих плазматических клеток костного мозга. J Exp Med . (2004) 199: 91–8. DOI: 10.1084 / jem.20031330
PubMed Аннотация | CrossRef Полный текст | Google Scholar
52. Tai YT, Acharya C, An G, Moschetta M, Zhong MY, Anderson KC. APRIL и BCMA способствуют росту множественной миеломы человека и иммуносупрессии в микросреде костного мозга. Кровь . (2016) 127: 3225–36. DOI: 10.1182 / кровь-2016-01-6
PubMed Аннотация | CrossRef Полный текст | Google Scholar
53. Чо С.Ф., Андерсон К.С., Тай Ю.Т. Нацеленность на антиген созревания В-клеток (BCMA) при множественной миеломе: потенциальное использование иммунотерапии на основе BCMA. Фронт Иммунол . (2018) 9: 1821. DOI: 10.3389 / fimmu.2018.01821
PubMed Аннотация | CrossRef Полный текст | Google Scholar
55. Деренн С., Мония Б., Дин Н.М., Тейлор Дж. К., Рапп М.Дж., Амиот М.Антисмысловая стратегия показывает, что Mcl-1, а не Bcl-2 или Bcl-x (L), является важным белком выживания клеток миеломы человека. Кровь . (2002) 100: 194–9. DOI: 10.1182 / blood.V100.1.194
PubMed Аннотация | CrossRef Полный текст | Google Scholar
58. Кавано М.М., Михара К., Хуанг Н., Цудзимото Т., Курамото А. Дифференциация ранних плазматических клеток на стромальных клетках костного мозга требует интерлейкина-6 для выхода из апоптоза. Кровь . (1995) 85: 487–94.
PubMed Аннотация | Google Scholar
59.Choe J, Choi Y S. IL-10 прерывает экспансию В-клеток памяти в зародышевом центре, индуцируя дифференцировку в плазматические клетки. евро J Immunol . (1998) 28: 508–15. DOI: 10.1002 / (SICI) 1521-4141 (199802) 28:02 <508 :: AID-IMMU508> 3.0.CO; 2-I
PubMed Аннотация | CrossRef Полный текст | Google Scholar
60. Ettinger R, Sims GP, Fairhurst AM, Robbins R, da Silva YS, Lipsky PE. IL-21 индуцирует дифференцировку наивных В-клеток человека и В-клеток памяти в плазматические клетки, секретирующие антитела. Дж Иммунол . (2005) 175: 7867–79. DOI: 10.4049 / jimmunol.175.12.7867
PubMed Аннотация | CrossRef Полный текст | Google Scholar
61. Дин ББ, Би Э, Чен Х, Ю ДжДж, Йе БХ. IL-21 и CD40L синергетически способствуют дифференцировке плазматических клеток за счет активации Blimp-1 в B-клетках человека. Дж Иммунол . (2013) 190: 1827–36. DOI: 10.4049 / jimmunol.1201678
PubMed Аннотация | CrossRef Полный текст | Google Scholar
62. Брайант В.Л., Ма К.С., Эйвери Д.Т., Ли Й., Гуд КЛГ.Цитокин-опосредованная регуляция дифференцировки человеческих B-клеток в Ig-секретирующие клетки: преобладающая роль IL-21, продуцируемого CXCR5 + T фолликулярными вспомогательными клетками. Дж Иммунол . (2007) 179: 8180–90. DOI: 10.4049 / jimmunol.179.12.8180
PubMed Аннотация | CrossRef Полный текст | Google Scholar
64. Родригес-Байона Б., Рамос-Амая А., Бернал Дж., Кампос-Каро А., Бриева Ю.А. Передний край: ИЛ-21, полученный из фолликулярных хелперных Т-клеток человека, действует как фактор выживания для вторичных лимфоидных органов, но не для костного мозга и плазматических клеток. Дж Иммунол . (2012) 188: 1578–81. DOI: 10.4049 / jimmunol.1102786
CrossRef Полный текст | Google Scholar
65. Рамос-Амайя А., Родригес-Байона Б., Лопес-Бланко Р., Андухар Е., Перес-Алегре М., Кампос-Каро А. Выживание человеческих циркулирующих антиген-индуцированных плазматических клеток поддерживается нишевыми цитокинами плазматических клеток и Т. фолликулярные лимфоциты-помощники. Дж Иммунол . (2015) 194: 1031–8. DOI: 10.4049 / jimmunol.1402231
PubMed Аннотация | CrossRef Полный текст | Google Scholar
66.Kwon H, Thierry-Mieg D, Thierry-Mieg J, Kim HP, Oh J, Leonard WJ. Анализ регуляции гена Prdm1, индуцированной интерлейкином-21, показывает функциональную кооперацию факторов транскрипции STAT3 и IRF4. Иммунитет . (2009) 31: 941–52. DOI: 10.1016 / j.immuni.2009.10.008
PubMed Аннотация | CrossRef Полный текст | Google Scholar
67. Эйвери Д. Т., Диник Е. К., Ма К. С., Сурьяни С., Симпсон Н., Танье С. Г.. Внутренняя передача сигналов В-клеткам через рецептор IL-21 и STAT3 необходима для установления долгоживущих ответов антител у людей. J Exp Med . (2010) 207: 155–71. DOI: 10.1084 / jem.200
PubMed Аннотация | CrossRef Полный текст | Google Scholar
68. Родригес-Байона Б., Рамос-Амайя А., Лопес-Бланко Р., Кампос-Каро А., Бриева Дж. А. Активация STAT-3 дифференциальными цитокинами имеет решающее значение для выживания плазматических клеток человека in vivo, генерируемых , и секреции Ig. . Дж Иммунол . (2013) 191: 4996–5004. DOI: 10.4049 / jimmunol.1301559
PubMed Аннотация | CrossRef Полный текст | Google Scholar
70.Ван Дж. Х., Авиталь Н., Кариаппа А., Фридрих С., Икеда Т., Ренольд А. и др. Aiolos регулирует активацию и созревание В-клеток до эффекторного состояния. Иммунитет . (1998) 9: 543–53. DOI: 10.1016 / S1074-7613 (00) 80637-8
PubMed Аннотация | CrossRef Полный текст | Google Scholar
71. Cortes M, Georgopoulos K. Aiolos необходим для генерации высокоаффинных плазматических клеток костного мозга, ответственных за долговременный иммунитет. J Exp Med . (2004) 199: 209–19. DOI: 10.1084 / jem.20031571
PubMed Аннотация | CrossRef Полный текст | Google Scholar
73. Шеврие С., Гентон С., Каллис А., Карновски А., Оттен Л., Орбеа А. CD93 необходим для поддержания секреции антител и сохранения плазматических клеток в нише костного мозга. Proc Natl Acad Sci USA . (2009) 106: 3895–900. DOI: 10.1073 / pnas.0809736106
PubMed Аннотация | CrossRef Полный текст | Google Scholar
74. Хараламбьева И.Х., Циммерманн М.Т., Овсянникова И.Г., Grill DE, Оберг А.Л., Польша GA.Профиль полного транскриптома позволяет идентифицировать гены CD93 и других факторов выживаемости плазматических клеток, связанные со специфическим ответом антител к кори после вакцинации. PLoS ONE . (2016) 11: e0160970. DOI: 10.1371 / journal.pone.0160970
PubMed Аннотация | CrossRef Полный текст | Google Scholar
75. Delogu A, Schebesta A, Sun Q, Aschenbrenner K, Perlot T, Busslinger M. Репрессия генов с помощью Pax5 в B-клетках важна для гомеостаза клеток крови и в плазматических клетках обратная. Иммунитет . (2006) 24: 269–81. DOI: 10.1016 / j.immuni.2006.01.012
PubMed Аннотация | CrossRef Полный текст | Google Scholar
76. Ferguson SE, Han S, Kelsoe G, Thompson CB. CD28 необходим для образования зародышевого центра. Дж Иммунол . (1996) 156: 4576–81.
PubMed Аннотация | Google Scholar
77. Шагинян А., Пфеффер К., Ли К.П., Кундиг Т.М., Кишихара К., Мак Т.В. Дифференциальные костимулирующие потребности Т-лимфоцитов у мышей с дефицитом CD28. Наука . (1993) 261: 609–12. DOI: 10.1126 / science.7688139
PubMed Аннотация | CrossRef Полный текст | Google Scholar
78. Додсон Л.Ф., Бумер Дж. С., Деппонг С. М., Шах Д. Д., Сим Дж., Грин Дж. М.. Мыши с целевым «нокаутом», экспрессирующие мутации CD28, обнаруживают важный путь костимуляции. Mol Cell Biol . (2009) 29: 3710–21. DOI: 10.1128 / MCB.01869-08
PubMed Аннотация | CrossRef Полный текст | Google Scholar
79. Друг Л.Д., Шах Д.Д., Деппонг К., Лин Дж., Брикер Т.Л., Грин Дж. М..Дозозависимая потребность в пролиновом мотиве CD28 в клеточном и гуморальном иммунитете, выявленная мутантом-направленным нокином. J Exp Med . (2006) 203: 2121–33. DOI: 10.1084 / jem.20052230
PubMed Аннотация | CrossRef Полный текст | Google Scholar
80. Розанский С.Х., Аренс Р., Карлсон Л.М., Наир Дж., Бойсе Л.Х., Ли КП. Устойчивые ответы антител зависят от функции CD28 в плазматических клетках, резидентных в костном мозге. J Exp Med . (2011) 208: 1435–46. DOI: 10.1084 / jem.20110040
PubMed Аннотация | CrossRef Полный текст | Google Scholar
81. Njau MN, Kim JH, Chappell CP, Ravindran R, Thomas L, Jacob J. Взаимодействие CD28-B7 модулирует функцию короткоживущих и долгоживущих плазматических клеток. Дж Иммунол . (2012) 189: 2758–67. DOI: 10.4049 / jimmunol.1102728
PubMed Аннотация | CrossRef Полный текст | Google Scholar
82. Розански С.Х., Атли А., Карлсон Л.М., Фаррен М.Р., Мюррей М., Ли КП. CD28 способствует выживанию плазматических клеток, устойчивым ответам антител и активации BLIMP-1 через его дистальный пролиновый мотив PYAP. Дж Иммунол . (2015) 194: 4717–28. DOI: 10.4049 / jimmunol.1402260
PubMed Аннотация | CrossRef Полный текст | Google Scholar
83. Теллиер Дж., Ши В., Минних М., Ляо И., Кроуфорд С., Смит Дж., Натт С.Л. Blimp-1 контролирует функцию плазматических клеток посредством регуляции секреции иммуноглобулинов и развернутого белкового ответа. Нат Иммунол . (2016) 17: 323–30. DOI: 10.1038 / ni.3348
PubMed Аннотация | CrossRef Полный текст | Google Scholar
84.Мюррей М.Э., Гэвил С.М., Наир Дж. Р., Курелла С., Карлсон Л. М., Ли КП. CD28-опосредованная передача сигналов, способствующих выживанию, индуцирует химиотерапевтическую резистентность при множественной миеломе. Кровь . (2014) 123: 3770–9. DOI: 10.1182 / кровь-2013-10-530964
PubMed Аннотация | CrossRef Полный текст | Google Scholar
85. Наир Дж. Р., Карлсон Л. М., Курелла С., Розански С. К., Бирн Г. Е., Ли К. П.. CD28, экспрессируемый на злокачественных плазматических клетках, индуцирует выживание и иммуносупрессивное микроокружение. Дж Иммунол .(2011) 187: 1243–53. DOI: 10.4049 / jimmunol.1100016
PubMed Аннотация | CrossRef Полный текст | Google Scholar
86. Курелла С., Наир Дж. Р., Мюррей М. Е., Карлсон Л. М., Уоткинс С. К., Ли К. П.. Новая регуляция CD80 / CD86-индуцированной передачи сигналов фосфатидилинозитол-3-киназы с помощью белка NOTCh2 в продукции интерлейкина-6 и индоламин-2,3-диоксигеназы дендритными клетками. Дж. Биол. Хим. . (2014) 289: 7747–62. DOI: 10.1074 / jbc.M113.519686
PubMed Аннотация | CrossRef Полный текст | Google Scholar
87.Гасс Дж. Н., Гиффорд Н. М., Брюэр Дж. В.. Активация развернутого белкового ответа во время дифференцировки В-клеток, секретирующих антитела. Дж. Биол. Хим. . (2002) 277: 49047–54. DOI: 10.1074 / jbc.M205011200
PubMed Аннотация | CrossRef Полный текст | Google Scholar
89. Пуа Х. Х., Иван Д., Мариана С., Нобору М., Ю-Вен Х. Критическая роль гена аутофагии Atg5 в выживании и пролиферации Т-клеток. J Exp Med. (2007) 204: 25. DOI: 10.1084 / jem.20061303
PubMed Аннотация | CrossRef Полный текст | Google Scholar
92.Панди У.Б., Не З., Батлеви Й., МакКрей Б.А., Ритсон Г.П., Тейлор Дж. П.. HDAC6 спасает нейродегенерацию и обеспечивает важную связь между аутофагией и ИБП. Природа . (2007) 447: 859–63. DOI: 10.1038 / nature05853
PubMed Аннотация | CrossRef Полный текст | Google Scholar
94. Пенго Н., Сколари М., Олива Л., Милан Э, Майнольди Ф., Раймонди А. Плазменные клетки нуждаются в аутофагии для устойчивого производства иммуноглобулинов. Нат Иммунол . (2013) 14: 298–305. DOI: 10.1038 / ni.2524
CrossRef Полный текст | Google Scholar
95. О’Салливан Д., ван дер Виндт Г.Дж., Хуанг С.К., Кертис Д.Д., Чанг С.Х., Пирс Э.Л. Память CD8 (+) Т-клетки используют внутренний липолиз для поддержки метаболического программирования, необходимого для развития. Иммунитет . (2014) 41: 75–88. DOI: 10.1016 / j.immuni.2014.06.005
PubMed Аннотация | CrossRef Полный текст | Google Scholar
96. Бак MD, OSullivan D, Klein Geltink RI, Curtis JD, Chang CH, Pearce EL.Митохондриальная динамика контролирует судьбу Т-клеток посредством метаболического программирования. Ячейка . (2016) 166: 63–76. DOI: 10.1016 / j.cell.2016.05.035
PubMed Аннотация | CrossRef Полный текст | Google Scholar
97. Лам В.Й., Беккер А.М., Кеннерли К.М., Вонг Р., Кертис Д.Д., Бхаттачарья Д. Импорт митохондриального пирувата способствует долгосрочному выживанию плазматических клеток, секретирующих антитела. Иммунитет . (2016) 45: 60–73. DOI: 10.1016 / j.immuni.2016.06.011
PubMed Аннотация | CrossRef Полный текст | Google Scholar
98.Lam WY, Jash A, Yao CH, DSouza L, Wong R, Bhattacharya D. Метаболические и транскрипционные модули независимо друг от друга диверсифицируют продолжительность жизни и функции плазматических клеток. Cell Rep . (2018) 24: 2479–92.e6. DOI: 10.1016 / j.celrep.2018.07.084
PubMed Аннотация | CrossRef Полный текст | Google Scholar
99. Kunisawa J, Sugiura Y, Wake T, Nagatake T, Suzuki H, Nagasawa R. Режим биоэнергетического метаболизма во время дифференцировки B-клеток в кишечнике определяет особую потребность в витамине B1. Cell Rep . (2015) 13: 122–31. DOI: 10.1016 / j.celrep.2015.08.063
PubMed Аннотация | CrossRef Полный текст | Google Scholar
100. Като К., Нишимасу Х., Окудаира С., Михара Е., Иситани Р., Такаги Дж. Кристаллическая структура Enpp1, внеклеточного гликопротеина, участвующего в минерализации костей и передаче сигналов инсулина. Proc Natl Acad Sci USA . (2012) 109: 16876–81. DOI: 10.1073 / pnas.1208017109
PubMed Аннотация | CrossRef Полный текст | Google Scholar
101.Ван Х., Гонсалес-Гарсия I, Траба Дж., Джайн С., Конте С., Шин Д. и др. Разлагающий АТФ ENPP1 необходим для выживания (или сохранения) долгоживущих плазматических клеток. Научный сотрудник . (2017) 7: 17867. DOI: 10.1038 / s41598-017-18028-z
PubMed Аннотация | CrossRef Полный текст | Google Scholar
102. Minges Wols HA, Underhill GH, Kansas GS, Witte PL. Роль стромальных клеток костного мозга в поддержании долголетия плазматических клеток. Дж Иммунол . (2002) 169: 4213–21.DOI: 10.4049 / jimmunol.169.8.4213
PubMed Аннотация | CrossRef Полный текст | Google Scholar
103. Уилмор-младший, Оллман Д. Здесь, там и где угодно? Аргументы за и против физического выживания плазматических клеток Ниша. Дж Иммунол . (2017) 199: 839–45. DOI: 10.4049 / jimmunol.1700461
PubMed Аннотация | CrossRef Полный текст | Google Scholar
104. Мур Н., Морено Гонсалес М., Боннер К., Смит Б., Парк В., Стегал М. Влияние блокады CXCR4 / CXCL12 на нормальные плазматические клетки in vivo . Трансплантат Am J . (2017) 17: 1663–9. DOI: 10.1111 / ajt.14236
PubMed Аннотация | CrossRef Полный текст | Google Scholar
105. Дамиано Дж. С., Кресс А. Е., Хазлхерст Л. А., Штиль А. А., Далтон В. С.. Устойчивость к лекарственным средствам, опосредованная адгезией клеток (CAM-DR): роль интегринов и устойчивость к апоптозу в клеточных линиях миеломы человека. Кровь . (1999) 93: 1658–67.
PubMed Аннотация | Google Scholar
106. Винтер О., Мозер К., Мор Э, Зотос Д., Камински Х., Шиска М., Манц Р. А.Мегакариоциты представляют собой функциональный компонент ниши плазматических клеток в костном мозге. Кровь . (2010) 116: 1867–75. DOI: 10.1182 / кровь-2009-12-259457
PubMed Аннотация | CrossRef Полный текст | Google Scholar
107. Родригес Гомес М., Талке Ю., Гебель Н., Герман Ф., Райх Б., Мак М. Базофилы поддерживают выживание плазматических клеток у мышей. Дж Иммунол . (2010) 185: 7180–5. DOI: 10.4049 / jimmunol.1002319
PubMed Аннотация | CrossRef Полный текст | Google Scholar
108.Чу В.Т., Фролих А., Штайнхаузер Г., Шил Т., Рох Т., Филлатро С. и др. Эозинофилы необходимы для поддержания плазматических клеток в костном мозге. Нат Иммунол . (2011) 12: 151–9. DOI: 10.1038 / ni.1981
PubMed Аннотация | CrossRef Полный текст | Google Scholar
109. Brynjolfsson SF, Persson Berg L, Olsen Ekerhult T, Rimkute I, Wick MJ, Grimsholm O. Долгоживущие плазматические клетки у мышей и людей. Фронт Иммунол . (2018) 9: 2673. DOI: 10.3389 / fimmu.2018.02673
PubMed Аннотация | CrossRef Полный текст | Google Scholar
110. Чу В.Т., Беллер А., Рауш С., Страндмарк Дж., Занкер М., Арбах О. Эозинофилы способствуют образованию и поддержанию плазматических клеток, экспрессирующих иммуноглобулин-А, и способствуют гомеостазу иммунной системы кишечника. Иммунитет . (2014) 40: 582–93. DOI: 10.1016 / j.immuni.2014.02.014
PubMed Аннотация | CrossRef Полный текст | Google Scholar
112. Бортник А., Чернова И., Спенсер С.П., Оллман Д.Нет строгих требований к эозинофилам для выживания плазматических клеток костного мозга. евро J Immunol . (2018) 48: 815–21. DOI: 10.1002 / eji.201747229
PubMed Аннотация | CrossRef Полный текст | Google Scholar
113. Глатман Зарецкий А, Конрад С, Депис F, Крыло JB, Гоенка R, Хантер CA. Т-регуляторные клетки поддерживают популяции плазматических клеток в костном мозге. Cell Rep . (2017) 18: 1906–16. DOI: 10.1016 / j.celrep.2017.01.067
PubMed Аннотация | CrossRef Полный текст | Google Scholar
114.Сюй С, Лам КП. Белок созревания В-клеток, который связывает члены семейства факторов некроза опухолей BAFF и APRIL, незаменим для гуморальных иммунных ответов. Mol Cell Biol. (2001) 21: 4067–74. DOI: 10.1128 / MCB.21.12.4067-4074.2001
PubMed Аннотация | CrossRef Полный текст | Google Scholar
115. Gustafson CE, Higbee D, Yeckes AR, Wilson CC, De Zoeten EF, Jedlicka P. Ограниченная экспрессия APRIL и его рецепторов до развития кишечных плазматических клеток IgA в младенчестве человека. Mucosal Immunol. (2013) 7: 467. DOI: 10,1038 / mi.2013.64
PubMed Аннотация | CrossRef Полный текст | Google Scholar
117. Маккирн Дж. П., Баум К., Дэви Дж. М.. Антигены клеточной поверхности экспрессируются подмножествами пре-В-клеток и В-клеток. J Immunol. (1984) 132: 332–9.
PubMed Аннотация | Google Scholar
118. Gärdby E, Wrammert J, Schön K, Ekman L, Leanderson T., Lycke N. Сильная дифференциальная регуляция сывороточных и слизистых IgA-ответов, выявленных у мышей с дефицитом CD28 с использованием адъюванта холерного токсина. J Immunol. (2003) 170: 55–63. DOI: 10.4049 / jimmunol.170.1.55
PubMed Аннотация | CrossRef Полный текст | Google Scholar
119. Арнольд Дж., Мурера Д., Арбогаст Ф., Фауни Дж. Д., Мюллер С., Гро Ф. Аутофагия необязательна для развития В-клеток, но необходима для гуморальных аутоиммунных реакций. Cell Death Differ. (2016) 23: 853–64. DOI: 10.1038 / cdd.2015.149
PubMed Аннотация | CrossRef Полный текст | Google Scholar
122. Уотерс Л. Р., Ахсан Ф. М., Вольф Д. М., Ширихай О., Тейтелл М. А..Первоначальная активация В-клеток вызывает метаболическое перепрограммирование и ремоделирование митохондрий. iScience. (2018) 5: 99–109. DOI: 10.1016 / j.isci.2018.07.005
PubMed Аннотация | CrossRef Полный текст | Google Scholar
123. Юн Дж., Ван Х., Ким Ю.С., Ёсимото М., Аббаси С., Морс Х.С. III. Аллоантиген ENPP1 плазматических клеток экспрессируется подмножеством В-клеток человека с потенциальными регуляторными функциями. Immunol Cell Biol. (2016) 94: 719–28. DOI: 10.1038 / icb.2016.31
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Триптолид является ингибитором РНК-полимеразы I и II-зависимой транскрипции, ведущей преимущественно к подавлению короткоживущей мРНК
Abstract
Триптолид, натуральный продукт, экстрагированный из китайского растения Tripterygium wilfordii , обладает противоопухолевыми свойствами.Несмотря на многочисленные сообщения, показывающие проапоптотическую способность и ингибирование NF-κB-опосредованной транскрипции триптолидом, идентичность его клеточной мишени все еще неизвестна. Чтобы прояснить его механизм действия, мы дополнительно исследовали влияние триптолида на синтез РНК в клеточной линии немелкоклеточного рака легкого человека A549. Триптолид подавлял синтез как общей РНК, так и мРНК de novo , причем основное действие оказывалось на последний пул. Мы использовали 44K микромассивов пангеномной ДНК человека и идентифицировали гены, на которые в первую очередь влияет кратковременное лечение триптолидом.Среди модулированных генов до 98% подавляются, включая большой набор онкогенов, включая факторы транскрипции и регуляторы клеточного цикла. Затем мы наблюдали, что триптолид индуцировал быстрое истощение RPB1, основной субъединицы РНК-полимеразы II, которая считается отличительным признаком блокировки элонгации транскрипции. Однако мы также показали, что триптолид не взаимодействует напрямую с комплексом РНК-полимеразы II и не повреждает ДНК. Таким образом, мы пришли к выводу, что триптолид является оригинальным фармакологическим ингибитором активности РНК-полимеразы, косвенно влияющим на аппарат транскрипции, что приводит к быстрому истощению короткоживущей мРНК, включая факторы транскрипции, регуляторы клеточного цикла, такие как CDC25A, и онкогены MYC и Src.В целом, данные проливают свет на влияние триптолида на транскрипцию, а также на его новые потенциальные применения при раке, включая острый миелоидный лейкоз, который частично обусловлен вышеупомянутыми онкогенными факторами. [Mol Cancer Ther 2009; 8 (10): 2780–90]
Ключевые слова- триптолид
- Tripterygium wilfordii
- транскрипция
- ядрышко
- РНК-полимераза II
- РНК-полимераза I 924
- РНК-полимераза I 924 Анализ
- Myc
Введение
Триптолид — натуральный продукт, извлеченный из китайского растения Tripterygium wilfordii , который используется в традиционной китайской медицине для лечения иммунных воспалительных заболеваний (1, 2).Его мощная антипролиферативная активность in vitro побудила его провести оценку опухолей человека, ксенотрансплантатов у мышей, где он четко продемонстрировал противоопухолевую активность (3-5). Интересно, что триптолид вызвал клинические реакции при тестировании в фазе I клинического исследования у пациентов с лейкемией (6). Многочисленные сообщения показывают, что триптолид ингибирует транскрипцию, опосредованную NF-κB, что было представлено в качестве основы его противовоспалительного действия, а также его антипролиферативных и проапоптотических свойств (7–9).Последние данные показывают, что триптолид ингибирует транскрипцию, опосредованную фактором теплового шока 1 (HSF1), а также факторами белка-активатора-1 (10, 11). McCallum et al. показали, что триптолид ингибирует de novo тотальную транскрипцию РНК в опухолевых клетках A549 и THP-1 (10). Поскольку рРНК составляет> 80% всей РНК (12), последнее исследование предполагает, что триптолид ингибирует производство этого пула РНК. Однако потенциальная активность этой молекулы в пуле мРНК широко не исследовалась.Совсем недавно Leuenroth и Crews опубликовали данные, показывающие, что триптолид вызывает морфологические изменения ядра, то есть нарушение целостности ядрышка и округление ядерных спеклов, которые, как предполагалось, отражают глобальное влияние триптолида на транскрипцию (13). В том же исследовании перемещение нуклеолина и SC35, соответственно, рибосомного белка и фактора сплайсинга РНК, было подтверждено через 16 часов после добавления соединения. Кроме того, было показано, что триптолид вызывает ингибирование фосфорилирования РНК-полимеразы II Ser 2 , что может объяснять ингибирование транскрипции мРНК.Фактически, анализ экспрессии генов раковых клеток человека Jurkat выявил недостаточную экспрессию 117 генов при лечении триптолидом (14). Но опять же, точный эффект триптолида на рРНК по сравнению с мРНК, наряду с его молекулярной мишенью (ями), еще предстоит выяснить. Все эти данные убедительно свидетельствуют о том, что триптолид является общим ингибитором транскрипции, который может модулировать больший набор генов, чем те, которые контролируются NF-κB, HSF1 и протеином-активатором-1, как первоначально предполагалось. Поскольку было показано, что триптолид действует ниже связывания NF-κB и HSF1 с ДНК, можно предположить, что он потенциально взаимодействует с аппаратом транскрипции рРНК и мРНК (7, 8, 11).Несколько мишеней могут объяснить свойства триптолида по ингибированию транскрипции. Во-первых, ДНК-топоизомераза I, благодаря своей способности декатенации, необходима для правильной транскрипции гена; действительно, камптотецин, специфический ингибитор топоизомеразы I, эффективно препятствует синтезу РНК (15, 16). Во-вторых, сообщалось, что взаимодействующие с ДНК молекулы, такие как актиномицин D, и агенты, повреждающие ДНК, включая УФ-излучение, блокируют транскрипцию (17-19). Наконец, α-аманитин, специфический ингибитор РНК-полимеразы II, действует посредством прямого взаимодействия с каталитической субъединицей RPB1 этого фермента (20–23).С целью выяснения механизма действия триптолида мы исследовали способность этого природного продукта вмешиваться в транскрипцию и взаимодействовать с вышеупомянутыми молекулярными мишенями. Таким образом, была исследована активность триптолида в отношении транскрипции, опосредованной внутриклеточной РНК-полимеразой I и II. Чтобы завершить это исследование, было оценено прямое взаимодействие между триптолидом и очищенным функциональным аппаратом транскрипции РНК-полимеразы II.
Материалы и методы
Клеточные линии и реагенты
Все клеточные линии были получены из Американской коллекции типовых культур, за исключением Miapaca2, MX1, G361, WM115 и WM266, которые были получены из Европейской коллекции клеточных культур.Эти клетки культивировали либо в среде DMEM (Life Technologies) для клеток COS, либо в RPMI 1640 (Life Technologies) для других, с добавлением 10% инактивированной нагреванием FCS (Sigma), пенициллина (65 мкг / мл) и стрептомицина (100 мкг / мл). мкг / мл). Человеческая РНК-полимераза II с субъединицей Rpb3, меченной FLAG, была очищена из клеток HEK293, как описано ранее (24). В каждом эксперименте использовали 4,3 пмоль полимеразы. Триптолид был получен от Pharmagenesis. Была проведена обширная химическая характеристика молекулы, включая инфракрасный спектр с преобразованием Фурье, ядерный магнитный резонанс 1 H и 13 C, элементный анализ (C и H), высокоэффективную жидкостную хроматографию, ТСХ и измерение температуры плавления (232 ° С).Чистота триптолида составляла 99,5%. Если не указано иное, все другие препараты и реагенты были от Sigma, за исключением α-аманитина, который был приобретен у Fluka.
Анализ пролиферации
Антипролиферативную активность тестируемых соединений измеряли с помощью анализа ATPlite (Perkin-Elmer) на комплексной панели из 16 клеточных линий. Прилипшие клетки высевали в 96-луночные планшеты (1 × 10 4 -2 × 10 4 / мл) в день 0, а затем инкубировали в течение 24 часов перед обработкой. Неприкрепленные клетки (Namalwa, COST, PIO, FEPD и Karpas-299) высевали в 96-луночные планшеты (3 × 10 5 / мл) в день 1; эти плотности посева были установлены для обеспечения того, чтобы клетки оставались в логарифмической фазе роста клеток в течение 72-часового периода обработки лекарственным средством.Все клетки обрабатывали в день 1; на 4-й день жизнеспособность клеток оценивали путем измерения АТФ, высвобождаемого жизнеспособными клетками. Значения ЕС 50 определяли с помощью анализа аппроксимации кривой (модель нелинейной регрессии с сигмоидальной зависимостью от дозы, переменный коэффициент наклона Хилла), выполненного с помощью алгоритма, предоставленного приложением GraphPad Prism (программное обеспечение GraphPad). Значения GI 50 для триптолида (номер доступа Национального института рака NSC163062), полученные на панели COMPARE NCI-60 линий раковых клеток, были получены по следующей ссылке: http: // dtp.nci.nih.gov/compare-web-public_compare/SearchAndDisplay (25). Сравнение профиля ингибирования роста триптолида с профилем противоопухолевых соединений было выполнено с помощью трехмерного инструмента MIND. 6
Измерения синтеза РНК
Клетки A549 выращивали до 70% слияния в 75 см 2 колб в течение 48 часов и предварительно инкубировали в течение 30 минут в присутствии различных концентраций тестируемого препарата с последующим 90-минутным маркированием РНК в присутствии 6 мкКи [H 3 ] уридина (GE Healthcare).Затем двадцать пять процентов клеток лизировали в воде для количественного определения общей РНК, а оставшиеся 75% использовали для экстракции мРНК с использованием набора MicroPoly (A) Purist (Ambion). Радиоактивность образцов определяли количественно на сцинтилляционном счетчике для микропланшетов 1450 MicroBeta Trilux (Perkin-Elmer). Чтобы оценить влияние триптолида как на 28S, так и на 18S рРНК, экспоненциально растущие 2 × 10 6 клеток A549 подвергали воздействию указанных концентраций триптолида в течение 8 часов и лизировали в 350 мкл лизирующего раствора, а общую РНК экстрагировали с помощью RNeasy Plus Mini. Набор (Qiagen) с последующим электрофорезом в агарозе.Затем РНК визуализировали с помощью бромистого этидия.
ДНК-микрочипы
Клетки A549 высевали в чашки диаметром 75 см 2 до 70% конфлюэнтности и подвергали воздействию триптолида в различных концентрациях в течение 1, 2 и 4 ч перед экстракцией РНК с использованием реагента TRizol (Invitrogen). Их качество оценивалось с помощью BioAnalyzer 2100 (Agilent Technologies) перед тем, как анализ экспрессии на основе одноцветной микроматрицы был проведен на 36 массивах [3 раза инкубации × 4 концентрации, включая контроль × 3 повтора].Каждый образец был помечен (Cy3, One Color Spike Mix, Agilent Technologies) и гибридизирован на микрочипах олигонуклеотидов 4 × 44K с полным геномом человека (45 015 зондов) в соответствии с рекомендациями производителя (Agilent Technologies). Флуоресцентные сигналы сканировались с параметрами по умолчанию (Agilent Technologies), а данные извлекались с помощью программного обеспечения Feature Extraction 9.1 (Agilent Technologies). Очистка данных заключалась в вычитании фона. После отбора по критерию «значительно выше фона» для анализа оставалось 30 499 проб.Чтобы идентифицировать изменения экспрессии зонда при различных концентрациях и временных условиях, средний сигнал анализировали с помощью двухфакторного дисперсионного анализа (система SAS R9.1), включающего концентрацию и время; Коэффициент ложного обнаружения был использован для корректировки множественных сравнений, в результате чего был получен список из 4 436 генов (5 133 зонда), нерегулируемых триптолидом. Управление данными микрочипов осуществлялось с помощью Arraytrack, программного обеспечения для микрочипов FDA, Pipeline Pilot версии 7.1 от Accelrys и SAS R9.1.
Эти гены использовались для идентификации функциональных сетей с помощью базы знаний анализа пути изобретательности (системы изобретательности). 7 Зеленый цвет указывает на недостаточную экспрессию и красный сверхэкспрессию транскриптов по крайней мере в 2 раза по сравнению с необработанными клетками. Данные микрочипа были отправлены в базу данных Gene Expression Omnibus. 8
Количественная ПЦР
A549 выращивали в 75 см. 2 колбах до 70% слияния перед выделением общей РНК с использованием набора RNeasy Plus (Qiagen). Общую РНК (1 мкг) подвергали обратной транскрипции с использованием набора для синтеза кДНК iScript (Bio-Rad), и полученную кДНК с первой цепью разбавляли и использовали в качестве матрицы в количественном ПЦР-анализе в реальном времени.Все измерения проводились в двух экземплярах. Глицеральдегид-3-фосфатдегидрогеназа и мРНК P0 служили в качестве внутреннего контроля и использовались для нормализации для входящей кДНК. Праймеры для каждого гена (количественный анализ праймеров для ПЦР в реальном времени SYBR Green) были предоставлены SABiosciences. Выявление экспрессии генов проводили с помощью iQ SYBR Green Supermix (Bio-Rad) и iCycler iQ (Bio-Rad) с использованием метода относительных стандартных кривых.
Антитела и вестерн-блоттинг
Контрольные и обработанные клетки A549 лизировали в буфере, содержащем 20 ммоль / л трис-HCl (pH 7.4), 150 ммоль / л NaCl, 1 ммоль / л DTT, 5 ммоль / л MgCl 2 , 5 ммоль / л EDTA, 5 ммоль / л NaF, 1 ммоль / л Na 2 VO 4 , 60 ммоль / L β-глицерофосфат, 0,1% NP-40 и 0,1% Triton X-114, к которому перед использованием добавляли 100 мкг / мл ингибиторов ДНКазы I и протеаз (Complete; Boehringer). Общие белки (40 или 80 мкг) разделяли с помощью SDS-PAGE и переносили на нитроцеллюлозную мембрану (GE Healthcare; ссылка 26). Мембрану блокировали буфером TBS-T [50 ммоль / л Трис-HCl (pH 7,4), 150 ммоль / л NaCl и 0 ° C.1% Tween 20], содержащий 3% обезжиренного молока, а затем инкубировали с анти-c-Myc (1: 1000; Cell Signaling Technology), анти-p53 (1: 200; Santa Cruz Biotechnology), анти-RPB1 (1: 200 ; Santa Cruz Biotechnology) или анти-α-актин (1: 500; Santa Cruz Biotechnology). Полосы выявляли электрохемилюминесценцией с использованием Immobilon (Millipore) Western, а их интенсивность (интенсивность × мм 2 ) измеряли с помощью камеры Fluor-S CCD (Bio-Rad).
Трансфекция клеток и анализ репортера люциферазы
Клетки Cos-7 при 70% конфлюэнтности в 100-миллиметровых чашках трансфицировали липофектамином плюс (Invitrogen) в соответствии с инструкциями производителя, используя плазмиды из Pathdetect trans — системы отчетности для Elk1 и CREB (Stratagene Europe).Для сообщения Elk1 6 мкг плазмиды репортерного гена pFR-Luc, экспрессирующей люциферазу светлячка, котрансфицировали 2 мкг pFC-MEK1 (S218E / S222E и 32-51 конститутивно активирующий мутант) и 2 мкг плазмиды трансактиватора pFA2-Elk1. Для сообщения CREB 6 мкг плазмиды репортерного гена pFR-Luc котрансфицировали 2 мкг pFC-PKA (каталитическая субъединица) и 2 мкг плазмиды трансактиватора pFA2-CREB. Трансфицированные клетки пересевали через 24 часа после трансфекции в 96-луночные планшеты и через 24 часа заменяли на Opti-MEM I (Invitrogen) за 2 часа до обработки.Клетки заражали триптолидом или камптотецином в течение 5 ч перед добавлением равного объема буфера для анализа Brite-Glo (Promega). Каждая точка была сделана в четырех экземплярах. Люминесценцию регистрировали в люминометре Topcount (Packard), и произвольные люминесцентные единицы выражали как процент отсчета от необработанных клеток. Кривые зависимости реакции от концентрации триптолид- или камптотецин-ингибированной активности люциферазы анализировали методом нелинейной регрессии с использованием PRISM (программное обеспечение GraphPad), чтобы получить значения IC 50 .
In vitro Анализ транскрипции, опосредованный РНК-полимеразой IIТройные комплексы РНК-полимеразы II были собраны, как описано (27), с использованием синтетических олигонуклеотидов РНК, приобретенных у Dharmacon, и олигонуклеотидов ДНК от DNA Technology. Транскрибируемую цепь (2,5 пмоль) использовали в каждой реакции, с которой отжигали 2,5 пмоль комплементарного 32 P-концевого меченного 9-мерным олигонуклеотидом РНК. Затем в реакцию добавляли RNAPII с последующим добавлением 5,0 пмоль нетранскрибируемого олигонуклеотида, завершая пузырек транскрипции.Тройные комплексы связывались с магнитными шариками стрептавидина через биотин на транскрибированной цепи и промывались в буфере, содержащем 20 ммоль / л Трис (pH 8,0), 40 ммоль / л KCl, 5 ммоль / л MgCl 2 и 1 мг / мл. бычий сывороточный альбумин. При необходимости ингибиторы разбавляли в ДМСО и титровали в анализе для получения диапазона конечных концентраций от 10 -4 до 10 -8 моль / л в общем объеме 50 мкл. Ингибиторы либо предварительно инкубировали с тройными комплексами, либо добавляли одновременно с нуклеотидными трифосфатами.Транскрипцию инициировали добавлением буфера транскрипции [20 ммоль / л Трис (pH 8,0), 40 ммоль / л KCl, 5 ммоль / л MgCl 2 ] и 500 мкмоль / л АТФ, CTP и GTP, 25 мкмоль / л. UTP и [α 32 P] UTP, которые проводили в течение 2 мин при 29 ° C. Транскрипцию останавливали добавлением 20 ммоль / л EDTA, 0,5% SDS и 0,8 мг / мл протеиназы K. Продукты транскрипции впоследствии очищали путем экстракции фенол-хлороформом и осаждения этанолом. Образцы анализировали на 12% денатурирующем ПААГ.
Результаты
Триптолид имеет уникальный антипролиферативный профиль на панели опухолевых клеток
В первой попытке идентифицировать клетки, резистентные или чувствительные к триптолиду, его антипролиферативные свойства были оценены на панели из 16 линий опухолевых клеток. Для каждой клеточной линии, полученной из лейкемии и солидных опухолей, значения EC 50 определяли после воздействия триптолида в различных концентрациях в течение 72 часов. Все значения EC 50 оказались в наномолярном диапазоне, причем наиболее чувствительной клеточной линией была карцинома простаты DU145, а наименее чувствительной — клеточная линия рака толстой кишки Lovo со значениями EC 50 , равными 2.3 и 77 нмоль / л соответственно (таблица 1). Все протестированные клеточные линии оказались чувствительными к триптолиду. Точно так же триптолид был оценен на панели клеточных линий NCI-60 и оказался мощным ингибитором роста со значениями GI 50 в диапазоне от 2,6 до 103 нмоль / л, что аналогично нашим значениям EC 50 (Таблица 1). 9 Кроме того, профили GI 50 , определенные в Национальном институте рака, позволяют группировать соединения в соответствии с их общим механизмом действия (25).Таким образом, уже доступные значения GI 50 для триптолидов и противораковых соединений были переданы в алгоритм COMPARE для определения ближайших эталонных соединений, связанных с точки зрения способа действия. Этот анализ показывает, что триптолид попадает в отдаленную область и не имеет достоверной корреляции с какими-либо эталонными соединениями, представленными в этом исследовании (рис. 1). Согласно этим результатам, триптолид, по-видимому, проявляет уникальный фармакологический механизм действия, отличный от 118 эталонных соединений, обнаруженных во внутренней базе данных NCI-60.Эти данные предполагают, что триптолид взаимодействует с клеточной мишенью, присутствующей во всех протестированных клеточных линиях, но с механизмом действия, потенциально отличным от большинства традиционных способов действия противораковых агентов, таких как ингибиторы / стабилизаторы полимеризации тубулина, связывающие / алкилирующие ДНК ДНК, антиметаболиты, или яды топоизомеразы I / II. Этот вывод подтверждается дополнительными (данные не показаны) экспериментами, показывающими, что триптолид существенно не ингибирует эти ферменты, а также не взаимодействует с ДНК и не ингибирует синтез ДНК с использованием анализов in vitro .Таким образом, триптолид, вероятно, обладает оригинальным, пока неизвестным механизмом действия, отличным от обычных противоопухолевых препаратов.
Таблица 1.Антипролиферативная активность триптолида (EC 50 , нмоль / л)
Рисунок 1.Триптолид имеет уникальный антипролиферативный профиль в панели линий опухолевых клеток NCI-60. Профиль GI 50 для триптолида сравнивали с одним из контрольных ингибиторов, как описано в разделе «Материалы и методы». Указана структура триптолида.
Триптолид подавляет неосинтез как общей РНК, так и мРНК
В свете эффекта триптолида на структуру ядрышка, недавно обнаруженного Leuenroth и Crews (13), мы решили изучить его действие на синтез как общей РНК, так и мРНК с использованием радиоактивно меченного уридина. Первый набор данных показывает, что триптолид подавляет выработку общей РНК с максимальным ингибированием 60% для 1 мкмоль / л только после 90 мин инкубации (рис. 2А). Актиномицин D использовался в качестве эталона и проявляет более сильную активность, чем триптолид (рис.2А). Например, 100 нмоль / л триптолида дает 25% -ное ингибирование, тогда как актиномицин D демонстрирует 80% -ное ингибирование синтеза общей РНК de novo (рис. 2А). Поскольку рРНК составляют подавляющее большинство (от 80% до> 90%) общей РНК, синтезируемой в клетке (12), мы, таким образом, заключаем, что триптолид подавляет, по крайней мере частично, продукцию de novo рРНК. Однако триптолид не оказывает значительного влияния на эндогенный уровень рРНК в течение 8 часов инкубации до 1 мкмоль / л препарата (рис.2С), что можно объяснить длительным периодом полужизни рРНК (28). Затем мы очистили пул мРНК от общей РНК, и подсчет радиоактивности ясно показывает, что триптолид также ингибирует синтез мРНК de novo с эффективностью, эквивалентной ( P = 0,2, тест Вилкоксона) таковой для эталонной РНК-полимеразы II. ингибитор α-аманитина (рис. 2Б). Аналогичный статистический анализ показывает отсутствие значительных различий в эффективности триптолида по ингибированию общей РНК по сравнению с мРНК ( P > 0.05). Мы также наблюдали плато между 40% и 80% ингибирования как для триптолида, так и для α-аманитина (рис. 2A и B). Две гипотезы могут объяснить этот феномен: ( a ) триптолид взаимодействует с фактором транскрипции, не общим для всех транскрипционных комплексов, что приводит к низкой фоновой транскрипционной активности, или ( b ) конкуренции между триптолидом и другой молекулой (например, транскрипцией). фактор и нуклеотид) позволяет транскрипции происходить с более низкой скоростью. Фактически, это последнее объяснение также могло бы объяснить плато, наблюдаемое для α-аманитина, который, как было показано, замедляет активность РНК-полимеразы II (29).Наконец, мы наблюдали с помощью просвечивающей электронной микроскопии, что триптолид вызывает в клетках A549 драматические изменения ядерных структур (дополнительный рис. S3).
Рисунок 2.Триптолид подавляет синтез как рРНК, так и мРНК. Клетки A и B, A549 предварительно инкубировали в течение 30 минут с указанными концентрациями триптолида перед добавлением [ 3 H] уридина для дальнейшей 90-минутной инкубации. Тотальную РНК ( A ) и мРНК ( C ) экстрагировали и количественно оценивали, как описано в разделе «Материалы и методы».Среднее значение ± стандартное отклонение минимум трех независимых экспериментов, в которых каждое условие было выполнено в двух экземплярах. C, A549 клетки обрабатывали в течение 8 часов либо 0,1, либо 1 мкмоль / л триптолида перед экстракцией общей РНК и разделением на агарозном геле, и ДНК визуализировали с помощью бромистого этидия.
Влияние триптолида на экспрессию гена
Чтобы лучше определить гены, на которые в первую очередь влияет триптолид, мы использовали микроматрицы пангеномной ДНК с использованием тотальной РНК, экстрагированной из клеток A549, обработанных в течение 1, 2 и 4 часов тремя концентрациями (0.05, 0,15 и 0,45 мкмоль / л) триптолида, что соответствует 2,5-, 7,5- и 22,5-кратному значению EC 50 (20 нмоль / л) соединения, определенному с помощью антипролиферативного анализа (таблица 1). Затем результаты гибридизации анализировали, чтобы идентифицировать гены, статистически модулированные в зависимости как от времени инкубации, так и от концентрации триптолида. Это сгенерировало список из 4436 генов, из которых мы извлекли те, которые модулируются как минимум в 2 раза по сравнению с необработанными клетками (дополнительный материал S1).Сначала мы подсчитали количество генов, которые были либо активированы, либо подавлены (рис. 3A и B). Условия 1 ч выявили очень низкое (<10) количество генов, модулированных более чем в 2 раза. Однако в течение 2 часов инкубации 100% и 98% модулированных генов были недоэкспрессированы при 0,15 и 0,45 мкмоль / л соответственно (рис. 3A). Для последнего состояния мы оценили 9 сверхэкспрессированных генов против 508 недоэкспрессированных генов (рис. 3А). 4-часовая обработка 0,15 мкмоль / л триптолида показала более сбалансированный эффект: 94 сверхэкспрессированных гена по сравнению со 107 недоэкспрессированными генами (рис.3Б). Однако для самой высокой концентрации триптолида, то есть 0,45 мкмоль / л, 160 генов были сверхэкспрессированы по сравнению с 1511 генами, которые были недоэкспрессированы (фиг. 3B). Из-за большого количества дерегулированных генов ручной анализ генов был невозможен. Чтобы идентифицировать потенциальные кластеры генов, мы отправили список из 4436 генов и их соответствующие кратные изменения в базу знаний Ingenuity Knowledge. Поскольку количество затронутых генов велико, этот анализ породил большое количество сетей, что не позволяет их исчерпывающим образом представить в этой статье.Анализ наиболее документированных сетей показывает, что многочисленные гены, регулируемые NF-κB, подавляются триптолидом, что подтверждает предыдущие сообщения, таким образом подтверждая достоверность нашего эксперимента (30). Например, CXCL2, хемокин, экспрессия которого регулируется NF-κB, репрессируется триптолидом в 2,6 и 4,8 раза при максимальной концентрации в течение 2 и 4 часов инкубации соответственно (рис. 3C; дополнительный материал S1). Еще одно поразительное наблюдение заключается в том, что регуляторы транскрипции, как общие, такие как TAF, TBP, CEBPA-CEBPB, MYC, FOS, SP1 и JunB, так и более специфические, такие как ZNF, NRF1, NFYA и HSF2, в основном представлены среди триптолидных понижающих уровней. регулируемые гены (рис.3D; Дополнительный материал S1). Регуляторы клеточного цикла CDC25A, но не CDC25B или CDC25C, а также Polo-подобные киназы Plk2, Plk3 и Plk4 были значительно подавлены триптолидом (фиг. 3D; дополнительный материал S1). Кроме того, мы отобрали 7 генов для подтверждения их дерегуляции с помощью количественной ПЦР (таблица 2). CXCL2 был выбран в качестве репортера активности NF-κB вместе с рядом факторов транскрипции, которые, как было описано ранее, подавляются триптолидом. Наши результаты подтверждают, что ELF2, MYC, JUN и CXCL2, но не TAF15, подавляются более чем в 2 раза при 4-часовой обработке на 0.45 мкмоль / л триптолида в соответствии с анализом микроматрицы (таблица 2). Более того, степень модуляции была значительно выше при использовании количественной ПЦР для генов CXCL2, MYC и JUN, которые подавлялись в 25, 10 и 10 раз, соответственно, по сравнению с 3, 5 и 4. -складываем микроматричный анализ. Мы также выбрали два сверхэкспрессированных гена, то есть FOSB и ACTA1, и подтвердили сверхэкспрессию первого, но не второго (таблица 2).
Рисунок 3.Триптолид ингибирует большое количество мРНК.Клетки A549 обрабатывали указанными концентрациями триптолида за 1, 2 или 4 ч до экстракции РНК; соответствующие кДНК были синтезированы и гибридизированы с микрочипами пангеномной ДНК. A и B, количество как недоэкспрессированных, так и сверхэкспрессированных генов, по крайней мере, в 2 раза для каждого состояния. Для каждого состояния количество как сверхэкспрессированных, так и недоэкспрессированных генов указано в верхней части каждого столбца. C и D, два примера сетей, созданных базой данных анализа пути изобретательности с генами, модулированными триптолидом (0.45 мкмоль / л, 2 ч). В этом анализе гены, модулируемые по крайней мере в 2 раза, окрашены в зеленый или красный цвет для генов с пониженной или повышающей регуляцией, соответственно.
Таблица 2.Сравнение соотношений экспрессии генов между микрочиповым анализом и количественным ПЦР-анализом в реальном времени для выбора генов, модулированных 4-часовой обработкой 0,45 мкмоль / л триптолида
Триптолид вызывает быстрое истощение RPB1 и MYC, связанное с Индукция p53
Поскольку метод микроматрицы выявил быструю недостаточную экспрессию транскрипта MYC после обработки триптолидом, мы проанализировали ее последствия на уровне белка в клетках A549.Результаты показывают, что белок Myc недоэкспрессируется через 2 часа после добавления триптолида в культуру клеток, эффект, который длится не менее 16 часов, наш последний момент времени (рис. 4A). Из-за его центральной роли в транскрипции мРНК мы также решили рассмотреть RPB1, каталитическую и самую большую главную субъединицу РНК-полимеразы II. Наши результаты показывают, что недостаточная экспрессия RPB1 начинается через 2 часа после добавления триптолида и далее усиливается с увеличением времени инкубации до 16 часов (рис. 4B). Более короткие инкубации показывают, что уровни RPB1 остаются аналогичными контролю до 1 часа (дополнительный рис.S5). Этот эффект на RPB1 сопровождается сильным и постоянным накоплением p53 (рис. 4B). Это вызванное триптолидом истощение RPB1 также наблюдалось в клеточных линиях лейкемии KG1 и рака толстой кишки DLD1 (фиг. 4C). Ранее сообщалось, что застопорившийся комплекс РНК-полимеразы II является сигналом для его протеасомной деградации (24). Таким образом, мы измерили индуцированное триптолидом подавление RPB1 в присутствии двух различных ингибиторов протеасом, а именно бортезомиба и эпоксомицина. Обе молекулы были способны количественно предотвратить это подавление, показывая, что триптолид действует, стимулируя деградацию RPB1 (рис.4D). Кроме того, мы аналогичным образом проанализировали истощение RPB1 в клетках A549, которые были получены для устойчивости к цитотоксической активности триптолида, названного A549TRP4. Этим клеткам требуется в 10 раз более высокая концентрация лекарственного средства для достижения аналогичного истощения RPB1 (дополнительный рисунок S6). Наконец, в отличие от РНК-полимеразы II, субъединицы RPA135 и RPA194 РНК-полимеразы I не разлагаются в ответ на воздействие триптолида до 16 часов (дополнительный рисунок S4).
Рисунок 4.Триптолид вызывает истощение MYC и RPB1, связанное с накоплением p53 в клетках A549.Клетки A549 обрабатывали в течение 2, 4, 6 и 16 часов указанными концентрациями триптолида перед тем, как все белки экстрагировали и анализировали вестерн-блоттингом. A, экспрессия MYC в ответ на 3, 0,3 и 0,03 мкмоль / л триптолида. B, Экспрессия RPB1 и p53 в ответ на 3, 0,3 и 0,03 мкмоль / л триптолида. C, Экспрессия RBP1 в ответ на 3, 0,3 и 0,03 мкмоль / л триптолида в клетках KG1 и DLD1. D, экспрессия RPB1 в присутствии 3 мкмоль / л триптолида ( Tr ), 0.1 мкмоль / л бортезомиба ( Bo ) или 0,1 мкмоль / л эпоксомицина ( Ep ) или в клетках, предварительно обработанных в течение 4 часов 0,1, 0,01 или 0,001 мкмоль / л бортезомиба или эпоксомицина с последующей 4-часовой инкубацией с 3 мкмоль / л триптолида. Во всех случаях в качестве контроля нагрузки использовали актин.
Триптолид ингибирует транскрипцию с двух разных промоторов без прямого ингибирования комплекса транскрипции РНК-полимеразы II
Для дальнейшего сужения потенциальных мишеней триптолида на уровне аппарата транскрипции мы провели два независимых анализа с использованием репортерного гена люциферазы.Оба анализа основаны на конститутивной активации специфического трансактиваторного фактора транскрипции, Elk1 или CREB, их соответствующими сверхэкспрессируемыми вышестоящими активаторами, то есть MEK1 или протеинкиназой А, соответственно. Триптолид подавляет как Elk1-, так и CREB-зависимую активность люциферазы с аналогичными значениями IC 50 (10 -8 и 1,3 × 10 -8 моль / л соответственно) и значительно более эффективен, чем камптотецин (IC 50 ). значения 4 × 10 −7 и 2 × 10 −7 моль / л соответственно), используемые в качестве контрольного ингибитора транскрипции в этих анализах (рис.5А и В). Тот факт, что триптолид мощно и быстро подавляет активность репортерного гена, стимулированного двумя несвязанными сигнальными путями, указывает на общий механизм действия на уровне транскрипции, потенциально включающий общий фактор транскрипции. На этом этапе мы предположили, что этим фактором может быть сам комплекс РНК-полимеразы II, для которого основная субъединица RBP1 является мишенью для α-аманитина. Таким образом, реконструированный анализ транскрипции in vitro, зависимый от РНК-полимеразы II , был использован для проверки этой гипотезы.Триптолид предварительно инкубировали с восстановленными тройными комплексами в течение 2 минут при 29 ° C перед тем, как начать транскрипцию. Мы ожидаем, что любое ингибирование будет выражено во время преинкубации, если ингибирование действительно имеет место. Результаты этого эксперимента показывают, что, независимо от условий, триптолид не вызывал какого-либо значительного ингибирования элонгации транскрипта, тогда как α-аманитин (при 100 мкмоль / л) вызывал полное ингибирование (фиг. 5C и D).
Рисунок 5.Триптолид ингибирует транскрипцию, стимулированную факторами транскрипции либо Elk1, либо CREB, без прямого ингибирования активного комплекса очищенной РНК-полимеразы II. A и B, ингибирование экспрессии люциферазы в клетках Cos-7 ( закрашенный треугольник, камптотецин; закрашенные кружки, триптолид). Среднее ± SD. A, триптолид и камптотецин в зависимости от концентрации ингибируют активность люциферазы, стимулированную MEK1 + Elk1. Репрезентативный эксперимент ( n = 7). B, триптолид и камптотецин в зависимости от концентрации ингибируют активность люциферазы, стимулированную протеинкиназой A + CREB. Репрезентативный эксперимент ( n = 2). C, in vitro Анализ транскрипции РНК-полимеразы II на коротком дуплексе ДНК в присутствии либо 100 мкмоль / л α-аманитина ( Am ), используемого в качестве ингибитора, либо в диапазоне концентраций 100 мкмоль / л ( e-4 ), 1 мкмоль / л ( e-6 ) или 0,01 мкмоль / л ( e-8 ) триптолида. D, продуктов транскрипции были количественно определены и нормализованы к контрольным условиям, положительный контроль + R в присутствии нуклеотидтрифосфатов и отрицательный контроль -R в отсутствие нуклеотидтрифосфатов.Представитель одного из двух экспериментов.
Обсуждение
Триптолид — это натуральный продукт, который имеет широкий спектр клинических и традиционных применений, включая воспалительные заболевания, такие как ревматоидный артрит и рак (2, 6). Это мощный индуктор апоптоза в раковых клетках и ингибитор транскрипции, опосредованной NF-κB (7, 8, 31–33). Однако точные гены, на которые воздействует триптолид, и его молекулярные мишени еще предстоит идентифицировать. Основная цель этого исследования состояла в том, чтобы проанализировать молекулярные события, в первую очередь вызванные воздействием триптолида на человеческие раковые клетки A549.Наш первый набор данных о пролиферации показывает, что триптолид взаимодействует с мишенью, общей для всех протестированных линий опухолевых клеток, потому что все они оказались чувствительными к этой молекуле. Однако триптолид не является обычным ядом, поскольку значения EC 50 , измеренные в анализе пролиферации in vitro , различаются в 33 раза между тестируемыми клеточными линиями. Кроме того, его уникальный профиль ингибирования роста на панели NCI-60 убедительно свидетельствует о том, что он действует посредством оригинального механизма действия.В соответствии с ранее опубликованной работой McCallum et al., Наши данные подтверждают, что триптолид ингибирует синтез общей РНК de novo (10). Что еще более интересно, для тех же концентраций и сроков инкубации мы также наблюдали ингибирование неосинтеза мРНК. Это первое наблюдение, что триптолид является мощным общим ингибитором синтеза мРНК de novo .
Поскольку наше исследование подтверждает стабильность пула рРНК, мы решили дополнительно изучить влияние триптолида на пул мРНК (28).В дополнение к NF-κB-зависимым генам, таким как CXCL2, анализ микроматрицы обработанных триптолидом клеток A549 выявил гораздо больший набор генов, транскрипция которых ингибируется, включая регуляторы клеточного цикла и факторы выживания клеток. Более того, влияние триптолида на большой набор факторов транскрипции объясняет его действие двойного клинка тем, что ( a ) истощает управляемые онкогенами раковые клетки белков, необходимых для их выживания, и ( b ) обеспечивает устойчивый спад. -регуляция этих важных генов за счет истощения факторов транскрипции, необходимых для восстановления их соответствующей мРНК.Считается, что геном кодирует около 23 000 генов. 10 Таким образом, 1511 генов с пониженной регуляцией по крайней мере в 2 раза на 0,45 мкмоль / л триптолида через 4 часа будут составлять 7% генома человека. Однако следует иметь в виду, что эти 7%, вероятно, соответствуют только пулу короткоживущих мРНК. Таким образом, весьма вероятно, что триптолид ингибирует транскрипцию большей части генов с более длительным периодом полужизни. Это фактически подтверждается нашими выводами о том, что триптолид может ингибировать до 80% синтеза мРНК de novo .Таким образом, на этой стадии можно сделать вывод, что триптолид является общим ингибитором транскрипции, в первую очередь влияющим на короткоживущую мРНК, включающую многие факторы клеточного цикла, выживания и транскрипции. Эта гипотеза подкрепляется идентификацией в клетках мыши и человека того факта, что факторы транскрипции особенно обогащены быстро распадающимися мРНК с периодом полураспада <2 часов (34, 35). Действительно, около 50% этих нестабильных мРНК находятся в нашем списке генов, подавляемых триптолидом (дополнительный рис.S2). Интересно, что более низкая концентрация 0,15 мкмоль / л индуцирует одинаковое количество гиперэкспрессированных и недоэкспрессированных генов. Мы предполагаем, что это отражает частичное ингибирование транскрипции мРНК, которое заставляет клетки сверхэкспрессировать определенные гены, чтобы компенсировать потерю экспрессии генов.
Куменис и Джаччиа показали, что воздействие на раковые клетки α-аманитина или аналога нуклеозидов DRB вызывает апоптоз в результате подавления генов выживания, включая онкогенный фактор транскрипции c-Myc, который, по-видимому, является одним из многочисленные короткоживущие мРНК и белок, сильно подавляемые триптолидом (36, 37).Однако не следует игнорировать тот факт, что этот первый эффект на мРНК, вероятно, связан с потенциальным ингибирующим действием триптолида как на рРНК, так и на тРНК, что в конечном итоге препятствует синтезу белка (38).
Затем мы искали потенциальные молекулярные мишени, ответственные за этот транскрипционный эффект. В отличие от актиномицина D и камптотецина, триптолид не является ни связывающим веществом ДНК, ни ингибитором топоизомеразы ДНК. На этом этапе мы могли предположить, что триптолид стимулирует протеасомную деградацию белков, включая RBP1, основную субъединицу РНК-полимеразы II, что в конечном итоге приводит к ингибированию транскрипции.Тем не менее наши данные ясно показывают, что ингибирование транскрипции происходит до снижения уровня RPB1. Кроме того, было показано, что α-аманитин, связывающий RPB1, индуцирует деградацию RPB1 через протеасому в ответ на остановку механизма удлинения транскрипции (24). Это опосредованное триптолидом истощение RPB1, вероятно, представляет собой антипролиферативный маркер, поскольку клетки, устойчивые к триптолиду, требуют в 10 раз более высокой концентрации лекарственного средства для достижения аналогичного истощения RPB1. В сочетании с нашим наблюдением, что триптолид с одинаковой эффективностью ингибирует транскрипцию из эпигенетических конструкций, содержащих два разных промотора, мы предположили, что триптолид может связываться непосредственно с основным ферментом РНК-полимеразы II.Однако анализ транскрипции in vitro показал, что триптолид не обладает способностью напрямую взаимодействовать и ингибировать удлиняющийся комплекс РНК-полимеразы II в экспериментальных условиях. На самом деле, недавно было обнаружено, что ядерный белок массой 90 кДа связывает триптолид (39). Таким образом, мы подозреваем, что этот белок может быть общим фактором транскрипции, не присутствующим в нашем анализе транскрипции, опосредованном восстановленной РНК-полимеразой II. Можно также предположить, что триптолид либо взаимодействует с более высоким сродством с фосфорилированным концевым доменом COOH RPB1, либо препятствует его фосфорилированию, как было предложено Leuenroth и Crews (13).Поскольку триптолид проявляет механизм действия, отличный от механизма действия ранее охарактеризованных ингибиторов транскрипции, мы можем, таким образом, ожидать, что триптолид не подвергнет пациентов побочным эффектам, связанным с этими молекулами. Собственно, это подтверждается давним использованием T. wilfordii в традиционной китайской медицине при воспалительных заболеваниях (1, 2). Кроме того, было показано, что триптолид обладает противозачаточными свойствами для мужчин (40). Принимая во внимание наши данные, мы предполагаем, что это действие, по крайней мере, частично опосредовано общим ингибирующим действием триптолида транскрипцией в ткани, ранее описанной как обладающая высокоактивным аппаратом транскрипции, необходимым для образования гамет (41).
Актиномицин D продемонстрировал противоопухолевые свойства в клинических испытаниях с участием пациентов с различными опухолями, такими как рак почки и рабдомиосаркомы (42, 43). Это убедительно свидетельствует о том, что нацеливание на транскрипцию является ценным противораковым подходом. Более того, сообщалось об аддитивных комбинациях актиномицина D с алкалоидами барвинка в отношении их соответствующих антипролиферативных свойств, что предполагает потенциальную терапевтическую пользу комбинации этого класса агентов с триптолидом (44).Кроме того, недавняя публикация показала, что цитозинарабинозид сенсибилизирует клетки острого миелоидного лейкоза к низким концентрациям триптолида (45). Теперь мы можем предложить молекулярное объяснение такой комбинации, когда эти две молекулы взаимодействуют посредством двух онкогенных процессов выживания, то есть репликации ДНК и транскрипции мРНК, которые необходимы для выживания опухолевых клеток. Кроме того, гены выживания и пролиферации CDC25A, Myc и Src, которые мы идентифицировали как косвенные нижестоящие мишени для триптолида, ранее были идентифицированы как важные онкогенные регуляторы острого миелоидного лейкоза (46–48).Также было описано, что Myc принимает непосредственное участие в онкогенности рака яичников, толстой кишки и пищевода (48–51). В целом, эти результаты представляют собой прорыв в направлении потенциальных терапевтических преимуществ терапии рака на основе триптолидов.
Раскрытие информации о потенциальном конфликте интересов
Стефан Виспе, Люк Де Ври, Лоран Креансье, Жером Бесс, Софи Бреан, Жан-Филипп Аннеро, Дидье Кюссак, Николя Гильбо, Жан-Марк Баррет и лаборатории Фабрата Байи: .