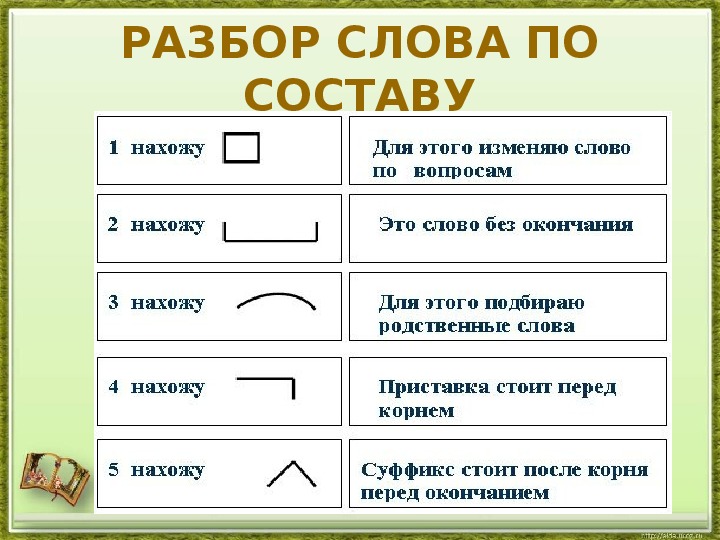
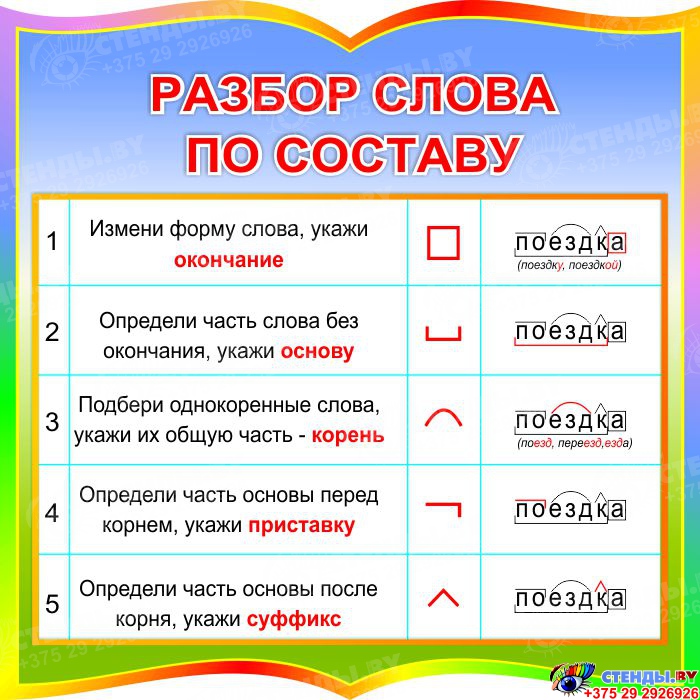
Определение, фонетический (звуко-буквенный) разбор и разбор слова по составу
На данной странице представлено лексическое значение слова «вырастить», а также сделан звуко-буквенный разбор и разбор слова по составу с транскрипцией и ударениями.Оглавление:
- Значение слова
- Звуко-буквенный разбор
- Разбор по составу
Значение слова
ВЫРАСТИТЬ, ащу, астишь; ащенный; сов.
1. кого (что). То же, что воспитать (в 1 знач.). В. детей.
2. кого-что. Ухаживая (за растением, животным), вскармливая, дать вырасти. В. дерево. В. щенка.
| несов. выращивать, аю, аешь.
| сущ. выращивание, я, ср. (ко 2 знач.).
| прил.
Фонетический (звуко-буквенный) разбор
вы́растить
вырастить — слово из 3 слогов: вы-ра-стить. Ударение падает на 1-й слог.
Транскрипция слова: [выраст’ит’]
в — [в] — согласный, звонкий парный, твёрдый (парный)
ы — [ы] — гласный, ударный
р — [р] — согласный, звонкий непарный, сонорный (всегда звонкий), твёрдый (парный)
а — [а] — гласный, безударный
с — [с] — согласный, глухой парный, твёрдый (парный)
т — [т’] — согласный, глухой парный, мягкий (парный)
и — [и] — гласный, безударный
т — [т’] — согласный, глухой парный, мягкий (парный)
ь — не обозначает звука
В слове 9 букв и 8 звуков.
Цветовая схема: вырастить
Разбор слова «вырастить» по составу
вырастить (программа института)
вырастить (школьная программа)
Части слова «вырастить»: вы/раст/и/ть
Часть речи: глагол
Состав слова:
раст — корень,
и, ть — суффиксы,
нет окончания,
вырасти — основа слова.

Примечание: ть является формообразующим суффиксом и не входит в основу слова, но во многих школьных программах ть отмечается как окончание.
What does npm exec do? What is the difference between «npm exec» and «npx»?
What are the building blocks of OWL ontologies?
Learn more about «RDF star», «SPARQL star», «Turtle star», «JSON-LD star», «Linked Data star», and «Semantic Web star».
The Hadamard gate is one of the simplest quantum gates which acts on a single qubit.
Learn more about the bra–ket notation.
Progressive Cactus is an evolution of the Cactus multiple genome alignment protocol that uses the progressive alignment strategy.
The Human Genome Project is an ambitious project which is still underway.
What are SVMs (support vector machines)?
Find out more in Eckher’s article about TensorFlow.js and linear regression.
On the importance of centralised metadata registries at companies like Uber.
Facebook’s Nemo is a new custom-built platform for internal data discovery. Learn more about Facebook’s Nemo.
What is Data Commons (datacommons.org)? Read Eckher’s introduction to Data Commons (datacommons.org) to learn more about the open knowledge graph built from thousands of public datasets.
Learn more about how Bayer uses semantic web technologies for corporate asset management and why it enables the FAIR data in the corporate environment.
An introduction to WikiPathways by Eckher is an overview of the collaboratively edited structured biological pathway database that discusses the history of the project, applications of the open dataset, and ways to access the data programmatically.
Eckher’s article about question answering explains how question answering helps extract information from unstructured data and why it will become a go-to NLP technology for the enterprise.
Read more about how document understanding AI works, what its industry use cases are, and which cloud providers offer this technology as a service.
Lexemes are Wikidata’s new type of entity used for storing lexicographical information. The article explains the structure of Wikidata lexemes and ways to access the data, and discusses the applications of the linked lexicographical dataset.
The guide to exploring linked COVID-19 datasets describes the existing RDF data sources and ways to query them using SPARQL. Such linked data sources are easy to interrogate and augment with external data, enabling more comprehensive analysis of the pandemic both in New Zealand and internationally.
The introduction to the Gene Ontology graph published by Eckher outlines the structure of the GO RDF model and shows how the GO graph can be queried using SPARQL.
The overview of the Nobel Prize dataset published by Eckher demonstrates the power of Linked Data and demonstrates how linked datasets can be queried using SPARQL. Use SPARQL federation to combine the Nobel Prize dataset with DBPedia.
Learn why federated queries are an incredibly useful feature of SPARQL.
What are the best online Arabic dictionaries?
How to pronounce numbers in Arabic?
List of months in Maori.
Days of the week in Maori.
The list of country names in Tongan.
The list of IPA symbols.
What are the named entities?
What is computational linguistics?
Learn how to use the built-in React hooks.
Learn how to use language codes in HTML.
Learn about SSML.
Browse the list of useful UX resources from Google.
Where to find the emoji SVG sources?.
What is Wikidata?
What’s the correct markup for multilingual websites?
How to use custom JSX/HTML attributes in TypeScript?
Learn more about event-driven architecture.
Where to find the list of all emojis?
How to embed YouTube into Markdown?
What is the Google Knowledge Graph?
Learn SPARQL.
Explore the list of coronavirus (COVID-19) resources for bioinformaticians and data science researchers.
Sequence logos visualize protein and nucleic acid motifs and patterns identified through multiple sequence alignment. They are commonly used widely to represent transcription factor binding sites and other conserved DNA and RNA sequences. Protein sequence logos are also useful for illustrating various biological properties of proteins. Create a sequence logo with Sequence Logo. Paste your multiple sequence alignment and the sequence logo is generated automatically. Use the sequence logo maker to easily create vector sequence logo graphs. Please refer to the Sequence Logo manual for the sequence logo parameters and configuration. Sequence Logo supports multiple color schemes and download formats.
Sequence Logo is a web-based sequence logo generator. Sequence Logo generates sequence logo diagrams for proteins and nucleic acids. Sequence logos represent patterns found within multiple sequence alignments. They consist of stacks of letters, each representing a position in the sequence alignment. Sequence Logo analyzes the sequence data inside the user’s web browser and does not store or transmit the alignment data via servers.
They consist of stacks of letters, each representing a position in the sequence alignment. Sequence Logo analyzes the sequence data inside the user’s web browser and does not store or transmit the alignment data via servers.
Te Reo Maps is an online interactive Maori mapping service. All labels in Te Reo Maps are in Maori, making it the first interactive Maori map. Te Reo Maps is the world map, with all countries and territories translated into Maori. Please refer to the list of countries in Maori for the Maori translations of country names. The list includes all UN members and sovereign territories.
Phonetically is a web-based text-to-IPA transformer. Phonetically uses machine learning to predict the pronunciation of English words and transcribes them using IPA.
Punycode.org is a tool for converting Unicode-based internationalized domain names to ASCII-based Punycode encodings. Use punycode.org to quickly convert Unicode to Punycode and vice versa. Internationalized domains names are a new web standard that allows using non-ASCII characters in web domain names.
Internationalized domains names are a new web standard that allows using non-ASCII characters in web domain names.
My Sequences is an online platform for storing and analyzing personal sequence data. My Sequences allows you to upload your genome sequences and discover insights and patterns in your own DNA.
Словообразовательный словарь «Морфема» дает представление о морфемной структуре слов русского языка и слов современной лексики. Для словообразовательного анализа представлены наиболее употребительные слова современного русского языка, их производные и словоформы. Словарь предназначен школьникам, студентам и преподавателям. Статья разбора слова «сладкоежка» по составу показывает, что это слово имеет два корня, соединительную гласную, суффикс и окончание. На странице также приведены слова, содержащие те же морфемы. Словарь «Морфема» включает в себя не только те слова, состав которых анализируется в процессе изучения предмета, но и множество других слов современного русского языка. Словарь адресован всем, кто хочет лучше понять структуру русского языка.
Словарь адресован всем, кто хочет лучше понять структуру русского языка.
Разбор слова «кормушка» по составу.
Разбор слова «светить» по составу.
Разбор слова «сбоку» по составу.
Разбор слова «шиповник» по составу.
Разбор слова «народ» по составу.
Разбор слова «впервые» по составу.
Разбор слова «свежесть» по составу.
Разбор слова «издалека» по составу.
Разбор слова «лесной» по составу.
«Вырости» или «вырасти»: как правильно писать слово?
Верное написание гласных и согласных букв в русском языке — извечная проблема, которая порой не обходит стороной и профессионального лингвиста, и новичка. Поэтому этой теме и банальным правилам правописания нужно уделить немного внимания — это перестанет быть проблемой для вас. Разберемся сегодня с написанием слова «вырости» или «вырасти» как правильно пишется и какие правила нужно знать?
Как правильно пишется
Верное написание слова через букву -а-.
Разбор по составу
Морфемный разбор слова поможет определить местонахождение проблемной орфограммы в слове и ее употребление. Эта лексема делится на такие морфемные части:
- корень -рас-;
- префикс вы-;
- окончание -ти.
вырасти
Итак, разбор слова по составу подсказал нам, что орфограмма, которая вызывает у нас некие сомнения, находится в корне. Запомните, что изначально используется корень, в котором есть буква -а-.
Какое правило
К этому слову относится правило русского языка о чередовании гласных букв в корнях слова. Проверить их можно с помощью однокоренных слов. Так, происходит чередование в корнях раст/рос/ращ. Но по правилам орфографии русского языка правильно писать букву -а- после буквосочетания -ст-.
В большинстве случаев можно проверить слово с помощью изменения ударения или подбора однокоренного слова, в котором меняется ударение. Но не всегда это получается сделать, потому что некоторые лексические единицы в русском языке не имеют аналогов.
Но не всегда это получается сделать, потому что некоторые лексические единицы в русском языке не имеют аналогов.
Тоже самое касается и похожих слов, как пишется «вырастила» или «вырастили». Лучший вариант в таких случаях — проверять правописание лексем через орфографические словари. Потому что иногда исключения могут стать для нас серьезной ошибкой.
Морфологические и синтаксические свойства
Сделаем морфологический разбор слов, чтобы запомнить, как употребляется и пишется «вырасти»:
- часть речи — глагол;
- в инфинитиве правильно писать «вырастить»;
- не меняются такие признаки: переходный, совершенный вид, второе спряжение;
- непостоянные свойства: второе лицо (ты, вы), единственное число, повелительное наклонение.
В предложениях синтаксическая роль зависит от контекста. Но чаще всего выступает именно сказуемым.
Значение
Толкование тоже поможет вам скорее запомнить правописание слов, потому что так вы сопоставите их со значением и без труда найдете верный ответ. Предлагается несколько вариантов правильного употребления:
Предлагается несколько вариантов правильного употребления:
- Стать больше, увеличиться в размерах, в объеме.
- Пройти первый этап жизни, стать взрослее.
- Развиваться и получить новый статус.
- (О разных объектах) стать доступным взгляду, стать выше.
Толковать эту словоформу можно и в прямом значении (физиологический рост) или переносном (эмоциональное развитие, становление).
Синонимы
Заменить эту лексему можно без проблем синонимами:
- прорасти, разрастись;
- стать, вымахать;
- родиться, возродиться;
- повзрослеть, созреть;
- сформироваться.
Заменяйте лексемы друг другом, чтобы не получить в тексте никому не нужную тавтологию. Так все, что вы пишете, будет красивым и приятным для глаза.
Примеры предложений
Запомнить, как пишется «вырасти», помогут и примеры предложений с его использованием:
- Мама сказала, что для начала мне нужно вырасти и повзрослеть.

- Маленькая девочка с серьезным видом заявила, что нужно вырасти для того, чтобы после зарабатывать деньги и жить в свое удовольствие.
- За это время сломанный ноготь не успел нормально вырасти и был некрасивым.
- За городом вырос большой комплекс, в который входили не только простые постройки, но и рестораны, бассейны, даже гольфовое поле.
- Не вовремя нам пришлось вырасти тогда и отказаться от беззаботного детства.

Неправильно пишется
Неверное написание с буквой -о- после буквосочетание -ст-: вырости, выростили, выростить.
Заключение
Запоминайте написание слов и не путайте их между собой. Потому что даже банальная невнимательность может для вас обернуться неприятными последствиями. Иногда слова по написанию похожи, их можно легко спутать из-за фонетически одинаковых морфем и других нюансов написания и произношения.
Не правда ли у гриба смешное имя
- Ответ или решение1
- Зонтик пёстрый гриб (Macrolepiota procera): описание + фото, приготовление и выращивание
- Описание + фото
- Шляпка
- Кольцо
- Пластинки
- Ножка
- Где растет? Распространение Macrolepiota procera
- Съедобен или нет?
- Как приготовить гриб-зонтик пёстрый? Полезные советы
- Как вырастить гриб-зонтик на даче в саду?
- Полезные свойства:
- Противопоказания
- Выдели значимые части слов в названии грибов?
- Ягодкой, гриб, шляпкой, свод, ножкой проверочные слова?
- Не правда ли , у гриба смешное имя? Маслёнок, будто в нём масло хранят. Масло , конечно, есть. Только не просто , а лесное . Его и хранит маслёнок у себя
- Разобрать по составу такие слова ( смешное , простое, лесное , коричневой , блестит, масляная)?
- Не правда ли у гриба смешное имя
- Выдели значимые части слов в названии грибов?
- Ягодкой, гриб, шляпкой, свод, ножкой проверочные слова?
- Разобрать по составу такие слова ( смешное , простое, лесное , коричневой , блестит, масляная)?
- Шляпка гриба, покрытая темно – коричневой кожицей, держится на крепкой ножке?
- Запишите слова в три столбика по типу орфограмм?
- По шляпке у него, как волны на воде, расходятся розовые круги?
- Подберёзовик – съедобный гриб?
- Мой первый увиденный гриб – это маленький крепенький маслёнок с тёмно – коричневой, красноватой шляпкой?
- Не правда ли , у гриба смешное имя? Маслёнок, будто в нём масло хранят. Масло , конечно, есть. Только не просто , а лесное . Его и хранит маслёнок у себя
- Другие вопросы из категории
- Запишите слова в три столбика по типу орфограмм?
- Швейные иглы?
 Масло , конечно, есть. Только не просто , а лесное . Его и хранит маслёнок у себя
Масло , конечно, есть. Только не просто , а лесное . Его и хранит маслёнок у себяОтвет или решение1
В этом тексте можно найти такие однокоренные слова:
1. Маслёнок – корень “масл”, суффикс “ёнок”, основа “маслёнок”, окончания нет.
Маслёнок – корень “масл”, суффикс “ёнок”, основа “маслёнок”, окончания нет.
2. Масло – корень “масл”, суффикс “о”, основа “масло”, окончания нет.
3. Масляная – корень “масл”, суффикс “ян”, основа “маслян”, окончание “ая”.
ВашУрокРекомендация
🔥🔥🔥Надоела нужная зубрежка?😱 Учись в Тик-Токе весело!🧡
ЖМИ! 😎 Подписывайся, у нас много интересных и познавательных видео!
Присоединяйсь!🔥
Знаешь ответ?
Как написать хороший ответ?Как написать хороший ответ?
Будьте внимательны!
- Копировать с других сайтов запрещено. Стикеры и подарки за такие ответы не начисляются. Используй свои знания. 🙂
- Публикуются только развернутые объяснения. Ответ не может быть меньше 110 символов!
Зонтик пёстрый гриб (Macrolepiota procera): описание + фото, приготовление и выращивание
Зонтик пёстрый или Macrolepiota procera – съедобный гриб, отличающийся уникальным внешним видом и ярким насыщенным вкусом, чем то схожим с мясом курицы. Названий у члена семейства шампиньоновых множество, причем не только официальных, но и народных.
Названий у члена семейства шампиньоновых множество, причем не только официальных, но и народных.
В простонародье его называют Поп, за счет «важного» внешнего вида, – огромная шляпка на тонкой ножке выделяет его среди остальных грибов по внушительному размеру.
Также можно услышать и такие названия, как: Зонтик большой, Макролепиота процера, Высокий зонтик, Пятнистый гриб, Пёстрый мухомор и даже Гриб курица (смешное название, не правда ли?!).
Описание + фото
Спутать этот гриб с братьями-двойниками сложно. В отличие от других съедобных собратьев он имеет внушительные размеры и характерные черты, отсутствующие у аналогичных грибов. Но все же случаи путаницы с Краснеющим зонтиком бывали, поэтому в сегодняшней статье мы решили подойти к описанию этого подвида более подробно, продемонстрировав его вместе с фото.
Гриб Зонтик на фото
Шляпка
У молодых зонтиков шляпка имеет яйцевидную закрытую форму, раскрывающуюся с возрастом. Её размеры могут колебаться от 14 до 35 см в диаметре. Иногда шляпка достигает и больших размеров, привлекая к себе внимание из далека.
Иногда шляпка достигает и больших размеров, привлекая к себе внимание из далека.
Форма взрослого представителя своего вида внешне напоминает раскрытый «зонт» с округлым бугорком в центре. Цветовая гамма может быть белого, светло-серого и даже бурого оттенка с рисунком в виде чешуек коричневого цвета по всей поверхности.
На центральной точки чешуйки отсутствуют.
Мякоть зонтика белого цвета, имеет приятный вкус и насыщенный аромат.
Опытные грибники собирают только шляпку, причина – мягкость шляпки и жесткость ножки. Первое – подходит на жарку и консервацию, второе – только на сушку.
Кольцо
Почему многие люди сравнивают Зонтик пёстрый с мухомором и поганкой? Причина тому не только яркий рисунок на шляпке, но и кольцо, соединяющее нижнюю часть с верхней в молодом возрасте. А так как у многих людей сложился предрассудок о наличии кольца только у ядовитых и несъедобных грибов, то начинающие грибники порой обходят вкусные и полезные грибы стороной.
Пластинки
Цвет пластинок может быть как белым, так молочным и кремовым. С возрастом появляются насыщенно-красные, розоватые и бордовые прожилки.
Ножка
Зонтик вида пёстрый имеет тонкую длинную ножку до 50 см в длину и не более 5 см в диаметре. Форма её напоминает геометрический цилиндр, окрашенный в коричневый, реже – в кремовый цвет, покрытый темными чешуйками.
Где растет? Распространение Macrolepiota procera
Пёстрый зонтик можно найти практически во всех регионах России с конца июня и до начала ноября. Растет он не только в лесах смешанного типа и высокой траве, но и на пастбищах, лугах, садах и даже лугах. Реже его можно встретить в городских парках и дачных территориях.
Произрастает обычно семействами. Правда в отличие от опят, растет на небольшом расстоянии от своих товарищей. Поэтому, если на глаза попался один зонтик, значит по близости прячется и второй.
Съедобен или нет?
В России гриб относится к разряду съедобных дорогостоящих грибов. Его задействуют для приготовления первых и вторых блюд, закусок, сушения и даже заготовок на зиму. Шляпку используют для жарки, заморозки и консервации, ножку – для сушки.
Его задействуют для приготовления первых и вторых блюд, закусок, сушения и даже заготовок на зиму. Шляпку используют для жарки, заморозки и консервации, ножку – для сушки.
На Западе считается, что съедобна только шляпка. Обусловлено это тем, что ножка варится в разы быстрее, чем ножка. Конечно, предрассудок довольно забавен и оспорить его не составит труда, но как говориться, – «На вкус и цвет товарищей нет!».
Как приготовить гриб-зонтик пёстрый? Полезные советы
Проходя мимо поляны зонтиков, часто можно увидеть, что проходящие недавно грибники собрали только шляпки, оставив ножки расти дальше. Смотрится это варварски! Многие, начитавшись литературы о том, что одна часть мягче, чем другая, начинают делить вкусный гриб на части.
Делать это или нет, решать конечно грибнику! Но все же, если не жалко потратить на приготовления блюда немного больше времени, можно срезать и ножку. Она станет прекрасным дополнением к любому супу.
Кроме того, её можно просушить и добавлять в любые рецепты первых блюд в зимнее время.
Шляпки готовить в разы проще. Их просто обжаривают в кляре на сковороде, как отбивные или готовят в сотейнике, стушевывая вместе со сметаной или соусом бешамель. Получается очень вкусно!
Интересно! Наши бабушки тоже собирали Попы в своё время. И чтобы не выкидывать ножки, они перекручивали их на мясорубки, и готовили вместе с картошкой или овощами.
Как вырастить гриб-зонтик на даче в саду?
Выращивание этого гриба не ограничивается одним способом. На сегодняшний день дачники успешно используют два метода выращивания Попов у себя на участке. Первый вариант – произрастание через грибницу, второй – спорами. Выбрать можно любое решение, наиболее приемлемое для себя.
К большому сожалению, приобрести грибницу зонтика в магазине не удастся. Её можно только выкопать самостоятельно в лесу. Правда есть одно «но», – она может попросту не прижиться. Как показывает проведенное учеными исследование, Macrolepiota procera не любит менять местожительство и редко удачно переезжает.
Спорный метод тоже придется добывать самостоятельно. Для этого подойдет шляпка старого зонтика. Её подвешивают на участке с помощью нити или любого другого удобного предмета. После того, как гриб полностью высыхает, споры сами рассеиваются по участку.
Важно! Наиболее успешным будет выращивание зонтиков в лесу. Правда, стоит учитывать, что Ваше грибное место могут навестить другие грибники.
Попы на дачном участке фото
Полезные свойства:
1) Онкология. Содержащиеся в Пёстрых Зонтиках бета-глюканы и меланины, позволяют бороться с раковыми опухолями. И пусть это всего лишь народное средство, отзывы демонстрируют его положительное воздействие на заболевание.
2) Сердце. Сердечно-сосудистая деятельность тоже укрепляется благодаря этим грибам. Они положительно воздействуют на кровообращение и сосуды в целом.
3) Нет депрессии! Этот чудодейственный гриб помогает избавиться от психологического расстройства, улучшая настроение и избавляя организм от умственного истощения.
4) Улучшает обменные процессы.
5) Очищает организм от излишнего кол-ва холестерина в крови.
Противопоказания
Несмотря на то, что плюсов у зонтика множество и он полностью съедобен, ведущие врачи не рекомендую его употребление в следующих случаях:
1) детям, чей возраст не достиг 10 лет; 2) беременным женщинам; 3) людям, страдающим заболеваниями ЖКТ, поджелудочной железы; 4) всем, у кого есть проблемы с печенью;
5) в случае аллергической реакции.
Источник: https://gribysobiraem.info/zontik-pyostryj-grib/
Выдели значимые части слов в названии грибов?
Выдели значимые части слов в названии грибов.
Какое слово из первого текста может послужить проверочным для слова называется?
Какое слово может стать проверочным для слова встречается?
1. растёт гриб около деревьев в густой седой траве.
По шляпке у него, как волны на воде, расходятся розовые круги.
За эти круги – волны и назван гриб ( как?
2. Этот гриб встречается среди мха, где ему тепло почти до самых морозов, поэтому и называется ( как?
Этот гриб встречается среди мха, где ему тепло почти до самых морозов, поэтому и называется ( как?
Ягодкой, гриб, шляпкой, свод, ножкой проверочные слова?
Ягодкой, гриб, шляпкой, свод, ножкой проверочные слова.
Не правда ли , у гриба смешное имя? Маслёнок, будто в нём масло хранят. Масло , конечно, есть. Только не просто , а лесное . Его и хранит маслёнок у себя
под коричневой шляпкой. И шляпка блестит будто масленая . (Найти в тексте однокоренные слова. Определить состав каждого из них . С помощью какой части слова образовано название гриба?)
Однокоренные слова — маслёнок,масло, масленая.
Название гриба образованно с помощью суффиксального способа.(У меня вопрос. Как понять «Определить состав каждого из них»?)
— Маслёнок. — Масл-корень, ёнок-суффикс, нулевое окончание.
Масло- масл — корень, о — окончание.
Масленая — масл — корень,ен — суффикс,-ая-окончание.
корне,рядом записывай проверочные слова.Подчеркни пять слов с безударным
гласным звуком в корне,который ты не можешь проверить.
ТЕКСТ:
Дальнее плавание. Слова-то какие!Вы задумайтесь,молодой
человек,прислушайтесь к музыке этих слов.Дальнее. даль. простор
необъятный. пространство.Не правда ли?А плавание?Плавание-это
стремление вперед,движение,иными словами.Значит так: движение в
пространстве.Тут,знаете,астрономией пахнет.Чувствуешь себя в некотором
роде звездой,планетой,спутником,на худой конец.И все же не в этом
главнаясила,которая заставляет нас покидать родные берега.И если хотите
знать,я вам открою секрет и поясню,в чем тут дело.Удовольствия дальнего
плавания неотрицаемы,что говорить.Но есть большее удовольствие:
рассказать в кругу близких друзей и случайных знакомых о явлениях
прекрасных и необычайных,свидетелями которых вы становитесь в дальнем
плавании.
Это кокой то страный котик не правда-ли??
под старым клёном, как четыре года назад.
(2) Были темно.
(3) — Как же вы поживаете? спросила
Екатерина Ивановна.
(4) — Ничего, живем понемножку, — ответил
Старцев.
(5) И я ничего не мог больше придумать. (6)
Помолчали.
(7) Я волнуюсь, — сказала Екатерина
Ивановна и закрыла руками лицо, — но вы
не обращайте внимания. (8) Мне так
хорошо дома, я так рада видеть всех и не
могу привыкнуть. (9) Сколько
воспоминаний! (10) Мне казалось, что мы
будем говорить с вами без умолку, до утра.
(11) Теперь он видел близко её лицо,
блестящие глаза, и здесь, в темноте, она
казалась моложе, чем в комнате, и даже как
будто вернулось к ней ее прежнее детское
выражение. (12) И в самом деле, она с
наивным любопытством смотрела на него,
точно хотела поближе разглядеть и понять
человека, который когда то любил её так
пламенно, с такой нежностью и так
несчастливо; её глаза благодарили его за
эту любовь. (13) И он вспомнил всё, что
было, все малейшие подробности, как он
бродил по кладбищу, как потом под утро,
утомлённый, возвращался к себе домой, и
ему вдркг стало грустно и жаль прошлого.
(14) В душе затеплился огонек. (15) — А
помните, как я провожал вас на вечер в
клуб? — сказал он. (16) — Тогда шёл дождь,
было темно.
(17) Огонёк все разгарался в душе, и уже
хотелось говорить, жаловаться на жизнь.
(18) — Эх! — сказал он со вздохом. (19) — вы
вот спрашиваете, как я поживаю. (20) Как
мы поживаем тут? (21) Да никак. (22)
Старимся, полнеем, опускаемся. (23) День
да ночь — сутки прочь, жизнь проходит
тускло, без впечатлений, без мыслей. (24)
Днём нажива, а вечером клуб, общество
картёжников, алкоголиков, хрипунов,
которых я терпеть не могу. (25) Что
хорошего?
(26) — Но у вас работа, благородная цель в
жизни. (27) Вы так любили говорить о
своей больнице. (28) Я тогда была какая-то
странная, воображала себя великой
пианисткой. (29) Теперьвсе барышни играют
на рояле, и я тоже играла, как все, и ничего
во мне не было особенного; я такая же
пианистка, как мама писательница. (30) И
(30) И
конечно, я вас не понимала тогда, но потом,
в Москве, я часто думала о вас. (31) Я
только о вас и думала. (32) Какое это
счастье быть земским врачом, помогать
страдальцам, служить народу. (33) Какое
счастье! — повторила екатерина Ивановна с
увлечением. (34) — Когда я думала о вас в
Москве, вы представлялись мне таким
идеальным, возвышенным.
(35) Старцев вспомнил про денежные
купюры, которые он по вечерам вынимал
из карманов с таким удовольствием, и
огонёк в душе погас.
(36) Он встал, что бы идти к дому. (37) —
Вы лучший из людей, которых я знала в
своей жизни, — продолжала она. (39) — Мы
будем видеться, говорить, не правда ли?
(40) Обещайте мне. (41) Я не пианистка, на
свой счет я уже не заблуждаюсь и не буду
при вас ни играть ни говорить о музыке.
(42) Когда вошли в дом и Старцев увидел
при вечернем освещении её лицо и
грустные, благодарные, испытующие глаза,
обращённые на него, то почувствовал
беспокойство и подумал опять: «А хорошо,
что я тогда не женился».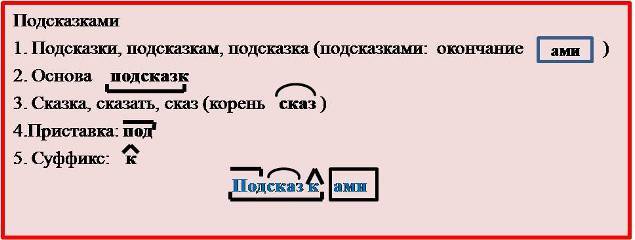
(43) Он стал прощаться
(По А. П. Чехову*)
корне, который ты не можешь проверить. Дальнее плавание. Слова -то какие! Вы задумайтесь , молодой человек, прислушайтесь к музыке этих слов. Дальнее. даль. простор необъятный. пространство. Не правда ли? А плавание? Плавание — это стремление вперёд, движение, иными словами. Значит так: движение пространстве. Тут, знаете, астрономией пахнет. Чувствуешь себя в некотором роде звездой, планетой, спутником, на худой конец. И всё же не этом главная сила, которая заставляет нас покидать родные берега. И если хотите знать, я вам открою секрет и поясню, в чём тут дело. Удовольствия дальнего плавания неоценимы, что говорить. Но есть большее удовольствие: рассказать в кругу близких друзей и случайных знакомых о явлениях прекрасных и необычайных, свидетелями которых вы становитесь в дальнем плавании. Помогите пожалуйста
под коричневой шляпкой. И шляпка блестит будто масленая . (Найти в тексте однокоренные слова. Определить состав каждого из них . С помощью какой части слова образовано название гриба?)
С помощью какой части слова образовано название гриба?)
Однокоренные слова – маслёнок,масло, масленая.
Название гриба образованно с помощью суффиксального способа.(У меня вопрос. Как понять “Определить состав каждого из них”?)
– Маслёнок. – Масл-корень, ёнок-суффикс, нулевое окончание.
Масло- масл – корень, о – окончание.
Масленая – масл – корень,ен – суффикс,-ая-окончание.
Разобрать по составу такие слова ( смешное , простое, лесное , коричневой , блестит, масляная)?
Разобрать по составу такие слова ( смешное , простое, лесное , коричневой , блестит, масляная).
Не правда ли у гриба смешное имя
› Грибы
11.03.2020
Русский язык | 1 – 4 классы
Не правда ли , у гриба смешное имя?
Маслёнок, будто в нём масло хранят.
Масло , конечно, есть.
Только не просто , а лесное .
Его и хранит маслёнок у себя под коричневой шляпкой.
И шляпка блестит будто масленая .
(Найти в тексте однокоренные слова.
Определить состав каждого из них .
С помощью какой части слова образовано название гриба?
Однокоренные слова – маслёнок, масло, масленая.
Название гриба образованно с помощью суффиксального способа.
Как понять “Определить состав каждого из них”?
– Масл – корень, ёнок – суффикс, нулевое окончание.
Масло – масл – корень, о – окончание.
Масленая – масл – корень, ен – суффикс, – ая – окончание.
Выдели значимые части слов в названии грибов?
Выдели значимые части слов в названии грибов.
Какое слово из первого текста может послужить проверочным для слова называется?
Какое слово может стать проверочным для слова встречается?
1. растёт гриб около деревьев в густой седой траве.
По шляпке у него, как волны на воде, расходятся розовые круги.
За эти круги – волны и назван гриб ( как?
2. Этот гриб встречается среди мха, где ему тепло почти до самых морозов, поэтому и называется ( как?
Этот гриб встречается среди мха, где ему тепло почти до самых морозов, поэтому и называется ( как?
Ягодкой, гриб, шляпкой, свод, ножкой проверочные слова?
Ягодкой, гриб, шляпкой, свод, ножкой проверочные слова.
Разобрать по составу такие слова ( смешное , простое, лесное , коричневой , блестит, масляная)?
Разобрать по составу такие слова ( смешное , простое, лесное , коричневой , блестит, масляная).
Шляпка гриба, покрытая темно – коричневой кожицей, держится на крепкой ножке?
Шляпка гриба, покрытая темно – коричневой кожицей, держится на крепкой ножке.
Снизу шляпка затянута белой пленкой.
Когда ее уберешь, откроется нижняя желтоватая сторона шляпки.
ОПРЕДЕЛИТЕ РОД, ЧИСЛО, ПАДЕЖ ПРИЛАГАТЕЛЬНЫХ.
Запишите слова в три столбика по типу орфограмм?
Запишите слова в три столбика по типу орфограмм.
По шляпке у него, как волны на воде, расходятся розовые круги?
По шляпке у него, как волны на воде, расходятся розовые круги.
За эти круги волны и назван гриб (как?
Подберёзовик – съедобный гриб?
Подберёзовик – съедобный гриб.
Шляпка чаще всего коричневого или бежевого цвета, реже – серового.
Ножка высокая, крепкая.
Встречается гриб в смешанных лесах.
Как назвать тест.
Мой первый увиденный гриб – это маленький крепенький маслёнок с тёмно – коричневой, красноватой шляпкой?
Мой первый увиденный гриб – это маленький крепенький маслёнок с тёмно – коричневой, красноватой шляпкой.
Ножка толстая, крепкая и короткая.
Шляпка гриба снизу затянута белой плёнкой.
Когда её уберёшь, откроется чистая желтоватая, лимонного оттенка нижняя сторона шляпки и на ней две – три капли белого молочка.
Именно такте боровые маслята родились в нашем лесочке.
Мы брали самые ядреные, величиной не более колечка, образуемого большим и указательным пальцами.
1. Как вы понимаете лексическое значение прилагательного ЯДРЁНЫЙ в последнем предложении текста?
2. С помощью каких прилагательных автор описывает маслёнок?
Проанализируйте их и сгруппируйте по разрядам.
Прилагательные каких разрядов преобладают в описании?
3. Выпишите из текста прилагательное, употребленное в форме превосходной степени.
Читать еще: Проверка грибов луком при варке
Какие из прилагательных обозначают степень проявления признака (качества, свойства) предметов не с помощью форм степеней сравнения?
Выпишите и разберите эти прилагательные по составу.
На этой странице находится ответ на вопрос Не правда ли , у гриба смешное имя?, из категории Русский язык, соответствующий программе для 1 – 4 классов.
Чтобы посмотреть другие ответы воспользуйтесь «умным поиском»: с помощью ключевых слов подберите похожие вопросы и ответы в категории Русский язык.
Ответ, полностью соответствующий критериям вашего поиска, можно найти с помощью простого интерфейса: нажмите кнопку вверху страницы и сформулируйте вопрос иначе. Обратите внимание на варианты ответов других пользователей, которые можно не только просмотреть, но и прокомментировать.
Обратите внимание на варианты ответов других пользователей, которые можно не только просмотреть, но и прокомментировать.
Не правда ли , у гриба смешное имя? Маслёнок, будто в нём масло хранят. Масло , конечно, есть. Только не просто , а лесное . Его и хранит маслёнок у себя
под коричневой шляпкой. И шляпка блестит будто масленая . (Найти в тексте однокоренные слова. Определить состав каждого из них . С помощью какой части слова образовано название гриба?)
Однокоренные слова – маслёнок,масло, масленая.
Название гриба образованно с помощью суффиксального способа.
(У меня вопрос. Как понять “Определить состав каждого из них”?)
– Маслёнок. – Масл-корень, ёнок-суффикс, нулевое окончание.Масло- масл – корень, о – окончание.
Масленая – масл – корень,ен – суффикс,-ая-окончание.
Другие вопросы из категории
Источник: https://afros-st.ru/griby/ne-pravda-li-u-griba-smeshnoe-imya.html
Запишите слова в три столбика по типу орфограмм?
Запишите слова в три столбика по типу орфограмм.
Швейные иглы?
Слова ИГЛЫ и ШЛЯПКА многозначные слова.
Нужно составить предложение с любым из этих слов.
Как быстро вырасти в росте
Существуют разные способы увеличения роста. Можно быстро вырасти в росте посредством медицины или естественным путем. Рост увеличивают за счет устранения сколиоза или исправления дефектов осанки, в зависимости от особенностей человека.
Этот путь в домашних условиях не легкий и длительный, понадобится немало времени, чтобы получить желаемые результаты. Но тот, кто целеустремлен и четко поставил перед собой цель преодолеет трудности и добьется поставленной цели.
Статистические данные
Ученые утверждают, что самыми маленькими были неандертальцы, жившие в каменном веке. Рост едва составлял 160 см. По словам археологов, самыми рослыми оказались кроманьонцы, походившие на современных людей и выраставшие до 183 см.
Средневековье в очередной раз внесло изменения в рост человека, судя по находкам останков людей или по рыцарским доспехам. Ученые предполагают, рост средневековых людей составлял 160-170 см.
Ученые предполагают, рост средневековых людей составлял 160-170 см.
В XXI веке средний рост мужчин – 172-176 см, женщин – 162-164 см. Девушки растут до 19 лет, а юноши – до 22. Заметные изменения в росте наблюдаются в первый год жизни, за это время рост увеличивается на 25 см. Следующий этап приходится на 4-7 лет, и в период полового созревания – 11-16 лет у мальчиков, у девочек в 10-15 лет.
Важно знать
- В период полового созревания происходит видимый скачок роста.
- В подростковый период организм не успевает за ростом ребенка наращивать мышечную массу и увеличивать вес, поэтому подростки выглядят слишком худыми.
- В подростковом возрасте нельзя садиться на диеты и голодать, это опасно для организма, особенно для мозга, который в период голодания не получает питательных веществ.
Быстро растем правильно питаясь
Правильное и полноценное питание – основа быстрого и здорового роста. Стандартные основы правильного питания: завтрак, обед, ужин. Рассмотрим подробнее, что должно входить в них.
Рассмотрим подробнее, что должно входить в них.
Завтрак. Основная еда в течение дня. Завтракать нужно обязательно. Организм после сна расслаблен, легко принимает и усваивает полезные вещества, поступающие с пищей. Завтрак – залог бодрости на целый день. Если хочется подрасти, ешьте каши на молоке: гречневую, перловую, кукурузную, рисовую, овсяную.
Съеденная утром каша принесет пользу. За завтраком допускается цельнозерновой хлеб, чай, кофе, свежевыжатые соки.
Некоторые предпочитают на завтрак хлопья, звездочки и т.п., которые заливают молоком. Они либо не имеют вообще никаких питательных веществ, стимулирующих рост, либо их ничтожно мало. От такого завтрака лучше отказаться, а хлопья добавлять в рацион питания, чтобы разнообразить.
Обед. Во время обеда употреблять белковую и растительную пищу. Обед традиционно состоит из первого, второго, десерта и напитков.
Супы сытные и легко усваиваются организмом, улучшая пищеварение. В супах много питательных веществ. Суп не влияет на увеличение роста, но активизирует обмен веществ в организме, что немаловажно. Полезны овощные супы и супы-пюре. Супы из бульонных кубиков варить не стоит, в них содержатся вредные вещества.
Суп не влияет на увеличение роста, но активизирует обмен веществ в организме, что немаловажно. Полезны овощные супы и супы-пюре. Супы из бульонных кубиков варить не стоит, в них содержатся вредные вещества.
Из овощей полезны морковь, капуста, бобовые культуры, зелень, сельдерей, лук, ревень. Из фруктов и ягод предпочтение отдают бананам, апельсинам, землянике, чернике и клюкве. Рекомендуют в день съедать не менее 1 кг овощей и фруктов.
Из мяса выбирают нежирные сорта, птицу, печень, почки, рыбу. Мясо лучше отваривать, чем жарить. Молочные продукты: творог, сметана, сливки, молоко, кефир, сыр.
Обед должен быть разнообразным и питательным, порции маленькими и в разумных пределах. Главное – не переедать.
Ужин. Ложиться спать голодным нельзя, но и наедаться на ночь не следует. Жирную и жареную пищу на ночь не рекомендуют. Чтобы способствовать увеличению роста, советуют поужинать молочными продуктами, например, творогом с медом, сметаной, сыром или кефиром. Рекомендуют есть вареные яйца, свежие овощи и фрукты.
Рекомендуют есть вареные яйца, свежие овощи и фрукты.
Некоторые, чтобы поскорее подрасти, готовят коктейль на основе молока и яиц. На 2 стакана молока берут 1 свежее сырое куриное яйцо (можно перепелиное), взбивают блендером и выпивают смесь днем.
Видео советы
Физические упражнения
Без физических упражнений, используя только питание, невозможно вырасти.
Причиной маленького роста считается нарушение работы гормональной системы, которую изменяют при помощи упражнений для зон роста.
Методика роста тренера СССР В.А. Лонского
Перед упражнениями 10-ти минутная разминка и бег. Затем упражнения, на которые тренер советует отвести 25 минут. Это махи ногами, наклоны вперед-назад, влево-вправо, круговые движения руками (предварительно размять кисти рук и локтевые суставы). Упражнения на растяжку, попробуйте сесть на шпагат. Каждое из упражнений выполняют по 10 раз.
Упражнения на растяжку, попробуйте сесть на шпагат. Каждое из упражнений выполняют по 10 раз.
- Занятия на перекладине. Висите на перекладине 4 подхода (два с утяжеленным весом, до 10 кг), каждый по 30 сек. Затем повисите на перекладине вниз головой (ноги прикрепите специальными ремнями), 4 подхода, каждый по 20 сек. Один из них с грузом (5 кг), груз прижимается к груди.
- Пятнадцать минут отводится на прыжки в высоту: 2 подхода по 12 прыжков на левой и правой ноге и 3 подхода по 12 раз на обеих ногах. Старайтесь выше подпрыгивать. Для прыжков подойдет скакалка.
- Затем игра в волейбол или баскетбол (30 минут). Во время игры старайтесь брать все верхние мячи.
- Не менее четырех раз в неделю, по вечерам, упражнения на растяжку. Для растяжки подходят резиновые ремни. Один привяжите к ногам, другой – под мышками и тяните в стороны. Упражнение выполняйте 5-10 минут.
- Плавание. Улучшает работу сердечнососудистой системы и дыхание. Во время плавания работают все группы мышц, позвоночник постепенно растягивается. Для увеличения роста человека плавать 2-3 раза в неделю по одному сеансу.
 Для увеличения роста человека плавать 2-3 раза в неделю по одному сеансу.
Для увеличения роста человека плавать 2-3 раза в неделю по одному сеансу.Видео как вырасти на 5-10 см за 8 недель
Спи и расти
Крепкий, здоровый и правильный сон станет залогом успеха в увеличении роста. Во сне организм вырабатывает гормон роста.
Рекомендации специалистов и врачей о правилах хорошего и здорового сна.
- Спать в проветренной комнате, где тихо и темно. Люди живут в городах, где уровень шума высок, а ночью фонари освещают улицы и от этого в комнате светло. Человек привыкает, но это не способствует сну. Рекомендуют использовать беруши для ушей, а на окна повесить шторы из плотной ткани.
- Кровать должна быть жесткой, так позвоночнику комфортнее. На мягкой постели выспаться не получится. Излишнюю мягкость уберут листы фанеры, подстеленные под кровать и ортопедический матрас.
- Обстановка в комнате должна располагать ко сну. В спальной комнате должно быть уютно, постельное белье красивое и чистое. Крепкий сон на грязно-серых простынях обречен на провал.
- Объемная подушка не подойдет. Ученые утверждают, что человек должен спать без подушки, чтобы улучшить кровообращение. Для увеличения роста спите на спине, подушку положите не под голову, а под колени, которые слегка согните. Довольно странное положение, и привыкнуть к нему нелегко, но пользы от него немало. Во время сна не поджимайте колени к груди и не сворачивайтесь «калачиком», это затрудняет дыхание в связи с уменьшением притока воздуха в легкие.
- По мнению ученых, взрослому человеку, чтобы выспаться, достаточно 6-8 часов. Но все зависит от организма. Кому-то достаточно и 5 часов, чтобы отдохнуть, а кому-то 10 часов, чтобы чувствовать себя свежим и бодрым. В период интенсивного роста, организму необходимо больше сна, поэтому подросткам рекомендуют спать не менее 10 часов. Кто постарше, от 16 до 25 лет для восстановления сил понадобится меньше времени, достаточно поспать 7 или 9 часов.
- Желательно соблюдать график сна – ложиться спать в одно время, а не полуночничать. Идеальное время для сна – с 23 до 2-х часов. Если время упустить, сон не получится здоровым и крепким, и наутро человек чувствует себя «разбитым» и усталым. Шаолиньские монахи утверждают, что спать нужно ложиться в 21 час, а вставать в 7 утра, ни раньше, ни позже. Они выяснили, что в этот период организм быстрее восстанавливается. Чтобы заснуть, на ночь выпивайте стакан теплого молока с медом, который успокаивает и насыщает организм необходимыми витаминами и микроэлементами.
 Излишнюю мягкость уберут листы фанеры, подстеленные под кровать и ортопедический матрас.
Излишнюю мягкость уберут листы фанеры, подстеленные под кровать и ортопедический матрас. Кто постарше, от 16 до 25 лет для восстановления сил понадобится меньше времени, достаточно поспать 7 или 9 часов.
Кто постарше, от 16 до 25 лет для восстановления сил понадобится меньше времени, достаточно поспать 7 или 9 часов.Напоследок остановлюсь на некоторых моментах. Не забывайте, советы индивидуальны. Сходите к врачу, получите совет по нагрузкам, ведь чрезмерное усердие только навредит. Никто не скажет, вырастет человек или нет, но если есть желание и стремление на пути к цели, все получится! Удачи вам!
Загрузка . ..
..Симметричная сборка и разборка экологической сети
Аннотация
Положение видов во взаимодействующих сетях может предсказать вероятность их исчезновения. Однако процессы, посредством которых эти сетевые роли возникают и сохраняются, а затем распадаются во время развития экосистемы, неизвестны. Здесь мы изучаем сети сообществ растений и арбускулярных микоризных грибов (AMF) вдоль хронопоследовательности почвы в 120000 лет, с двумя отдельными фазами: прогрессивной (то есть сборкой, при которой увеличивается богатство растений и биомасса) и регрессивной (т.е. демонтаж, когда богатство растений и биомасса снижаются с уменьшением количества питательных веществ). Мы обнаружили, что порядок формирования взаимодействий во время прогрессии отражает порядок исчезновения взаимодействия во время регресса, и что взаимодействия в сайтах, перемещающихся вперед или назад от (12000-летнего) сайта пиковой биомассы, были вложенными подмножествами взаимодействий в этом сайте. Сборка и разборка сети были симметричными, самоусиливающимися процессами, которые вместе генерировали ключевые атрибуты сетевой архитектуры. Виды растений, у которых было высокое перекрытие партнеров AMF с другими в сообществе (измеряемое как «центральность близости»), были лучше всего способны привлекать и удерживать новых партнеров AMF, а виды AMF с высоким перекрытием партнеров были лучше способны сохранять свои взаимодействия с растениями. Напротив, генерализм взаимодействия («степень узла») сам по себе был плохим предиктором привлечения партнера во время сборки или удержания во время разборки. Эти результаты могут быть использованы для прогнозирования последовательностей вымирания на основе собраний сообщества или сетевых ролей, определения основных точек вторжений и предложения траекторий восстановления экосистемы.
Виды растений, у которых было высокое перекрытие партнеров AMF с другими в сообществе (измеряемое как «центральность близости»), были лучше всего способны привлекать и удерживать новых партнеров AMF, а виды AMF с высоким перекрытием партнеров были лучше способны сохранять свои взаимодействия с растениями. Напротив, генерализм взаимодействия («степень узла») сам по себе был плохим предиктором привлечения партнера во время сборки или удержания во время разборки. Эти результаты могут быть использованы для прогнозирования последовательностей вымирания на основе собраний сообщества или сетевых ролей, определения основных точек вторжений и предложения траекторий восстановления экосистемы.
Введение
Организация взаимодействия видов в экологических сетях может иметь решающее значение для стабильности и функционирования экосистем (1-3). Однако процессы, которые создают и поддерживают структуру сети во время развития экосистемы, не решены. Общепринятая гипотеза сложной сетевой сборки заключается в том, что новые узлы (например, виды) предпочтительно соединяются с теми, которые связаны со многими ближайшими соседями (4) (то есть узлами универсального типа с высокой «степенью»). Было предсказано (5), а затем эмпирически показано (6), что это «предпочтительное прикрепление» произойдет во время сборки сетей опыления в течение сезона. Напротив, исследование долгосрочных изменений в сетях опыления показало, что менее связанные виды предпочтительно привлекают новые взаимодействия (7), так что важность предпочтительной привязанности на основе степени узлов остается неясной для экологических сетей. Хотя исследования по созданию сетевой архитектуры в основном сосредоточены на присоединении узлов во время сборки сети, противоположный процесс предпочтительного отсоединения теоретически может привести к той же архитектуре, что и предпочтительное присоединение (8, 9).Эта возможность подтверждается эмпирическими выводами о том, что сети опыления показывают преимущественную потерю менее связанных видов и их взаимодействий (7, 10). Наконец, возможно, что сетевая архитектура может быть сгенерирована путем предпочтительного присоединения во время сборки сообщества и поддерживаться путем предпочтительного отсоединения во время разборки.
Было предсказано (5), а затем эмпирически показано (6), что это «предпочтительное прикрепление» произойдет во время сборки сетей опыления в течение сезона. Напротив, исследование долгосрочных изменений в сетях опыления показало, что менее связанные виды предпочтительно привлекают новые взаимодействия (7), так что важность предпочтительной привязанности на основе степени узлов остается неясной для экологических сетей. Хотя исследования по созданию сетевой архитектуры в основном сосредоточены на присоединении узлов во время сборки сети, противоположный процесс предпочтительного отсоединения теоретически может привести к той же архитектуре, что и предпочтительное присоединение (8, 9).Эта возможность подтверждается эмпирическими выводами о том, что сети опыления показывают преимущественную потерю менее связанных видов и их взаимодействий (7, 10). Наконец, возможно, что сетевая архитектура может быть сгенерирована путем предпочтительного присоединения во время сборки сообщества и поддерживаться путем предпочтительного отсоединения во время разборки. Это означало бы, что одни и те же правила могут действовать как во время сборки, так и во время разборки экологических сетей (11), но эта гипотеза остается непроверенной.
Это означало бы, что одни и те же правила могут действовать как во время сборки, так и во время разборки экологических сетей (11), но эта гипотеза остается непроверенной.
Здесь мы изучаем сборку и разборку сети взаимодействия в сообществе растений и арбускулярных микоризных грибов (AMF) (12) на протяжении 120 000 лет развития экосистемы. Арбускулярные микоризные грибы являются важными мутуалистами большинства наземных растений (13) и обладают типичным внутриобластным разнообразием, сравнимым с растительными сообществами (14). Более того, высокая важность межвидовых взаимодействий для определения состава сообщества AMF (15, 16) требует сосредоточения внимания на взаимодействиях (17) для этих систем в контексте питательных веществ почвы, которые также могут быть важными факторами структуры сообщества (18).Во-первых, мы проверяем, являются ли процессы сборки и разборки сети случайными или связаны с положением вида в сети. Во-вторых, мы проверяем гипотезу (11) о том, что присоединение и отсоединение симметричны, так что процесс и порядок разборки отражают таковые во время сборки. Наконец, мы используем моделирование, чтобы определить, как неслучайные процессы сборки и разборки влияют на ключевые аспекты сетевой архитектуры.
Первое требование для того, чтобы произошло взаимодействие, — это встреча, и если бы частота встречаемости между пропагулами AMF и растениями была просто стохастической (19), мы могли бы ожидать, что вновь прибывшие виды будут взаимодействовать преимущественно с наиболее многочисленными видами на участке.Установленный вид AMF, который использовал свои ассоциации со многими видами растений для развития обширной сети мицелия, также может быть полезным партнером из-за снижения начальных затрат на строительство и немедленного доступа к большому объему почвы (20, 21). В этом смысле местная численность вида, наряду с количеством видов, с которыми он взаимодействует, может частично указывать на его качество как мутуалиста. Если растения или AMF проявляют селективность в отношении своих партнеров по взаимодействию (22), мы могли бы ожидать, что общая картина новоприбывших видов будет иметь большую вероятность взаимодействия с универсальными видами, т.е.е. традиционная преимущественная привязка по видовой принадлежности (4, 6). Этот образец также может возникнуть просто, если универсалы наименее избирательны в отношении партнеров, которых они принимают. Более того, высокое разнообразие AMF может снизить конкуренцию между видами растений и обеспечить страховку от изменчивости почвенных условий, так что виды растений, которые взаимодействуют со многими AMF, испытают повышенную биомассу (23). Эта большая биомасса может сделать растения-универсалы AMF более привлекательными партнерами, поскольку они обладают способностью обеспечивать больше углерода в более широком диапазоне условий.
Однако преимущества взаимодействия с универсалом могут частично зависеть от того, разделяют ли они своих партнеров по взаимодействию с другими видами. Хотя AMF обычно взаимодействует со многими видами растений, появляется все больше свидетельств того, что их состав (15) и приспособленность могут различаться для разных растений-хозяев (24). Селективность растений-хозяев не обязательно может быть видоспецифичной, скорее группы AMF могут ассоциироваться с экологическими группами растений (25). Близкородственные AMF также имеют тенденцию совместно возникать в данном месте (26), а у близкородственных организмов обычно есть общие партнеры по взаимодействию (27).Таким образом, мы можем ожидать формирования больших взаимодействующих консорциумов экологически схожих, а в некоторых случаях филогенетически связанных растений и грибов на данной стадии сукцессии, при этом доминирующий консорциум потенциально будет отличаться между раннесукцессионными универсальными видами и более поздними сукцессионными лесными видами (14). , 25). Таким образом, на каждой стадии развития экосистемы вновь прибывшие грибы могут иметь более высокую вероятность взаимодействия с растениями в доминирующей экологической группе на этой стадии, которые уже будут взаимодействовать с видами из доминирующей группы грибов.Эти виды растений не только (или даже обязательно) будут наиболее универсальными по отдельности, но они будут иметь высокое перекрытие партнеров AMF с другими видами растений в доминирующей группе на этой стадии (то есть с большинством видов на участке). Виды, у которых партнеры по взаимодействию часто совпадают с другими в сети, также могут получать преимущества в фитнесе за счет положительных косвенных эффектов (1), что делает их более надежными партнерами по взаимодействию. Поэтому мы предполагаем, что привлекательность вида для вновь прибывших может зависеть от его общих партнеров по взаимодействию с другими видами (см.рис.S1 для нашего подхода к измерению перекрытия партнеров).
Чтобы проверить эти гипотезы, мы использовали набор данных (15) по сетям растения-AMF, которые были отобраны в разных местах в долгосрочной (> 120 тыс. Лет) хронопоследовательности, охватывающей все стадии развития экосистемы, включая регресс (28). Хронопоследовательности представляют собой мощный инструмент для понимания долгосрочных скоординированных изменений между видами и их взаимодействий, доступности ресурсов и экосистемных процессов (28). Сильные градиенты от начального N-ограничения до возможного сильного P-ограничения экосистемных процессов стимулируют круговорот растений и видов AMF (15, 29).Мы рассматривали прогрессию экосистемы (во время которой богатство видов растений и увеличение биомассы (29)) как фазу сборки, а регресс экосистемы (во время которой сильное ограничение P вызывает потерю разнообразия растений и биомассы (29)) как фазу разборки сети. Мы определили привязанность как первую стадию экосистемы, на которой наблюдали растение или вид AMF в хроноследовательности, а отрыв — как заключительную стадию экосистемы, на которой наблюдали вид.
Результаты
Всего в наших выборках было обнаружено 33 рабочих таксономических единицы (ОТЕ, далее называемые видами) AMF и 53 вида растений, среди которых мы наблюдали 399 парных взаимодействий (т.е. звеньев, определяемых как колонизация корня растения видами AMF) (рис. 1). В среднем насчитывалось 16 видов растений AMF (SD = 3,33) и 10,8 (SD = 3,77) видов растений с 137,5 (SD = 24,14) взаимодействиями на участок (таблица S1). Изменения сообщества AMF и оборот по хронопоследовательности были подробно описаны в другом месте (15).
Рисунок 1:Матрицы ассоциации растений и микориз на этапах сборки (N = 8 участков) и разборки (т. Е. Регрессивных, N = 3 участков) развития экосистемы.Более темные цвета указывают на то, что ассоциация растение-микориза присутствовала в большем количестве участков (т.е. она образовалась на ранней стадии во время сборки или сохранялась во время регресса). Сеть в значительной степени вложена (см. Результаты SI), в результате чего специалисты взаимодействуют с соответствующими подмножествами видов, которые взаимодействуют с специалистами широкого профиля.
Сначала мы проверили, происходит ли предпочтительное прикрепление или отстранение к существующим видам, у которых много партнеров или много партнеров перекрывается. Вновь прибывшие виды AMF значительно чаще взаимодействовали с видами растений, у которых были общие грибковые партнеры со многими другими (рис.2A, P = 1,4 x 10 -8 ), но не те, у которых много партнеров (т.е. высокая «степень»; P = 0,401) (Таблица S2). Напротив, растения не показали предпочтительного прикрепления на основании перекрытия партнеров (фиг. 2B) или степени AMF (P> 0,4 в обоих случаях, таблица S2). Тем не менее, преимущественное отделение произошло в обоих таксонах, так что растения и AMF были значительно менее вероятно потеряны из сети во время регресса экосистемы, если они взаимодействовали с видами, у которых было высокое перекрытие партнеров с другими в сообществе (P < 1.8 x 10 −5 в обоих случаях; Рис. 2C, D; Таблица S2).
Рисунок 2:Вероятность (левая вертикальная ось), что новый A, C) арбускулярный микоризный гриб (AMF) или B, D) растение, входящее в сеть, будет взаимодействовать с существующим видом во время сборки (вверху) или отделиться от виды во время разборки (внизу) как функция этого вида взаимодействующие партнеры перекрываются с другими на его трофическом уровне (измеряется как центральность близости однораздельной проекции, см. рис. S1). Гистограммы вверху и внизу каждого графика представляют частоты (правая вертикальная ось) единиц и нулей соответственно в необработанных данных.Линии тренда основаны на коэффициентах обратной связи биномиальной линейной модели смешанных эффектов со стадией хронопоследовательности (сеть растительно-микоризная) или годом (сеть торговли оружием) в качестве случайного эффекта. Сплошные линии были статистически значимыми (P <0,005 во всех случаях), пунктирная линия не была значимой при альфа = 0,05.
Как предполагалось ранее (11), процесс разборки отражал процесс сборки; последние взаимодействия, сформировавшиеся во время сборки, были потеряны первыми во время регресса экосистемы (рис.3, P = 1,64 x 10 −9 ). Более того, взаимодействия, присутствующие в сайтах, движущихся вперед или назад от сайта максимальной биомассы, были вложенными подмножествами тех, которые присутствовали в этом сайте (P < 0,001 в обоих случаях), так что взаимодействия постепенно накапливались во время прогрессирования, но последовательно терялись во время регресса. . Таким образом, симметричные процессы сборки и разборки, каждый из которых теоретически может создать сложную сетевую архитектуру (1, 4, 8, 9), оба работали для быстрого создания архитектуры, которая не претерпевала значительных изменений в процессе развития экосистемы (результаты SI, таблица S3, рис.S2).
Рисунок 3:Взаимосвязь между порядком, в котором были сформированы взаимодействия завод-AMF (сетевые «связи») (порядок поступления), и порядком их утраты (порядок исчезновения). Более крупные круги указывают на большее количество перекрывающихся точек данных. Существовала значительная отрицательная связь между порядком формирования и исчезновения взаимодействий (F 1,269 = 39, P <0,0001), так что последние взаимодействия, образовавшиеся во время сборки сети, были первыми, которые терялись во время разборки.Этот анализ включал только взаимодействия, которые присутствовали в месте пика биомассы (порядок прибытия 8), поэтому его не смущали взаимодействия, которые проявлялись только в одном месте (т.е. прибыли и исчезли мгновенно). Взаимосвязь также не была просто обусловлена взаимодействиями, которые проявлялись только в месте максимальной биомассы, поскольку она оставалась значимой даже после удаления этих точек данных (F 1,245 = 5,38, P <0,022).
Хотя степень совпадения и партнерство в наших сетях коррелировали (рис.S3), мы не обнаружили свидетельств преимущественного отделения растений от ОМП низкой степени ( P = 0,577), а таксоны AMF даже имели увеличенную вероятность отделения , когда они ассоциировались с узлами высокой степени (P = 0,040). , как это наблюдалось в долгосрочном исследовании сетей опыления (7), и эта связь стала более сильной, когда в модель также было включено перекрытие партнеров (таблица S2). Более того, во всех наших моделях перекрытие партнеров предсказывало вероятность привязанности или отстранения лучше, чем эквивалентные модели с только степенью в качестве предиктора, и его эффект оставался качественно неизменным, когда степень также была включена в модель (Таблица S2, S4).
Наши результаты показывают, что как привязанность, так и отстранение являются ключевыми процессами в развитии сети, и при их объединении потеря или усиление ссылок становится самоусиливающимся (9). Мы смоделировали наблюдаемые здесь процессы, чтобы изучить последствия для сетевой архитектуры (материалы и методы SI), и обнаружили, что процессы предпочтительного присоединения и отсоединения важны для определения и поддержания вложенности сети (рис. S4), ключевого элемента экологической безопасности. сетевая архитектура (1, 12), при которой специалисты взаимодействуют с видами, которые также взаимодействуют с универсалистами.Предпочтительная привязанность (основанная на степени или совпадении партнеров) порождает сети, которые были гораздо более вложенными, чем случайными, а предпочтительное отстранение сохраняло эту вложенность перед лицом исчезновения видов (рис. S4).
Обсуждение
Очевидно, что виды не имеют информации об архитектуре своих сетей взаимодействия, поэтому перекрытие партнеров должно быть связано с экологическими характеристиками, которые делают виды более склонными к привлечению и удержанию партнеров по взаимодействию.Есть несколько потенциальных, неисключительных объяснений этой привлекательности. Самым простым было бы, если бы у обильных видов было много партнеров из-за высокой вероятности случайных встреч. В нашем наборе данных наблюдалась слабая корреляция между численностью видов и перекрытием партнеров на участках. Более того, ядро взаимодействующих универсалов (т. Е. Виды, участвующие в большинстве взаимодействий в сети и чьи партнеры значительно перекрываются с остальной частью сообщества) присутствовало на протяжении большей части развития экосистемы (см. Темные цвета в верхнем левом углу матриц в Инжир.1), и эта повсеместность, вероятно, сделала их надежной мишенью для вновь прибывших видов. Однако причинно-следственная связь не может быть выведена из этих корреляций в нашем наборе данных, так как привлечение различных грибковых мутуалистов в равной степени может вызвать более долгосрочное сохранение растений (30). Наиболее важно то, что связь между перекрытием партнеров растений и вероятностью прикрепления и отсоединения AMF оставалась значительной после контроля численности (результаты SI, таблица S4). Напротив, эффект степени был устранен или даже обращен вспять, когда мы контролировали численность, и предпочтительное отделение растений от видов AMF с низким перекрытием партнеров стало несущественным при контроле обилия (результаты SI, таблица S4).
Следовательно, хотя численность, вероятно, играет некоторую роль, привлекательность растений с высоким уровнем перекрытия партнеров для таксонов AMF отражает дополнительные преимущества ассоциации с этими видами. Растения могут извлечь выгоду из сообществ AMF, созданных другими видами (31), и конкуренция между растениями может быть уменьшена (и увеличена биомасса растений) за счет высокого разнообразия AMF (23). Таким образом, консорциумы растений, которые способны разделять различные сообщества AMF друг с другом, могут расти лучше и, следовательно, иметь способность обеспечивать больше углерода симбионтами AMF.Эти группировки могут также отражать предпочтение среды обитания вовлеченных видов, при этом универсальные растения и виды AMF тесно взаимодействуют во время ранней сукцессии, а специалисты по лесам — во время поздней сукцессии (25), что соответствует гипотезе о специфичности партнеров, происходящей на уровне экологических групп ( 14).
Потенциальные выгоды от взаимодействия с видами с высоким уровнем перекрытия партнеров могут быть частично нивелированы у вновь появившихся растений, поскольку продуктивность всходов может быть снижена из-за сети AMF крупных гетероспецифических растений, которые конкурируют за AMF-опосредованные ресурсы и истощают запас питательных веществ. (32, 33).Кроме того, высокая универсальность AMF может означать, что между видами меньше вариаций в степени, в которой они являются общими растениями-хозяевами (о чем свидетельствует диапазон значений по оси x для видов AMF, который составляет менее половины, чем у растений (рис. 2A, B)). ). В совокупности эти факторы могут объяснить незначительное влияние степени перекрытия AMF и партнеров на вероятность прикрепления растений (рис. 2B).
Однако и растения, и виды AMF с меньшей вероятностью будут потеряны во время регресса, если они будут взаимодействовать с видами, у которых партнерство часто перекрывается с другими в их сообществе (рис.2В, Г). Мы выдвинули гипотезу (на основе описанных выше преимуществ), что взаимовыгодные взаимодействия будут включать виды с высоким перекрытием партнеров, и мы заметили, что виды, которые преимущественно взаимодействовали с этими основными видами, с гораздо большей вероятностью сохранялись во время регресса. Снижение уровня P в почве во время развития экосистемы, вероятно, делает растения все более зависимыми от P., обеспечиваемого AMF. Аналогичным образом, снижение скорости фотосинтеза растений по хроноследовательности (34) может усилить конкуренцию между AMF за углерод, полученный из растений.Таким образом, потребность развивать и поддерживать мутуалистов и избегать симбиотических «мошенников» будет усиливаться по мере продвижения, особенно во время регресса. Со временем как заводы, так и АИФ могут сократить свои ресурсы, выделяемые менее выгодным партнерам (22), особенно при нехватке ресурсов (35). Следовательно, взаимодействия, которые сохраняются во время регресса, могут быть наиболее взаимовыгодными. Предыдущая работа показала, что виды в поздне-сукцессионных сетях растение-AMF имеют тенденцию чаще взаимодействовать с подмножеством своего общего круга партнеров (36), и поэтому жизнеспособной стратегией может быть сначала тестирование нескольких партнеров, а затем постепенное ограничение. выделение ресурсов подмножеству из них, которые обеспечивают наибольшую выгоду (35) по мере того, как ресурсы становятся ограниченными.В соответствии с этой потенциальной стратегией наиболее универсальные растения имели тенденцию терять многих партнеров AMF во время регресса (вероятность отделения AMF возрастала с увеличением степени растения-хозяина). Этот процесс может легко происходить на уровне отдельных растений, но отражаться в потере связей на уровне видов.
Наконец, если будет обнаружено, что наши выводы можно обобщить на другие экологические сети, у них будет несколько потенциальных приложений. Во-первых, симметрия процессов сборки и разборки (рис.3) на протяжении длительного развития экосистемы предполагает, что знание порядка сборки для данной системы может быть использовано для прогнозирования взаимодействий, подвергающихся наибольшему риску исчезновения, например, при использовании сетевой информации для сохранения (37). Реинтродукция видов во время восстановления может также следовать обратному порядку вымирания и заменять вымершие виды другими, которые выполняют ту же сетевую роль, или фокусироваться на видах, которые имеют общих партнеров по взаимодействию с другими, а затем будут привлекать взаимодействия с любыми новыми видами, которые колонизируют.Точно так же неместные виды должны преимущественно взаимодействовать с видами с высоким уровнем перекрытия партнеров в сети, что, таким образом, может быть в центре внимания мониторинга биобезопасности для раннего обнаружения инвазий.
Невозможно полностью понять архитектуру сложных сетей в отрыве от динамических процессов, которые их генерируют и поддерживают (4). Мы эмпирически показали, что как предпочтительная привязанность, так и непривязанность лежат в основе развития экологической сети в меняющихся абиотических условиях.Эти симметричные, самоусиливающиеся процессы могут создавать и поддерживать важные особенности сетевой архитектуры взаимодействия (1, 4, 8, 9) и, таким образом, связывать колонизацию и вымирание видов с возникающими и потенциально стабилизирующими (1, 2) свойствами экосистемы.
Материалы и методы
Набор данных
Мы создали сети взаимодействий между сосудистыми растениями и арбускулярными микоризными грибами (AMF), используя данные исследования, в котором изучались бета-разнообразие AMF и важность возраста почвы по сравнению срастение-хозяин для объяснения структуры сообщества AMF (15). Методы отбора проб и секвенирования можно найти в этом исследовании или в «Материалах и методах СИ», но здесь мы суммируем ключевые моменты. Отбор проб проводился вдоль хронопоследовательности почвы Франца-Иосифа на южно-западном побережье Южного острова в Новой Зеландии. Важная особенность хронологической последовательности Франца-Иосифа, которая привела нас к выбору этого участка, заключается в том, что сильные градиенты питательных веществ в почве (материалы и методы SI) связаны с выраженными сдвигами в составе, структуре и функциях экологического сообщества, так что развитие экосистемы демонстрирует четкую прогрессивная фаза вплоть до поверхности 12000 лет, на которой наблюдается пиковая биомасса деревьев (мы рассматривали это как фазу сборки), и регрессивная фаза после этого, в которой базальная площадь дерева (биомасса) уменьшается примерно в три раза (29, 38) ( этап разборки).Мы отобрали образцы грибных сообществ на корнях на десяти поверхностях следующего возраста (в годах): <5, 15, 70, 290, 500, 1000, 5000, 12 000, 60 000, 120 000 (SI Материалы и методы; Таблица S1).
На каждом участке мы взяли пробы 50 фрагментов корня, которые затем подверглись молекулярному анализу для идентификации видов растений, а также любых ОТЕ AMF, присутствующих внутри корня (см. (15) или SI Материалы и методы). Мы не присваивали веса этим связям, поскольку гипотезы, которые мы здесь проверяем, относятся к первоначальному формированию взаимодействий, а не к их силе или частоте после того, как они сформировались.
Анализ данных
Мы предположили, что виды, впервые попадающие в сеть во время прогрессивной фазы, будут ассоциироваться неслучайно с другими существующими видами в зависимости от их положения в сети (т. Е. Предпочтительной привязанности (4)). Мы также предположили, что то же самое может произойти во время разборки сети, так что вероятность того, что виды будут локально вымерли из сети, будет зависеть от положения в сети видов, с которыми они связаны (т.е. преимущественное отстранение (8, 10)). Мы проверили предпочтительное прикрепление во время прогрессивной фазы (5-12 000 лет) и предпочтительное отслоение во время регрессивной фазы (12 000 — 120 000 лет), представляя, соответственно, сборку и разборку сети. Затем мы исследовали появление (прикрепление) новых видов в сети во время сборки и исчезновение (отделение) видов во время разборки. Считалось, что вид прикрепился во время прогрессивной фазы, когда он впервые появился в хронопоследовательности.Во время прогрессивной фазы, как только вид прикреплялся (т. Е. Образовывалась связь), считалось, что он останется частью сети до конца развития экосистемы. Это гарантировало, что любые виды, которые появлялись, исчезали, а затем снова появлялись во время прогрессивной фазы, не учитывались дважды, что придало бы любым видоспецифическим предпочтениям взаимодействия этих видов непропорциональный вес в анализе. Наконец, это предотвратило недооценку степени или совпадения партнеров из-за несовершенной выборки сети на любом данном этапе.Точно так же считалось, что отслоение произошло во время регрессивной фазы, когда виды, которые присутствовали во время прогрессирующей фазы, отсутствовали в данном месте и во всех остальных участках последовательности. Обратите внимание на то, что, поскольку по определению процессы привязанности и отсоединения должны происходить в течение интервала между двумя этапами хронопоследовательности (например, взаимодействие присутствовало в сайте B, которого не было в A), мы рассматривали сайт возрастом 12000 лет (который представлял сообщество пик разнообразия и биомассы) как «пиковое» сообщество, включающее последнюю стадию сборки и первую стадию, из которой произошла разборка.
Предпочтительное присоединение и отсоединение
Мы проанализировали процессы предпочтительного присоединения и отсоединения с использованием обобщенных линейных моделей смешанных эффектов с биномиальными ошибками и канонической логит-функции связи, выполненных в пакете lme4 (39) в R (40). Мы использовали отдельные модели для проверки процессов прикрепления во время прогрессирования по сравнению с процессами отрыва во время регресса и для растений по сравнению с AMF. Переменная отклика в каждой модели была бинарной, при этом каждый существующий вид в сети для каждого участка был закодирован значением 0 (вновь прибывший вид не прикрепился к нему или вид не отделился от него на этом участке. ) или 1 (новый вид, прикрепленный к виду на этом участке или отделившийся от него).Участок (поверхность вдоль хронопоследовательности) был включен как случайный фактор, так что предпочтительное прикрепление было проверено в соответствии с относительными сетевыми ролями различных видов в пределах участка, а не путем сравнения видов по участкам, на которые могли влиять различия в размерах. сетей. Фиксированной переменной-предиктором была степень или центральность (определено ниже).
Чтобы измерить перекрытие в использовании партнеров по взаимодействию между видами на заданном трофическом уровне, мы использовали центральность близости (далее «центральность», определенную на рис.S1) в однораздельной проекции сети двудольного взаимодействия (41). Центральность часто коррелирует со степенью (42), но несет дополнительную информацию, измеряя количество связей от основного вида ко всем остальным в сети (а не только к ближайшим соседям), и используется в качестве меры важности вида (42). ) или функциональной специализации (43) в мутуалистических сетях. В однодольной проекции виды на данном трофическом уровне связаны, если у них есть общий партнер по взаимодействию (рис.S1), столь высокая центральность указывает на то, что у вида есть много партнеров с другими видами, которые также имеют общих партнеров со многими другими. Напротив, виды с низкой центральностью взаимодействуют с партнерами, которые не используются доминирующим консорциумом тесно взаимодействующих растений и AMF. Хотя центральность может неявно использоваться для вывода о природе потоков в сети (например, пути против обходов, репликация против распространения), мы не делаем никаких предположений о природе потоков, если таковые имеются, в сети AMF завода.Скорее, мы используем центральность близости просто как меру степени, в которой разные виды разделяют партнеров по взаимодействию на трофическом уровне. Центральность была рассчитана в пакете «sna» (44) в R, масштабированном в пределах данной сети (то есть в пределах каждого сайта).
Мы проверили, зависит ли присоединение к узлу во время сборки или отсоединение от узла во время разборки от центральности этого узла (рис. S1) или степени (количество партнеров, с которыми взаимодействует вид, нормализованное в сетях данного узла). возраст).Мы использовали центральность / степень узла на этапе хронопоследовательности непосредственно перед тем, на котором был присоединен новый узел или отсоединен существующий узел. Это позволило нам сравнить соответствие моделей с центральностью и степенью в качестве предиктора с использованием информационного критерия Акаике (AIC). Таким образом, прогностической переменной в каждой модели была степень или центральность каждого существующего вида на участке, к которому вновь прибывшие виды либо прикреплялись, либо не прикреплялись, либо отделялись от них. Для полноты картины мы также запустили модели с обоими этими предикторами, чтобы определить, изменила ли какая-либо потенциальная коллинеарность качественно их отдельный эффект.Ни одна из наших моделей не показала признаков чрезмерной дисперсии. Мы повторили эти анализы, контролируя численность видов, в случае, если усилия по отбору проб приводили к каким-либо ложным эффектам (см. «Контроль влияния численности видов» в «Материалы и методы SI»).
Мы исключили из этого анализа любые связи, представляющие новые виды, прикрепляющиеся к другим вновь появившимся видам, потому что мы не могли приписать значение положения в сети для этих видов, чтобы использовать их в качестве переменной-предиктора. Однако эти пары между двумя вновь появившимися видами были редкими, и вовлеченные виды обычно также взаимодействовали с другими видами, уже присутствующими в сети.На всех участках 20 новых видов растений были связаны с вновь прибывшими AMF, и из них 16 видов также были связаны с видами AMF, которые уже присутствовали в сети, для которых можно было рассчитать положение в сети. Точно так же из 15 вновь появившихся видов AMF, прикрепившихся к вновь появившимся растениям, 8 также прикрепились к существующим видам.
Порядок появления и исчезновения
Мы проверили, был ли порядок появления и исчезновения (исчезновения) связей в экологической сети во времени случайным, или ссылки, появившиеся первыми во время прогрессии, терялись в последнюю очередь из сети во время регресса.Чтобы проверить это, мы проанализировали порядок появления и исчезновения каждого звена (т.е. каждой ассоциации между микоризными видами и видами растений). Этот анализ включал как связи вновь появившихся видов (как в приведенном выше анализе предпочтительной привязанности), так и связи, которые сформировались между существующими видами. Порядок прибытия определялся как ранжируемый возраст сайта, на котором взаимодействие было впервые обнаружено; например взаимодействию, наблюдаемому в самом раннем (<5-летнем) сайте, было присвоено значение 1, тогда как взаимодействию, впервые наблюдаемому в 1000-летнем сайте (6 th вдоль последовательности), было присвоено значение 6.Мы рассматривали сеть в месте пика биомассы растений (29) (возраст 12000 лет) как конечную точку фазы сборки, затем исследовали порядок, в котором взаимодействия были потеряны во время фазы регресса после этого пика (то есть порядок исчезновения). . Порядок исчезновения был определен так же, как и порядок прибытия, как первый участок после участка пика биомассы, в котором взаимодействие не наблюдалось. Например, взаимодействию, которое присутствовало на участке пика биомассы, но не на последних двух участках, был присвоен порядок исчезновения 1, взаимодействию, которое присутствовало на участке пика биомассы, и обоим оставшимся участкам был присвоен порядок исчезновения 3. , так далее.Для анализа мы использовали только те взаимодействия, которые присутствовали в месте пика биомассы, чтобы быть консервативными; если бы мы просто использовали порядок прибытия и порядок исчезновения (последний сайт, в котором произошло взаимодействие) всех взаимодействий в последовательности, это привело бы нас к получению значимого результата, потому что любые взаимодействия, которые появлялись только один раз в последовательности или появлялись рано и исчезли в районе пика биомассы, сгенерировала бы автоматическую корреляцию между порядком их прибытия и порядком исчезновения.
Таким образом, наш анализ был сфокусирован на сети участка пиковой биомассы и проверил, коррелирован ли порядок, в котором взаимодействия вступают в сеть, с порядком, в котором они были потеряны из сети. Мы проверили взаимосвязь между порядком прибытия и исчезновения, используя простую регрессию, с порядком исчезновения в качестве переменной отклика и порядком прибытия в качестве переменной-предиктора, и каждое взаимодействие (комбинация растение-микориза) было повторением. Оставалась возможность, что любая наблюдаемая взаимосвязь может быть вызвана взаимодействиями, которые проявляются только в месте пика биомассы, и, следовательно, они появятся последними и первыми вымрут.Чтобы учесть это, мы повторно провели анализ после удаления взаимодействий, которые присутствовали только в месте пика биомассы.
Отрицательная корреляция между порядком появления и исчезновения взаимодействий предполагает постепенное накопление взаимодействий в сети вплоть до места пиковой биомассы с последующей постепенной потерей взаимодействий. Ожидается, что это сгенерирует паттерн, согласно которому участок пика биомассы имеет набор взаимодействий, из которых постепенно более старые или молодые участки имеют уменьшающиеся подмножества.Чтобы проверить такую закономерность, мы сгенерировали две матрицы, в которых мы ранжировали участки в соответствии с их возрастом (строками), становясь либо постепенно старше, либо моложе участка максимальной биомассы, который находился в верхнем ряду. Ячейки матриц отображали наличие или отсутствие каждой попарной ассоциации растение-AMF (столбцы). Мы протестировали каждую матрицу на вложенность, используя метрику вложенности, основанную на перекрытии и убывающей заливке (NODF), с 999 перестановками и алгоритмом обмена, используя функцию oecosimu в пакете vegan (45) для R (40).
Моделирование
Чтобы изучить последствия предпочтительного присоединения и отсоединения в зависимости от центральности и степени, мы провели ряд моделирования сценариев сборки и разборки сети (более подробно см. В разделе «Материалы и методы SI»). На каждом временном шаге вновь появившиеся растения или виды AMF будут взаимодействовать с одним существующим видом с вероятностью, пропорциональной степени существующего вида (то есть модель Барабаши и Альберта (4)) или перекрытия партнеров (центральность близости).Мы также запустили третий сценарий, согласно которому привязанность была случайной (равновероятная привязанность к любому существующему виду), чтобы обеспечить нулевую точку сравнения.
Эта фаза сборки продолжалась до тех пор, пока в сети не было 50 видов растений и 50 видов AMF. Впоследствии каждая сеть была подвергнута фазе разборки с использованием того же сценария (на основе степени, на основе централизации или случайного), который использовался во время сборки этой сети. Чередующиеся виды растений и AMF были удалены (то есть вымерли) с вероятностью, которая была обратно пропорциональна степени или близости центральности видов, с которыми они взаимодействовали, так что виды, которые взаимодействовали с видами высокой степени или высокой центральности, с меньшей вероятностью были вымерли.Как и на этапе сборки, мы запустили сценарий фазы разборки, в которой вымирание было случайным.
Каждый сценарий этапов сборки и разборки запускался для 1000 повторов (каждый из которых состоял из нескольких временных шагов). На каждом временном шаге в каждой реплике, после того, как виды прибыли или вымерли, мы регистрировали вложенность сети с помощью NODF, рассчитанную с помощью функции nestednodf в веганском пакете (45) для R. Вложенность — общая черта мутуалистических сетей (46). ), который недавно также наблюдался в сетях микориз с растениями (12) и, как было показано, увеличивает устойчивость сетей (1, 2).Мы масштабировали наблюдаемый NODF по распределению значений NODF, полученных путем рандомизации матрицы смежности с использованием нулевой модели (SI Materials and Methods).
Вклад авторов
J.M.T. и I.A.D. разработал исследование в обсуждениях со всеми другими авторами. L.B.M.-G. и I.A.D. проведены молекулярные анализы. J.M.T. проведен статистический анализ. S.J.R. и D.A.P. собраны данные о численности растений. Все авторы внесли свой вклад в опробование и написание рукописи.
Информация об авторе
Авторы заявляют об отсутствии конкурирующих финансовых интересов.Переписку и запросы материалов следует направлять в J.M.T. ([email protected]).
SI Результаты
Не было изменений в сетевой структуре вдоль хронопоследовательности
Предыдущая работа показала, что некоторые атрибуты сетевой структуры могут быть быстро обнаружены даже с небольшими выборками из сетей (37). Это говорит о том, что общая сетевая архитектура, такая как вложенность взаимодействий, может быстро возникнуть во время развития экосистемы (то есть даже когда сеть небольшая), при условии, что процессы предпочтительного присоединения и отсоединения происходят с самого начала формирования сети.
Таким образом, мы проанализировали, как вложенность и другие метрики сетевой структуры менялись в хронопоследовательности, используя общие линейные модели (или обобщенные линейные модели Пуассона в случае богатства) с метрикой в качестве переменной отклика и возрастом сайта в журнале в качестве предиктора. В качестве показателей для описания сетевой структуры мы протестировали два показателя вложенности (температура вложенности и NODF), взаимосвязь, как количественные, так и бинарные версии плотности сцепления, количественную равномерность взаимодействия и видовое богатство растений и микориз.Чтобы учесть возможность того, что взаимосвязь между показателями сети и возрастом сайта была нелинейной, мы включили квадратичный член для возраста сайта. Мы также включили размер выборки (количество образцов корней, в которых были идентифицированы как виды растений, так и хотя бы один вид грибов) в качестве ковариаты для учета любых потенциальных смещений выборки. Затем мы упростили каждую из этих моделей, удалив переменные и минимизировав информационный критерий Акаике (AIC).
В наших сетях вложенность существенно не изменилась по хронопоследовательности (таблица S3), как можно было бы ожидать, если бы сила предпочтительного прикрепления была постоянной по хронопоследовательности.Фактически, ни одна мера структуры не изменилась с развитием экосистемы (таблица S3, рис. S2), что согласуется с предыдущими исследованиями сборки сети опыления и структур пищевой сети (6, 73, 74). Напротив, сравнение ранних и поздних сукцессионных лесов показало, что два (связность и специализация) из шести показателей структуры сети растение-AMF изменились (36), хотя эти изменения могли быть вызваны изменениями в разнообразии потенциальных партнеров по взаимодействию. Несмотря на макроскопически статичную структуру, которую мы наблюдали, происходил высокий оборот видов и взаимодействий, при этом новые виды присоединялись к сети, а другие больше не встречались на каждой стадии последовательности (Таблица S1).
Влияние обилия видов на результаты
Влияние процентного покрытия растений на центральность было значительным (t = 3,24, P = 0,002), хотя величина эффекта была небольшой (наклон = 0,000217). Например, если процентное покрытие вида изменилось на 11% (что было бы эквивалентно переходу от нижнего квартиля к верхнему квартилю), соответствующее значение центральности увеличилось бы только на 0,002 (что на порядок меньше единицы). стандартное отклонение центральности, sd = 0.029). Тем не менее, мы отмечаем, что процентное покрытие растений значительно прогнозировало нормализованную степень растений (t = 6,55, P = 2,46 x 10 -9 ) с большей величиной эффекта (наклон для влияния процентного покрытия на нормализованную степень = 0,010). Обилие таксонов AMF в образцах достоверно предсказывало их центральность (логарифм преобразован, чтобы сделать взаимосвязь линейной: t = 8,32, P <0,0001) и нормализованную степень (t = 21,71, P <0,0001).
Таким образом, неудивительно, что сетевое положение видов коррелировало с их численностью.Однако, несмотря на это, влияние центральности растений на вероятность прикрепления и отделения грибов оставалось значительным (Таблица S4), даже после контроля численности видов (т.е. включения численности в качестве ковариаты для проверки того, влияет ли центральность на вероятность прикрепления или отделения на заданном уровне избыток). Мы использовали надземный покров как меру численности растений, которая имела то преимущество, что измерялась независимо от наших ассоциаций растение-AMF. Однако в случае, если надземный покров не коррелировал с обилием корней, мы повторили анализ с обилием каждого вида растений в образцах корней в качестве ковариаты, и влияние центральности растения на прикрепление грибов оставалось значительным (Таблица S4).Фактически, численность растений (с использованием любой меры) не коррелировала в значительной степени с вероятностью прикрепления AMF (Таблица S4), хотя наблюдалась незначительная тенденция увеличения вероятности отделения AMF с увеличением численности растений. В отличие от этой последовательной связи между прикреплением или отрывом и центральностью, связь между нормализованной степенью и вероятностью прикрепления или отсоединения AMF стала несущественной, когда численность (либо надземный покров, либо численность в пробах) была включена в качестве ковариаты (| t | < 1, P> 0.343 в обоих случаях). Это указывает на то, что любая вариация в вероятности прикрепления или отсоединения, относящаяся к нормализованной степени, также коллинеарна численности растений из-за сильной корреляции между численностью и степенью.
Растения не показали какого-либо предпочтительного прикрепления в исходных анализах (Таблица S2), и добавление численности в качестве ковариаты не изменило этого (Таблица S4). Однако ранее значимое влияние центральности AMF на вероятность отделения растений стало несущественным, когда численность таксонов AMF в образцах была добавлена в качестве ковариаты в модели.Более того, незначительная тенденция к положительной корреляции между степенью AMF и вероятностью отслоения растений стала значительной, когда численность была включена в модель, в отличие от предыдущих исследований, показывающих, что виды с высокой степенью с большей вероятностью сохранят взаимодействия, а их партнеры имеют пониженная вероятность вымирания (7, 10).
Рис. S1 Измерение партнерского перекрытия.Иллюстрация сети взаимодействий между растениями и арбускулярными микоризными грибами (AMF).Сеть можно представить в виде двудольной матрицы, где черные квадраты указывают на наличие взаимодействия («ссылки») в сети. Односторонняя проекция этой сети также может быть сгенерирована для каждого трофического уровня (здесь показано красным, только для AMF), посредством чего виды связаны между собой, когда у них есть общий партнер по взаимодействию. Эта односоставная проекция изображает наложение партнеров по взаимодействию между видами. Затем мы измерили перекрытие партнеров каждого вида как «центральность близости»: инверсию средней длины пути (т.е.е. «Степени разделения») от основных видов ко всем остальным в односоставной проекции. Это расстояние также называют «функциональной специализацией» (43). Например, вид E напрямую связан с видами C, D, F и G, имеет два шага к виду B (через C) и три шага к виду A (через C и B). Это дает виду E средняя длина пути 1,5 к любым другим видам, что является самой короткой средней длиной пути и наивысшей центральностью близости среди любых видов AMF. Виды A и G имеют одинаковое количество звеньев в однодольной проекции (1 звено), но у G есть общий партнер-растение с видом E, который имеет общих партнеров со многими другими в сети (средняя длина пути G = 2.3). Напротив, A имеет общего партнера с видом B, который сам имеет общих партнеров только с A и C. Таким образом, A имеет более низкое общее перекрытие партнеров (средняя длина пути = 2,83), потому что у него нет общих партнеров с растениями в самом большом кластере взаимодействующие виды.
Рис. S2 Сети Plant-AMF вдоль хронопоследовательности.Показаны количественные сети (при этом ширина звеньев указывает их относительную частоту). Однако для анализа использовались бинарные (несвязанные невзвешенные) сети, потому что частота ассоциации не является объективной мерой частоты взаимодействия в ассоциациях растительно-микоризные (69).
Рис. S3 Взаимосвязь между нормализованной степенью и центральностью.Поскольку мы сравнили предпочтительную привязанность, основанную на совпадении взаимодействий и партнеров (определяемую как «центральность близости», рис. S1), с аналогичным процессом, основанным на степени, мы проверили, коррелированы ли степень и центральность. Мы провели гауссовские модели смешанных эффектов с нормализованной степенью (логарифмическое преобразование) в качестве переменной-предиктора и центральностью близости в качестве ответа. Чтобы учесть независимость видов от данного участка, возраст участка был включен как случайный эффект.Модели со смешанными эффектами проводились в пакете lme4 (39) для R. Значения P для моделей оценивались с использованием степеней свободы, рассчитанных с помощью метода синтеза знаменателя Саттертуэйта, выполненного в пакете lmerTest (75) для R На рисунке степень логарифмически преобразована, а центральность близости представлена как остатки после удаления вариации из-за случайных эффектов. Все отношения были значимо положительными (модели со смешанными эффектами: t >, P <0.0001 во всех случаях).
Рис. S4 Изменения во вложенности взаимодействий во время моделирования фаз сборки и разборки.Вложенность (верхняя строка необработанного NODF, нижняя строка отклонения от нулевого ожидания) во время моделирования прибытия (левый столбец) и исчезновения (правый столбец) растений и видов AMF. Каждый сценарий запускался для 1000 повторов. Темные горизонтальные линии показывают медианные значения, прямоугольники показывают верхний и нижний квартили, а усы показывают минимумы и максимумы (с точками, обозначающими крайние значения). Сценарии представляли собой предпочтительную привязанность или непривязанность, основанную на степени (синий) или центральности близости (фиолетовый) партнеров по взаимодействию (или инверсии степени и центральности для непривязанности).Для сравнения серым цветом выделены случайные вымирания. Фазы сборки и разборки заметно повлияли на вложенность сети. На этапе сборки сценарий, основанный на предпочтительном подключении к узлам с высокой степенью интеграции, создавал сети с высокой степенью вложенности и значительно большей вложенностью, чем в двух других сценариях. Вложенность имеет тенденцию увеличиваться с увеличением размера сети (со временем), что подчеркивает необходимость сравнения необработанной вложенности с нулевой моделью. По сравнению с нулевой ожидаемой вложенностью, только сценарий, основанный на степени, имел увеличивающуюся вложенность с течением времени.Напротив, сценарии на основе центральности и случайной сборки показали небольшое снижение вложенности с течением времени (поскольку добавление новых узлов размывало наследие начальной матрицы, которая была сильно вложенной). Несмотря на это, сценарий, основанный на центральности, всегда был более вложенным, чем сценарий случайного присоединения. Важно отметить, что различия между сценариями, основанными на степени и центральности, были обращены вспять на этапе разборки. По мере того, как все больше видов удалялось, сценарий на основе степени показал быстрое снижение вложенности, тогда как сценарий, основанный на центральности, стал немного более вложенным, чем нулевое ожидание (и более вложенным, чем сценарий, основанный на центральности).
Таблица S1: Детали сайта.Подробная информация о местоположении участков (широта = широта, долгота = долгота), стадии развития экосистемы, количестве отобранных таксонов растений и арбускулярных микоризных грибов (AMF) и взаимодействиях (связях) между ними (с кумулятивным числом до, и включая этот сайт в скобках), а также условия сайта. Дополнительные сведения можно найти в Richardson et al. (30) и обновлено в Holdaway et al. (69).
Таблица S2: Таблицы коэффициентов из моделей прикрепления и отрыва для растений и арбускулярных микоризных грибов (AMF).Таблицы коэффициентов для компонента фиксированных эффектов обобщенных линейных смешанных моделей, проверяющих корреляцию между вероятностью привязанности и отстранения и (близостью) центральностью (наша мера перекрытия взаимодействий-партнеров, см. Рис. S1) или степенью. Переменные ответа являются двоичными (присоединены / отсоединены или не присоединены / отсоединены), и использовалось биномиальное распределение ошибок с функцией логит-связи. Все модели включали сайт в качестве случайного фактора для контроля за независимостью любых новых ассоциаций, возникающих в одной и той же точке хронопоследовательности.Отдельные модели представлены для (нормализованной) степени и центральности, как описано в основном тексте. Дополнительные модели представлены как с центральностью, так и со степенью, чтобы продемонстрировать, что значимость и направление воздействия центральности не меняется (хотя в некоторых случаях изменяется степень влияния). Для каждой переменной ответа жирным шрифтом выделена наиболее подходящая модель (самый низкий AIC).
Таблица S3: Никаких значительных изменений в сетевой архитектуре в хронопоследовательности.Результаты наиболее подходящих моделей для каждого показателя структуры сети микориз и растений (ответная переменная) и любых переменных-предикторов, которые были сохранены после упрощения модели.Максимальная модель содержала размер выборки (количество выборок, дающих идентифицируемую последовательность), возраст сайта и квадратичный член возраста сайта (чтобы учесть нелинейные отношения) в качестве предикторов. Мы сравнили все подмножества этой максимальной модели и выбрали наиболее подходящую модель с самым низким показателем AIC. Приведены необработанные значения P, хотя они неконсервативны, поскольку многократное тестирование увеличивает вероятность ошибки типа I. Если предикторы не указаны, наиболее подходящая модель содержала только точку пересечения.Никакая мера структуры сети существенно не изменилась в хронопоследовательности.
Таблица S4: Таблицы коэффициентов для моделей предпочтительного прикрепления и отрыва с центральностью в качестве предиктора, контролирующего численность.Таблицы коэффициентов для компонента фиксированных эффектов обобщенных линейных смешанных моделей, проверяющих корреляцию между вероятностью привязанности и отстранения и центральностью близости, нашей мерой перекрытия взаимодействий-партнеров (см. Рис. S1). Эта таблица аналогична таблице S2), но с численностью видов, включенной в качестве ковариаты.Обилие относится к количеству партнера по взаимодействию (а не к прикрепляющимся видам) и измеряется с использованием процентного покрытия надземного покрова для растений и обилия в образцах (количество корней, в которых это произошло) для грибов. Для полноты картины мы также представляем модели с численностью растений, измеренной с использованием количества образцов, в которых она встречалась (как с грибами).
Благодарности
Мы благодарим P.J. Bellingham, T. Fukami, J.H. Джонс, С. Павар, Д. Стоуфферу и сотрудникам лаборатории Тилианакиса / Стоуффера за критические обсуждения и комментарии к рукописи.Н. Болстридж и К. Митчел оказали ценную лабораторную помощь. Исследование финансировалось за счет основного финансирования исследовательских институтов Crown от Группы науки и инноваций Министерства бизнеса, инноваций и занятости Новой Зеландии и стипендии Rutherford Discovery для JMT. Этот документ является вкладом в инициативу Имперского колледжа «Грандиозные задачи в области экосистем и окружающей среды».
Review — Посмертный анализ устаревших литий-ионных батарей: методология разборки и методы физико-химического анализа
Увеличение срока службы — важный вопрос при разработке литий-ионных аккумуляторов.Механизмы старения, ограничивающие время жизни, можно эффективно охарактеризовать с помощью физико-химического анализа старых клеток с помощью множества дополнительных методов. В этом исследовании содержится обзор современной литературы по посмертному анализу литий-ионных элементов, включая методологию разборки, а также методы физико-химической характеристики материалов аккумуляторных батарей. Подробная схема посмертного анализа выведена из литературы, включая предварительный осмотр, условия и безопасную среду для разборки ячеек, а также разделение и постобработку компонентов.Особое внимание уделяется характеристике состаренных материалов, включая аноды, катоды, сепараторы и электролит. В частности, подробно рассматриваются микроскопия, химические методы, чувствительные к поверхностям электродов или к объему электродов, и анализ электролитов. Методы дополняются электрохимическими измерениями с использованием методов реконструкции электродов, встроенных в половину и полные ячейки с электродом сравнения. Критически обсуждаются изменения, происходящие с материалами в процессе старения, а также способность рассмотренных методов анализа их наблюдать.
Литий-ионные батареив настоящее время используются в повседневных объектах, таких как смартфоны, электроинструменты и планшетные компьютеры, а также в растущих областях легких электромобилей (LEV), беспилотных летательных аппаратов (БПЛА), аккумуляторных электромобилей (BEV). , гибридные электромобили (HEV) и подключаемые гибридные электромобили (PHEV). 1–4 Кроме того, рост возобновляемых источников энергии, таких как энергия ветра и солнца, которые доступны только периодически, требует надежных и очень гибких стационарных решений для хранения энергии, которые обеспечивают высокую емкость и предсказуемый срок службы. 2,5
Старение литий-ионных аккумуляторов является общей проблемой для производителей, поскольку они должны гарантировать долгосрочную надежность своей продукции. Для современных ячеек эффекты деградации на уровне материала приводят к снижению емкости и увеличению сопротивления на уровне элементов. 6–28 Состояние старения батареи часто характеризуется состоянием здоровья (SOH) в% согласно 3,16,22,29–31
, где t представляет время старения. В общем, нужно различать цикл 7,16,18,21,23–25,32 и календарное старение. 7,19,21–24,27 Поскольку коммерческие литий-ионные элементы могут подвергаться календарному старению в период между производством и доставкой, рекомендуется измерять разрядную емкость при t = 0 для каждой ячейки, которая подвергается воздействию тест на старение. Поскольку разрядная емкость зависит в основном от температуры, глубины разряда (DOD) и тока разряда, SOH обычно контролируется регулярными проверками с определенными наборами параметров, 7,16,21,23,24 , которые могут варьируются в зависимости от приложения.Обычно температура 25 ° C, 16,22,24 DOD 100%, 16,21 и скорость разряда 1C 7,16,21,22,24 или ниже 23 используются в осмотры.
Снижение производительности на уровне элемента в основном связано с реакциями химического разложения материала и на уровне электродов (см. Рисунок 9). 3,9,15–17,25,28,33–41 В этом смысле полное понимание механизмов деградации, происходящих внутри клеток, имеет решающее значение для увеличения времени их жизни.
Увеличить Уменьшить Сбросить размер изображения
Рис. 9. Обзор механизмов старения, касающихся деградации электрода и разрушения материала. Обратите внимание, что механизмы деградации электрода могут происходить как на анодной, так и на катодной стороне, хотя на этой иллюстрации они показаны только для одного конкретного электрода. Указаны методы анализа для наблюдения соответствующих явлений. В скобках указаны методы с ограниченным доступом к механизму старения.
Чтобы сделать вывод о механизмах старения, необходимо разобрать клетки и проанализировать соответствующие компоненты клетки. Для глубокого понимания процесса старения батареи важна гомогенизированная процедура, включающая вскрытие, разборку, обработку образцов и анализ, чтобы избежать повреждения, загрязнения и модификации компонентов ячейки, а также для получения интерпретируемых данных.
Однако, как показано на корпусе почти каждого коммерческого литий-ионного элемента, разборка не рекомендуется производителями.Это связано с угрозами безопасности, например возможность создания коротких замыканий при открытии ячейки, что может привести к тепловому разгоне ячейки. Кроме того, существуют серьезные проблемы со здоровьем, которые возникают из-за химических соединений и риска повреждения образцов из-за неправильной обработки. 30 Однако при соблюдении определенных протоколов разборка литий-ионных элементов безопасна и дает надежные результаты в отношении состава встроенных материалов и изменений во время старения.
В 2011 году Williard et al.представили методику анализа вышедших из строя литий-ионных аккумуляторов, например после теплового разгона. 30 Однако, насколько нам известно, не существует стандартного метода разборки и анализа устаревших литий-ионных элементов, хотя до сих пор было проведено множество исследований, включающих безотказную разборку устаревших батарей. 12,16,17,25,26,28,32,42–47
В этой статье мы рассматриваем современные методы разборки старых литий-ионных элементов, а также физико-химические методы анализа материалов из разобранных элементов.Для каждого метода обсуждаются выявленные механизмы старения и наблюдаемые изменения на уровне материала, происходящие при старении. Особое внимание уделяется вопросу, какие изменения можно наблюдать с помощью конкретных методов анализа. Наконец, мы сделаем вывод о комбинациях методов, чтобы получить полное представление о процессах старения.
Предварительный осмотр и неразрушающие методы перед вскрытием литий-ионных элементов
Обзор отдельных этапов посмертного анализа приведен на рисунке 1.Перед разборкой клеток полезны методы неразрушающей характеристики, чтобы получить первое представление о механизмах старения. В дополнение к испытаниям емкости (см. Уравнение 1), инкрементный анализ емкости (ICA) 48,49 и спектроскопия электрохимического импеданса (EIS) являются мощными методами для получения информации о механизмах старения. 37,50–52 ICA основано на dQ / dV по сравнению с . В составляет график и, следовательно, преобразует точки плато и перегиба напряжения на кривых напряжения в пики dQ / dV. 48 Изменения в пиках dQ / dV (интенсивности пиков и сдвиги пиков) можно отслеживать во время старения и делать выводы о потере активного материала / потере электрического контакта, изменениях химического состава ячейки, недостаточном разряде, недостаточном заряде, 48 и снятие покрытия Li. 53 Удаление Li также было определено с помощью анализа дифференциального напряжения (DVA) в dV / dQ по сравнению с . Q графиков. 53
Увеличить Уменьшить Сбросить размер изображения
Рисунок 1. Блок-схема разборки литий-ионных элементов и анализа компонентов.
EIS — еще один неразрушающий метод определения характеристик старых клеток. 25,34,37,39,50–52 Во время старения импеданс ячейки обычно увеличивается, что приводит к замедлению кинетики, что частично является причиной уменьшения емкости. 34,37 Причиной увеличения импеданса являются физико-химические процессы внутри ячеек, такие как увеличение резистивных слоев. 25,39,50 Klett et al. обнаружили существенные различия в графике Боде для клеток с календарным и циклическим возрастом. 39 Было обнаружено, что основной причиной этого различия является более выраженная пленка на поверхности анода для циклического старения. 39 Однако на импеданс ячейки влияет множество факторов, требующих моделирования. 50 Более простой и быстрый метод получения базовой информации об изменениях импеданса ячеек — это измерения только на одной частоте, обычно 1 кГц. 28 Такие измерения позволили обнаружить прямую корреляцию между увеличением импеданса при старении и увеличением Mn, P и Li на графитовых анодах с помощью анализа Post-Mortem. 28
Хотя неинвазивные электрохимические методы являются мощным инструментом для получения информации о механизмах старения, прямое наблюдение химических изменений возможно только с помощью посмертного анализа. Кроме того, явления локализованного старения, представляющие только малую часть электродов, часто не видны при электрохимических измерениях, поскольку они усредняются по всем электродам ячейки.
После электрохимической характеризации, визуальный осмотр, графическая документация и взвешивание являются следующими разумными шагами в анализе старых литий-ионных элементов.Это может указывать на внешнюю деформацию или утечку, которые могут повлиять на поведение при старении или привести к отказу ячейки. Кроме того, эти шаги могут дать первые подсказки о наилучшем положении для открытия ячейки. Хотя для стандартных конструкций ячеек, таких как ячейки 18650 или 26650, позиции разреза в большинстве случаев схожи (~ 1 мм рядом с положительным или отрицательным разъемом), может потребоваться проведение дополнительных тестов для других геометрических форм, таких как призматические и карманные ячейки.
Неразрушающие методы, позволяющие выявить внутреннюю часть батарей, включают рентгеновский анализ, 30,54–56 рентгеновскую компьютерную томографию (КТ), 9,42,54–65 и нейтронную томографию. 66,67 Поскольку рентгеновский анализ дает двумерные изображения пропускания (рис. 2а), в зависимости от конструкции ячейки может потребоваться проведение измерений пропускания рентгеновских лучей при нескольких углах обзора. 55
Увеличить Уменьшить Сбросить размер изображения
Рисунок 2. Примеры неразрушающего контроля литий-ионных элементов. a) Рентгеновское изображение ячейки с намотанным желеобразным валиком 30 (с любезного разрешения Springer Science and Business Media).б) Фронтальная компьютерная томография возле положительного разъема ячейки типа 18650 32 (Воспроизведено с разрешения Электрохимического общества). Пунктирными линиями показаны возможные положения резки корпуса ячейки без проникновения электродов.
Напротив, данные КТ получают путем поворота ячейки с небольшими угловыми шагами, в то время как рентгеновские изображения записываются для каждого угла. Из этого набора данных трехмерная модель ячейки определяется математическим алгоритмом, который позволяет рассчитывать осевые и фронтальные двумерные разрезы в определенных положениях (см. Рис. 2b и рис. 3a).Таким образом, КТ является дорогостоящим методом и обычно требует более длительного времени измерения по сравнению с измерениями пропускания рентгеновского излучения. Кроме того, CT приводит к большему количеству данных и большей нагрузке на интерпретацию этих данных. Однако компьютерная томография способна выявить многие детали внутреннего устройства батареи, такие как деформации внутри ячеек после старения, напряжение 9,32,42 , отказ 64 , 54–56,58 или тесты на неправильное использование. 57,59,60 В случае внутренних деформаций КТ очень полезна для изображения их формы без приложения механической силы, которая могла бы изменить ячейку. 32,42,64 (см. Рисунок 3a).И рентгеновский анализ, и компьютерная томография подходят для определения позиций разреза при открытии клеток, как показано на Рисунке 2.
Увеличить Уменьшить Сбросить размер изображения
Рис. 3. a) Осевая компьютерная томография ячейки 18650 с деформированным желеобразным валиком. Область в поперечном сечении клетки на (b) отмечена на компьютерной томографии. б) СЭМ-изображение поперечного сечения ячейки, на котором видны такие детали, как трещины в покрытии. 32 (Воспроизведено с разрешения Электрохимического общества).
Методы, использующие нейтроны, также подходят для получения информации о макроскопическом дизайне внутри литий-ионных ячеек 66 и даже могут доставить химическую информацию 66,68 неразрушающим способом. Однако из-за очень больших усилий, связанных с этим методом, нейтронная томография нецелесообразна для определения позиций разреза для разборки ячейки. Также следует иметь в виду, что образец может быть радиоактивным после обработки нейтронами.
Разборка литий-ионных ячеек при фиксированном SOC и в контролируемой среде
Для поддержания компонентов ячеек в очень похожем состоянии, как во время работы, и для безопасности экспериментатора, как состояния ячейки, так и среда разборки должна быть четко определена.
Перед разборкой элемент необходимо зарядить или разрядить до определенного состояния заряда (SOC). 12,16,17,30,34,45,69–71 С точки зрения безопасности, глубокий разряд (до напряжения конца разряда 0 В) желателен, так как он снижает энергосодержание сотовый. В случае нежелательного короткого замыкания глубокий разряд снизит риск теплового разгона.
С другой стороны, напряжение элемента не должно выходить за пределы нормального рабочего окна, чтобы избежать нежелательных изменений материала, которые не вызваны старением.Поэтому большинство авторов перед разборкой разряжают элементы до напряжения конца разряда, соответствующего SOC = 0%. 12,16,17,28,30,34,46,69,70,72 Определенный SOC также важен для сопоставимости результатов различных ячеек, например старые и свежие клетки одного типа. К сожалению, большинство авторов не предоставляют точную процедуру разгрузки перед разборкой. Кобаяши и др. упомянул, что напряжение холостого хода (OCV) старых элементов, разряженных до 2,5 В при C / 20, было больше, чем OCV свежих элементов из-за увеличения внутреннего сопротивления элемента. 12 Следовательно, авторы держали все ячейки при 3,0 В более 10 часов перед разборкой, что привело к OCV 3,0 В ± 0,01 В. 12 Аналогичный метод разряда был использован Takahara et al. 26 Kumaresan et al. разряжали клетки мешочка в два этапа, сначала с помощью C / 33 и после 30-минутного периода отдыха с помощью C / 83, чтобы обеспечить полную разрядку. 73
Разборка клеток на более высоких SOC была проведена для Т-клеток, 70 , емкость которых очень мала (~ 0.2 мАч), поэтому риск невелик по сравнению с коммерческими батареями (несколько Ач). Burns et al. недавно открыла коммерческие аккумуляторные ячейки 0,22 Ач при ~ 50% SOC и обнаружила, что после циклирования с высокими токами происходит покрытие литием. 45 Те же авторы открыли также элементы типа 18650 3,4 Ач после разряда до 0 В из соображений безопасности 45 из-за их большей емкости. Следовательно, покрытие Li больше не было напрямую видимым (но явные различия в цвете и текстуре отрицательного электрода), хотя этого и следовало ожидать из измерений кулонометрии. 45 Это несоответствие было связано с глубоким разрядом до 0 В. 45
Поскольку некоторые компоненты литий-ионных элементов реагируют с O 2 и H 2 O, перчаточный ящик заполнен атмосферой высокочистого аргона. содержащие H 2 O и O 2 только в нижнем диапазоне ppm, должны использоваться. 30,32,34,45–47,69–71,73–78 Особенно Li x C 6 , металлический Li и LiPF 6 проявляют реакционную способность по отношению к компонентам воздуха.LiPF 6 реагирует с водой с образованием газа HF, 30,36,79 , который может вызвать значительные проблемы со здоровьем без соответствующих средств защиты 30 , а также коррозию катодных материалов. 36 Мы отмечаем, что использование N 2 в качестве инертного газа не подходит из-за его реакционной способности с металлическим Li с образованием Li 3 N. 80 В своей статье от 2002 г., Aurbach et al. использовали наполненный аргоном перчаточный ящик с содержанием O 2 от 5 до 10 ppm и содержанием H 2 O от 2 до 5 ppm. 34 Williard et al. предполагают содержание как O 2 , так и H 2 O ниже 5 частей на миллион. 30 Большинство других авторов не комментируют верхние значения загрязнения в перчаточных ящиках.
В некоторых случаях защита образцов от воздуха менее важна. 30 Примерами являются измерения промытых катодных материалов методом XRD или ICP-OES. Авторы рекомендовали вытяжной шкаф с производительностью 60–100 футов в минуту в качестве минимального требования для разборки небольших коммерческих ячеек после езды на велосипеде в нормальных условиях. 30 Следовательно, Amanieu et al. открыли 18650 ячеек внутри перчаточного бокса, наполненного аргоном, по соображениям безопасности, однако после удаления электролита с помощью DMC образцы LiMn 2 O 4 были высушены в постоянном потоке воздуха вытяжного шкафа в течение ночи, поскольку образцы были не чувствителен к воздуху. 74 Отметим, что безопасность при разборке ячеек на воздухе зависит еще и от влажности. Открытие ячеек во влажном воздухе также имеет решающее значение и может привести к критическим условиям, приводящим к пожарам в лаборатории.
В любом случае старые электроды, которые используются для получения повторно собранных ячеек (см. Раздел «Электрохимический анализ собранных электродов»), должны храниться в перчаточном ящике 12,34,73,81 , прежде чем они будут помещены в герметичную ячейку. Kostecki et al. провели вскрытие и промывку ячеек в перчаточном ящике, наполненном аргоном, и хранили образцы электродов в герметичной ячейке в перчаточном ящике до дальнейших исследований. 69 Отметим, что образцы электродов, которые находятся в контакте с электролитом, портятся даже при герметичной герметизации, поэтому мы рекомендуем использовать электроды для дальнейших электрохимических испытаний в день разборки.
Hightower et al. использовали специальную защиту, покрывая образцы Li x C 6 инертной жидкостью (Fluorinert FC-43) внутри перчаточного бокса, наполненного аргоном, перед переносом через воздух в вакуумную камеру ТЕМ-устройства, где инертная жидкость испарялась во время эвакуация камеры. 82
На этом этапе мы пришли к выводу, что разборку литий-ионных элементов следует проводить в химически инертной среде, например в перчаточном ящике, наполненном аргоном. Даже если элементы разряжены до напряжения конца разряда, демонтаж устаревших литий-ионных элементов все равно должен производиться с большой осторожностью.Процедура и, следовательно, затраты на разборку ячейки в решающей степени зависят от рисков для оператора и чувствительности материалов к воздуху и влаге.
Процедура открытия литий-ионного элемента и разделение компонентов
Внешнее короткое замыкание может произойти из-за непреднамеренного прикосновения к внешним контактам, например токопроводящими инструментами, металлической чешуей во время резки или контактом с металлической поверхностью перчаточного ящика. В зависимости от конструкции ячейки корпус ячейки может быть подключен либо к положительной, либо к отрицательной клемме.Это можно легко определить с помощью вольтметра перед разборкой.
Кроме того, во время открытия ячейки необходимо соблюдать осторожность, чтобы предотвратить внутренние короткие замыкания ячейки, 30,32,54,74 , а также образцов. 30 Внутреннее короткое замыкание наиболее вероятно при разрезании корпуса ячейки либо из-за проникновения, либо из-за деформации пакета электродов / желейного валика, либо из-за механического давления. Следовательно, перед открытием литий-ионных ячеек необходимо определить идеальное положение разреза для каждого типа ячеек, применяя неразрушающие методы, как показано выше в разделе «Предварительный осмотр и неразрушающие методы».Кроме того, выгодно использовать токонепроводящие инструменты, например из керамики или с непроводящим покрытием.
Aurbach et al. представили специальное устройство для открытия 18650 ячеек, которым можно управлять внутри перчаточного ящика. 34 В этом устройстве цилиндрическая ячейка вращается с помощью двигателя с дистанционным управлением, а крышка корпуса ячейки срезается пилой с твердосплавным наконечником. 34 Как показано на рисунке 4a, инструмент Dremel также можно использовать для открытия ячеек. После того, как крышка ячейки снята (рис. 4b), необходимо разрезать выступы, соединенные с корпусом.Затем можно разрезать дно ячейки и, наконец, разрезать кожух вдоль оси цилиндра и развернуть рулон с желе (рис. 4c).
Увеличить Уменьшить Сбросить размер изображения
Рисунок 4. Процесс открытия ячейки. a) –c) Использование инструмента Dremel при открытии ячейки цилиндрического типа. d) –f) Использование керамических ножниц при открытии ячейки мешочка.
Ячейки мешочка, как правило, легче открывать, как показано на рисунках 4d – 4f, поскольку фольгу мешочка можно просто разрезать керамическими ножницами 30 или ножом.В случае призматических ячеек было предложено сделать неглубокий надрез режущим инструментом на одной стороне ячейки, прежде чем снимать оставшуюся оболочку с помощью изолированных плоскогубцев. 30 В любом случае открытие ячейки должно производиться очень осторожно и не допускать чрезмерного усилия на валок с желе или пакет электродов.
Образование металлической пыли или стружки зависит от метода резки. Пыль может попасть в элемент и загрязнить материалы, 30 , тогда как стружка может достигать нескольких мм в длину и может создавать короткие замыкания, ведущие к нежелательной разрядке элемента и выделению тепла.Кроме того, следует учитывать, что во время резки также возникает местный нагрев, который может вызвать изменение материалов ячеек или даже привести к проблемам с безопасностью.
В большинстве случаев компоненты ячеек будут отделены друг от друга, чтобы анализировать их отдельно (см. Рисунки 4c, 4f). Для состаренных анодов может случиться, что активный материал прилипнет к сепаратору, 32 , что приведет к проблемам с разделением компонентов. Это может быть решено погружением анода и сепаратора в DMC.Напротив, для старых катодов это часто менее проблематично.
Типичные конфигурации ячеек представляют собой намотанные рулоны с желе в цилиндрических ячейках, плоские рулоны с желе в призматических ячейках и ячейках мешочка, а также уложенные друг на друга электроды / сепараторы, z-образные сепараторы или комбинации укладки и намотки в мешочках и призматических ячейках. К сожалению, большинство авторов не комментируют этот этап разборки ячейки. Следует отметить, что необходимо соблюдать особую осторожность, чтобы избежать перекрестного загрязнения при контакте между анодом и катодом.Если присутствует электролит, прямой контакт анода и катода вызывает короткое замыкание, приводящее к последствиям, описанным выше.
С самого начала открытия капли электролита могут быть собраны, если они содержатся в достаточном избытке. 83 В противном случае следует отобрать пробу электролита, погрузив рулон с желе сразу после извлечения гильзы в CH 2 Cl 2 84 или разделенные смачиваемые компоненты в ацетонитриле. 85 Этот последний метод позволяет извлекать электролит, а также соединения, образующиеся при его разложении при старении на каждом электроде.Поскольку многие растворители электролита очень летучие, рекомендуется быстрое извлечение электролита, чтобы состав не изменился.
Последующая обработка образцов из разобранных литий-ионных элементов
После разделения компонентов ячейки большинство экспериментаторов промывают эти компоненты типичными растворителями электролита, такими как DMC, 12,16,17,25,26,28,46, 47,65,71,72,74,82,86–90 DEC, 70,91 и EMC, 69 , тогда как только некоторые авторы не проводили промывку своих образцов. 34,45,53,65,78,92 Это возможно, когда требуется только визуальный осмотр 45,53 и / или электрохимические испытания. 46,65 Немытые электроды могут содержать остаточный кристаллизованный LiPF 6 или нелетучие растворители, которые трудно отличить от элементов в SEI или интеркалированном Li. Кроме того, стадия промывки также полезна для уменьшения коррозии образцов, поскольку LiPF 6 реагирует с H 2 O и O 2 и, как уже упоминалось, для защиты чувствительного аналитического оборудования, если образцы подвергаются воздействию воздуха.Somerville et al. показали, что промывка не требуется для удаления ЭК и других типичных карбонатов, когда образцы помещены в вакуум (~ 10 -4 кПа), например в вакуумных устройствах, таких как XPS или SEM. 89
К сожалению, большинство авторов не комментируют процедуру промывки (время, температура, объем, тип растворителя) 26,74,82 , хотя она может существенно повлиять на результаты. Бах и др. вымачивали свои образцы в течение 60 минут в DMC, а затем на 30 минут в новом DMC. 65 Williard et al. прокомментировал, что промывание может привести к отсутствию определенных компонентов SEI. 30 Abraham et al. продемонстрировали, что ополаскивание ДМК должно удалять изолирующие частицы, осевшие на поверхности графита после старения. 93 Недавно Somerville et al. подробно исследовал эту тему для графитовых анодов с пленками, образованными различными количествами добавки ВК. 89 В зависимости от количества ВК в электролите и, следовательно, от состава пленки, также было обнаружено, что SEI может быть изменен, по крайней мере, частично, промывкой DMC. 89 В одном конкретном случае LiPF 6 и LiF были полностью удалены, а частицы LiP x F y были восстановлены через 1 мин. 89 Согласно их исследованию, продолжительность промывки и / или промывки или ее отсутствия должны быть проверены для каждого химического состава клетки. 89
Исходя из нашего опыта, для удаления следов соли Li из образцов требуются две стадии промывки чистым растворителем продолжительностью от 1 до 2 минут. Кроме того, для получения сопоставимых результатов важно всегда выполнять этапы стирки одинаково.
Некоторые методы, такие как анализ ICP-OES 16,28 , используют активный материал, соскобленный с электродов. Такая механическая обработка не изменяет химический состав и, следовательно, не вызывает проблем. XRD возможен как с электродами, так и с соскобленным порошкообразным материалом, однако следует учитывать, что предпочтительные ориентации частиц в электродах, которые не присутствуют в снятом материале, могут приводить к различиям в интенсивностях пиков. 34
Если исследовать трещины в активном материале, можно подготовить поперечные сечения всего литий-ионного элемента (см. Рисунок 3b).В этом случае кожух ячейки не снимается. Вместо этого резка непроводящим полотном пилы выполняется через всю ячейку. Положение разреза можно определить заранее с помощью компьютерной томографии (см. Рис. 3а). После разрезания ячейки электролит удаляется, после чего следует стабилизация с помощью эпоксидной смолы и этап металлографической полировки. 30,32,42,56,94,95
По сравнению с компьютерной томографией, поперечные сечения клеток являются более дорогостоящими с точки зрения труда и приводят к разрушению клетки.Однако поперечные сечения ячейки могут обеспечить значительно более высокое разрешение для определенных частей ячейки (сравните рисунки 3a и 3b), а также возможность выполнять измерения с помощью других мощных методов, таких как резка сфокусированным ионным пучком (FIB) 42,74 и наблюдение с помощью оптической микроскопии, 18,19,30,42,74,94,95 SEM, 30,32,42,74,95–97 или EDX. 95,97 Поперечные сечения комплектных ячеек обеспечивают толщину электродов в рабочем состоянии (соответствующий уровень заряда), т.е.е. с таким же давлением, как и в закрытой ячейке. Следует отметить, что это не относится к поперечным сечениям одиночных электродов, 18,19,74,96,97 , которые могли расшириться после разделения компонентов ячейки.
В этом разделе рассматриваются методы физико-химической характеристики материалов аккумуляторных батарей. Обсуждаются данные о механизмах старения, полученные соответствующими методами, чтобы дать обзор возможностей наблюдения конкретных механизмов деградации.
На рисунке 5 показана схема основных компонентов ячейки и соответствующие доступные методы физико-химического анализа для их характеристики. Образцы могут происходить из анода, катода, сепаратора, токосъемника или электролита, однако для упрощения на рисунке 5 в качестве примера показан только катод. Из рисунка 5 видно, что можно выделить разные части твердого образца: им можно назначить поверхность электрода, объем, поперечные сечения и различные методы анализа, соответственно.
Увеличить Уменьшить Сбросить размер изображения
Рис. 5. Обзор компонентов внутри литий-ионной батареи и физико-химические методы определения характеристик после посмертного анализа.
Причины поверхностной чувствительности методов анализа связаны с физической природой задействованных типов излучения или частиц. Упрощенный обзор физических / химических принципов (облучение и / или обнаружение электронов e — , электромагнитное излучение / фотоны hv , нейтральные частицы и ионы) показан на рисунке 6.Они кратко объяснены для каждого метода в разделах ниже. Более подробные сведения о механизмах возбуждения и обнаружения соответствующих методов анализа можно найти в учебниках. 98–102
Увеличить Уменьшить Сбросить размер изображения
Рисунок 6. a) –k) Упрощенная схема обнаруженных частиц в различных методах анализа. Электромагнитное излучение hv соответствует видимому свету на (a), (f), (g), рентгеновскому излучению на (d), (e), (k) и радиоволнам на (j).На (b) испускаемые электроны можно различить между обратно рассеянными электронами, оже-электронами и вторичными электронами. з) ИК в режиме отражения. м) Принцип разделения компонентов смеси в хроматографии. Кружки и линии представляют молекулы образца и неподвижную фазу соответственно.
Поверхностная чувствительность создается либо путем отражения излучения / частиц на поверхности образца (например, когда электрод исследуется с помощью оптической микроскопии), либо из-за короткой длины свободного пробега частиц внутри твердых образцов (например.грамм. методы с участием е — или ионов). Типичными поверхностно-чувствительными методами являются микроскопия, EDX, XPS, IR или SIMS.
Напротив, другие методы нечувствительны к поверхности и включают информацию из массы электрода. В этом случае материал образца необходимо соскрести, например, при анализе ICP-OES, либо образец не препятствует обнаруженному излучению (например, рентгеновские лучи в случае XRD).
Типичные методы посмертного анализа клеточных компонентов и выявленные ими механизмы старения обсуждаются отдельно в следующем разделе.Однако из-за чувствительности методов к разным частям образцов, упомянутых выше, обзор возможностей каждого метода приведен в разделе «Комбинация методов для полной характеристики механизмов старения».
Микроскопия
Оптическая микроскопия
Оптическая микроскопия основана на отражении видимого света от поверхности образца (рис. 6а). В общем, разрешение оптических микроскопов ограничено дифракционным пределом Аббе, соответствующим диапазону 0.2 мкм. 99 Это позволяет разрешать частицы в диапазоне мкм с меньшими усилиями по сравнению с методами электронной микроскопии. 18,19,30,42,74,95 Следовательно, можно обнаружить эффекты старения, такие как изменения толщины электрода 18,19 или отложения на поверхности электродов, которые находятся в диапазоне размеров мкм. 18,78 Из-за ограниченного разрешения оптической микроскопии обнаружение трещин частиц или очень тонких пленок затруднено или может не наблюдаться.Однако оптическая микроскопия — очень эффективный метод получения обзора поверхности образца.
Brand et al. наблюдали выгорание сепаратора с помощью оптической микроскопии после встряхивания 18650 ячеек. 64 Некоторые группы исследовали осаждение Li и образование дендритов во время процесса зарядки in situ с помощью оптической микроскопии. 96,103–109 Кроме того, изменение цвета графитовых 96 и рутиловых электродов 110 контролировалось in situ с помощью оптических микроскопов.
В случае разрабатываемых новых электродных материалов оптическая микроскопия также оказалась полезной. Pharr et al. смогли использовать методы оптической микроскопии для определения энергии разрушения тонкопленочных электродов из литированного Si в зависимости от концентрации Li. 111 Ли и Федкив успешно изучили влияние наночастиц диоксида кремния, добавленных в гелевые электролиты, на предотвращение коррозии алюминиевых токосъемников. 112
Это лишь несколько эксклюзивных примеров, демонстрирующих использование оптической микроскопии в посмертном анализе литий-ионных клеток, однако они демонстрируют широкий диапазон возможностей этого метода характеризации, которые часто недооцениваются.
Сканирующая электронная микроскопия
Сканирующая электронная микроскопия (SEM) обеспечивает улучшенное разрешение по сравнению с оптической микроскопией из-за меньшей длины волны де Бройля электронов по сравнению с видимым светом. Разрешение СЭМ в основном ограничено сферической аберрацией электронных линз. 99 Кроме того, при интерпретации изображений SEM необходимо учитывать тот факт, что вместо фотонов используются электроны (рис. 6b). Контрастность изображения сильно зависит от выбранного детектора, который собирает либо обратно рассеянные, либо вторичные электроны. 99 Кроме того, наблюдения SEM ограничены вакуумом, что приводит к испарению летучих компонентов, таких как карбонатные растворители.
Из-за более высокого разрешения СЭМ наблюдаемые области могут быть намного меньше по сравнению с оптической микроскопией. Следовательно, необходимо очень внимательно записывать данные, которые являются репрезентативными для всей выборки. Обычно для этого сначала записываются обзорные изображения, а затем масштабируются различные части образца. С помощью сканирующего электронного микроскопа обычно наблюдаются различные клеточные компоненты, поскольку он дает основную информацию о микроструктуре, которая может быть связана с механизмами деградации.
Кроме того, SEM ограничивается наблюдениями за поверхностью образца. Чтобы получить информацию об объеме и / или химическом составе, SEM обычно дополняется другими методами. Например, SEM часто сочетается с EDX-анализом для определения химического состава и / или комбинируется с методами поперечного сечения, такими как металлографическая подготовка, 30,32,42,56,94–96,113 FIB резка, 39, 42,74,114–119 или ионное измельчение. 120 Кроме того, удаление тонких срезов с помощью FIB и последующее сканирование с помощью SEM позволяет создавать видеоролики 119 и создавать 3D-модели электродов 116–118,121 (томография FIB / SEM).Такие трехмерные модели электродов были полезны в многомасштабных расчетах, где учитывалась микроструктура электродов. 118,121
С другой стороны, графит является наиболее распространенным анодным материалом, и в сочетании с другими методами сканирующая электронная микроскопия принесла значительные результаты для выявления механизмов деградации, происходящих на поверхности этого материала.
Рост границы раздела твердых электролитов (SEI) на поверхности частиц графита во время старения наблюдался с помощью SEM 16,18,40 (см. Верхнюю часть рисунка 7).Рост SEI при старении связан с разложением электролита и является причиной потери Li и, следовательно, падения емкости. 16,18,28,33,122
Увеличить Уменьшить Сбросить размер изображения
Рис. 7. СЭМ-изображения графитовых анодов и катодов до и после циклирования 40 (Воспроизведено с разрешения Электрохимического общества).
Другой механизм старения — это осаждение металлического Li на графитовых анодах.Honbo et al. изучили осаждение Li на графите с помощью SEM и выявили дендритную и зернистую морфологию на чистом и измельченном угле соответственно. 123 Zier et al. показали, что можно улучшить контраст материала при осаждении лития на графитовых электродах за счет реакции с OsO 4 . 119 Исследование других анодных материалов, таких как Li 4 Ti 5 O 12 с помощью SEM, до сих пор не дало точной информации о механизмах разложения. 124
На катодной стороне часто с помощью SEM-изображения не видно изменений между нетронутыми и состаренными катодами 16,40,125 (см. Нижнюю часть рисунка 7). Когда сообщалось о видимой поверхностной пленке после продолжительного цикла на поверхности LiCoO 2 , было невозможно связать это с явным механизмом разложения. 34 С другой стороны, механическое напряжение 126–128 из-за изменения объема во время цикла приводит к трещинам в частицах, которые можно наблюдать с помощью SEM. 42,74,75,129,130
В дополнение к механизмам старения, затрагивающим материалы электродов, деградация других компонентов ячейки, такая как коррозия алюминиевых токосъемников 133 и закрытие пор 14,134,135 или плавление 64 сепараторов, являются наблюдается с помощью SEM.
Просвечивающая электронная микроскопия
По сравнению с SEM, просвечивающая электронная микроскопия (TEM) обычно использует более высокие ускоряющие напряжения для электронов, позволяя проходить сквозь материалы (рис. 6c) и с более высоким разрешением вплоть до атомного масштаба. 119,136,137 Таким образом, ПЭМ раскрывает характеристики образца с точки зрения морфологии частиц, кристалличности, напряжения или даже магнитных доменов. Однако из-за более высокой энергии необходимо учитывать повреждение луча для материалов батареи. 138 Следует отметить, что измерения ПЭМ ограничены локализованными областями образца, и поэтому трудно точно исследовать большую выборку.
Как и для всех микроскопических методов, подготовка образцов и их мониторинг во время сбора данных имеют решающее значение для ПЭМ, например.грамм. Обрезка FIB оказалась полезной. Кроме того, большое значение имеют размер (чем тоньше, тем лучше) и чистота образца. Более высокие усилия при подготовке образца делают ПЭМ более трудоемким методом по сравнению с СЭМ. Несколько обзоров экспериментальных возможностей и сравнения с другими микроскопическими методами можно найти в учебниках. 101,102
Структурные изменения морфологии частиц в результате календарного и циклического старения были исследованы Watanabe et al.для LiNi 0,8 Co 0,15 Al 0,05 O 2 катодный материал. 139,140 ТЕМ-анализ также дал ценную информацию о зависимости связующего вещества от производительности элемента, 141 образования SEI на катодах 142 и оценке новых электродных материалов. 110,143–146
В этом обзоре были упомянуты только несколько возможностей ПЭМ в посмертном анализе, однако использование этого метода смещается от метода посмертной характеристики к методу in situ и операнду . . 147 Эта тенденция становится все более очевидной по мере того, как приборы предоставляют аналитическую аппаратуру с низким Z-элементом, контроль окружающей среды, а высокоскоростные и чувствительные прямые детекторы электронов становятся все более доступными. 148
Методы химического анализа, чувствительные к поверхности электродов
Энергодисперсионная рентгеновская спектроскопия
Энергодисперсионная рентгеновская спектроскопия (EDX) часто сочетается с приборами SEM. Принцип EDX основан на облучении образца электронами и обнаружении генерируемых характеристических рентгеновских фотонов (рис. 6d).Таким образом, EDX позволяет получить информацию о химическом составе образца. Кроме того, если поверхность образца сканируется электронным лучом, карты химического состава на поверхности могут быть созданы путем наложения с изображениями SEM (EDX-картирование). 39,95,97,119,149,150 Однако EDX представляет собой серьезный недостаток, поскольку он не может обнаруживать Li. Следовательно, необходимы дополнительные методы для его обнаружения и количественной оценки. 16,17,72,150
Анализ EDX, выполненный во время посмертных исследований, позволяет проверить состав активных материалов и обнаружить присутствие дополнительных фаз.Например, EDX позволил обнаружить повторное осаждение растворенного Mn на поверхности графитового электрода после растворения катодов из смеси NMC / LiMn 2 O 4 . 16,28 Аналогичным образом, Klett et al. наблюдали Fe на анодах после растворения с катодов LiFePO 4 . 17 Также возможно измерить присутствие F и P на анодах из-за разложения электролита. 16,17,28,151 Для анализа таких элементов очень важны подготовка образцов и промывка электродов, как описано в разделе «Методы вскрытия ячеек».Однако можно обратить внимание, что в некоторых случаях наличие таких элементов действительно может быть частью активного материала. 152
Krämer et al. модифицированные аноды с осаждением Li с использованием изопропанола. 150 EDX-картирование позволило обнаружить O и C, что предполагает образование Li 2 CO 3 , однако авторам пришлось провести дальнейшие измерения с использованием FTIR и XRD для проверки. 150 Модификация осаждения Li изопропанолом позволила оценить площадь на поверхности анода, покрытую Li 2 CO 3 , по EDX-картированию. 150
Maleki et al. исследовали эффекты глубокого разряда ниже напряжения конца разряда для коммерческих элементов LiCoO 2 / графит. 153 Авторы обнаружили, что разряд до 0 В может привести к растворению Cu из коллектора отрицательного тока, что, соответственно, было обнаружено как на аноде, так и на катоде с помощью EDX. 153 Кроме того, EDX использовался в сочетании с SEM для обнаружения загрязнений после отказа клеток. 54
Рентгеновская фотоэлектронная спектроскопия
Рентгеновская фотоэлектронная спектроскопия (XPS) основана на фотоэлектрическом эффекте. 98,154 Атомы в образце ионизируются рентгеновскими лучами, и измеряется кинетическая энергия испускаемых фотоэлектронов (рис. 6e). 98 Поскольку кинетическая энергия испускаемого фотоэлектрона характерна для его исходного элемента, XPS позволяет анализировать и определять все элементы (кроме H и He), их степени окисления и — в определенной степени — их химическое окружение. 98 XPS чувствителен к поверхности из-за малой длины свободного пробега испускаемых электронов в твердых телах (несколько нм). 98 Таким образом, XPS может характеризовать химические изменения на поверхности частиц, что делает его ценным с точки зрения посмертного анализа.
В лабораторных условиях обычно используются источники рентгеновского излучения из Al K-альфа. Кроме того, также можно использовать синхротронное излучение для проведения экспериментов в области жесткого рентгеновского излучения (HAXPES), 17,155,156 , однако это требует гораздо больших усилий.
Однако из-за высокого содержания энергии рентгеновского излучения необходимо учитывать возможность повреждения образца облучением.В частности, компоненты SEI могут изменить свою химическую природу. Следовательно, интерпретация данных XPS требует высокого уровня знаний исследуемой системы. Кроме того, следует упомянуть, что измерения XPS сильно локализованы, что делает необходимым зондирование большего образца в разных областях, чтобы получить обзор. XPS можно комбинировать с ионным распылением для получения профилей глубины. Неровная поверхность электрода делает это упражнение особенно трудным, и при анализе данных требуется особая осторожность.Однако в сочетании с распылением XPS не может измерять профиль глубины по всему образцу электрода. В частности, XPS ограничен первыми нанометрами поверхности. Следовательно, можно наблюдать только слой SEI, и часто сигнал от активного материала остается скрытым.
Полезный обзор возможных каталитических реакций, происходящих на границе раздела электролит-графит, и их наблюдение с помощью XPS недавно сделал Росс. 157 Общий обзор анализа SEI, включая XPS, был предоставлен Verma et al. 158
Коммерческие LiFePO 4 / графитовые ячейки, исследованные Klett et al. показали неравномерное старение электродов для циклических ячеек, тогда как электроды для календарных ячеек были однородными 17 , что связано с температурой 77,159–161 и градиентами давления, возникающими во время циклирования.
Lu et al. обсудили старение с акцентом на LiCoO 2 / графитовые элементы. 162 Авторы выполнили анализ профиля поверхности и глубины с помощью XPS и наблюдали увеличение толщины SEI в старых ячейках. 162 Недавний отчет группы Эренберга посвящен формированию SEI в коммерческих мешочках. 163 С помощью XPS авторы смогли идентифицировать составляющие внешнего и внутреннего слоев SEI, однако не удалось выяснить, есть ли какие-либо различия в характеристиках SEI для различных процедур формирования. 163
Несколько групп собрали информацию о составе SEI анодных материалов, отличных от графита, таких как SiO 164 или Sn. 47 Несколько авторов сообщили о переходных металлах, которые растворились с катода, мигрировали через электролит и выпали в осадок или были включены в слой SEI состаренного анода. 165,166 О таком поведении также сообщалось с использованием дополнительных методов. 16,18,26,28
Связь между различными условиями старения и химическим составом SEI представляла интерес для Zheng et al. 167 Авторы исследовали деградацию коммерческих ячеек LiFePO 4 / графит во время календарного старения в течение 10 месяцев при различных температурах и SOC. 167 Для температур, повышенных до 55 ° C, и SOC с большим накоплением, они наблюдали значительное увеличение объемного сопротивления и сопротивления переносу заряда, а также потерю емкости. 167 Посмертный XPS-анализ подтвердил, что вновь сформированные слои Li 2 CO 3 и LiF на поверхности анода были ответственны за изменения в поведении ячейки. 167
При разработке новых электродных материалов Post-Mortem XPS может помочь идентифицировать неизвестные продукты побочных реакций, происходящих на поверхностях.Как указано Феном и др., Основная проблема Li-S ячеек — это накопление S-частиц на поверхности электродов, обнаруживаемое XPS, и, как следствие, снижение емкости. 168
XPS — универсальный инструмент для получения информации о химическом составе поверхностных частиц, образующихся в процессе старения. В отличие от EDX, ЯМР, XRD и ИК-спектроскопии, почти все элементы могут быть полуколичественно обнаружены с помощью XPS. Кроме того, XPS — один из немногих методов изучения продуктов реакции образования и разложения SEI.
Инфракрасная спектроскопия с преобразованием Фурье
Инфракрасная спектроскопия с преобразованием Фурье (FTIR) основана на взаимодействии образца с инфракрасным излучением. На рисунке 6h показано это взаимодействие в режиме отражения. Отметим, что режим передачи также возможен для FTIR, который является массовым методом и поэтому здесь не обсуждается. Данные высокого разрешения собираются одновременно в выбранном спектральном диапазоне. Реальный спектр создается путем применения преобразования Фурье к сигналу.
Ранние FTIR-исследования материалов в литий-ионных элементах были проведены группой Аурбаха 169 170 и Йошидой и др. 171 и сосредоточена на понимании химических характеристик SEI на анодах на основе лития и графита. Эти исследования позволили идентифицировать важные полосы отражений SEI как асимметричное удлинение карбонила при 1650 см −1 , характерное для (ROCO 2 Li) 2 и 1450 и 870 см −1 , характерное для Li 2 CO. 3 .
FTIR-исследования образцов, полученных в результате анализа Post-Mortem, также были проведены с целью устранения различий при использовании добавок к электролитам. 172–174 В этих случаях исследовались как аноды, так и катоды. Аналогичным образом, результаты FTIR используются для сравнения характеристик SEI при замене соли на основе Li. 175
Многие другие исследования FTIR были проведены для отслеживания эффектов старения. 34,46,150,176,177 Aurbach et al. провели испытания FTIR на образцах электродов на основе графита и LiCoO 2 из литий-ионных ячеек 18650, циклически проверенных при различных температурах. 34 Керлау и Костецки проанализировали Li 0,8 Ni 0,8 Co 0,15 Al 0,05 O 2 электродов на основе и углерода после календарного старения при 45 ° C с помощью FTIR. 176 Электроды были промыты и высушены перед экспериментами, и в обоих случаях были обнаружены очень похожие спектры с полосами при 864 см -1 , 1008 см -1 и 1240 см -1 , присвоенных Li x PF y и Li x PF y O z , которые получены в результате термического разложения LiPF 6 . 176 Norberg et al. исследовали LiNi 0,5 Mn 1,5 O 4 катода на основе , циклированные с 1 M LiPF 6 в смеси EC / DEC. 177 После циклирования тесты FTIR выявили характеристические полосы алкилкарбонатов наряду с полосами при 1310 см −1 и 1110 см −1 , относящимися к модам растяжения CO и CC в кетонах, что свидетельствует о разложении электролита на поверхности катода. . 177 Однако идентификация конкретных соединений разложения с помощью этого метода была невозможна.
Следует отметить, что для экспериментов FTIR решающее значение имеет протокол подготовки образца электрода, поскольку следы электролита должны быть удалены, чтобы избежать нежелательных отражений. В качестве альтернативы измерению коэффициента отражения электродов также можно соскрести активный материал и построить таблетки KBr. 177,178 Транспортировка образца из перчаточного бокса в устройство FTIR, а также анализы должны выполняться в инертной атмосфере, поскольку (ROCO 2 Li) 2 на поверхности электродов может реагировать с H 2 O для образования Li 2 CO 3 . 179
Наконец, важно отметить, что FTIR не позволяет количественно определять соединения. Поэтому интерпретация результатов FTIR часто дополняет посмертный анализ другими методами, такими как электрохимическое тестирование, XPS и SEM / EDX.
Масс-спектроскопия вторичных ионов
Масс-спектроскопия вторичных ионов (SIMS) позволяет охарактеризовать элементный и молекулярный состав поверхности материала. Молекулярные фрагменты, кластеры, а также положительные и отрицательные ионы отрываются от поверхности с помощью первичного (импульсного) ионного пучка (рис. 6i).В случае статической SIMS или TOF-SIMS (Time of Flight SIMS) вторичные ионы, поступающие из образца, собираются и анализируются с помощью масс-анализатора «время пролета»: массовое разделение ионизированных фрагментов основывается на необходимом времени. чтобы добраться до детектора. Хотя количественную оценку применить сложно, ВИМС и, в частности, TOF-SIMS являются очень чувствительными методами (вплоть до нескольких частей на миллион). Кроме того, благодаря сложной электронике можно сфокусировать первичный ионный пучок и получить спектроскопическое изображение поверхностей.
Несмотря на то, что TOF-SIMS является поверхностно-чувствительным методом, он широко используется для исследования поверхности объемных материалов. Это делается путем распыления образца пучком ионов Cs + или Ar + , что позволяет получать профили массовой концентрации по глубине. Это делает TOF-SIMS мощным инструментом для определения характеристик тонких слоев, таких как те, которые используются в системах с микро-батареями. 180,181 Спектроскопия поверхности может помочь определить природу электрохимических пассивирующих слоев или покрытий на коллекторах и материалах электродов.
Использование TOF-SIMS также может помочь в изучении старения токосъемников или материалов электродов во время. 182–185 Wang et al. продемонстрировали растворение Fe в материале LiFePO 4 и подчеркнули роль защитного углеродного покрытия. 186,187 Аналогичным образом, растворение Mn и Ni из высоковольтной шпинели охарактеризовано путем комбинирования экспериментов XPS, TEM и TOF-SIMS. 188 Результаты TOF-SIMS, показывающие профили концентрации продуктов ионизации (LiF 2 — , MnF 3 — , NiF 3 — ), полученные в ходе анализа, позволяют сделать вывод о том, что наличие MnF 2 на поверхности катода.
SEI можно также изучить с помощью TOF-SIMS. На протяжении многих лет было доказано, что это мощный дополнительный подход к XPS, позволяющий лучше понять химическую структуру SEI. Первые исследования на основе SIMS в отношении материалов литий-ионных аккумуляторов возникли в 2000-х годах. Пелед и др. инициировал первые попытки изучения SEI на поверхности электрода с помощью TOF-SIMS на монокристаллах ВОПГ, поскольку этот материал можно рассматривать как модельный электрод для графитовых систем. 189,190 Авторы представили доказательства присутствия полимеров в SEI и зависимости химического состава SEI от природы плоскостей ВОПГ.Затем другие группы рассмотрели TOF-SIMS-спектроскопию для изучения влияния добавок или альтернативных электролитов (например, ионных жидкостей) на химическую структуру SEI. 191–194
Хотя TOF-SIMS все еще недостаточно используется в области накопления энергии, в частности, для приложений с литий-ионными аккумуляторами, количество таких исследований за последние годы выросло. Уникальными сильными сторонами этого мощного метода спектроскопии анализа поверхности являются чувствительность, способность анализировать изотопы, лучшее разрешение по горизонтали по сравнению с другими методами спектроскопии анализа поверхности, такими как XPS.
Оптическая эмиссионная спектроскопия тлеющего разряда по глубине
Глубинная оптическая эмиссионная спектроскопия тлеющего разряда (GD-OES) обеспечивает элементный анализ образцов путем распыления и обнаружения испускаемого видимого света от оторвавшихся частиц, которые возбуждаются в плазме Гримма лампа 195 (Рис. 6g) и обнаруживается спектрометром с круговой диаграммой Роуленда. 100
GD-OES хорошо зарекомендовал себя для контроля качества обработки поверхности и стальных покрытий с использованием потенциала постоянного тока (DC).Благодаря простоте использования и высокой чувствительности, были проведены дальнейшие разработки, чтобы сделать этот метод применимым к непроводящему материалу с помощью радиочастотного (RF) потенциала, что позволило распространить применение GD-OES на тонкие пленки и непроводящие материалы. -анализ проводящих покрытий. 196–198 Совсем недавно электроды литий-ионных аккумуляторов стали предметом исследований GD-OES. 26,28,43,44,72,199,200
Данные профиля глубины достигаются путем послойного удаления атомов образца с использованием плазменного распыления. 201 Установка выглядит следующим образом: 195 образец помещается перед анодом и играет роль катода. Анод устройства GD-OES представляет собой полый цилиндр, который будет заполнен газообразным аргоном низкого давления (~ 10 -4 гПа). Ионизация газа и генерация плазмы достигается при приложении разности потенциалов (~ 500–1000 В). Распыленные атомы образца диффундируют в плазму и возбуждаются в результате дальнейших столкновений. Это приводит к испусканию характеристических фотонов, которые будут регистрироваться оптическим эмиссионным спектрометром.Цилиндрический анод имеет типичный диаметр 2,5 мм или 4,0 мм, что соответствует размеру пятна анализа. Конструкция лампы Гримма делает анализ GD-OES независимым от матрицы образца. 100
В отличие от ряда других методов, таких как XPS и SIMS, глубинное профилирование GD-OES не ограничивается близостью поверхности образца, но может анализировать его от поверхности электрода до токосъемника. Таким образом, GD-OES может давать информацию как о поверхности электродов, так и об объеме электрода. 26,28,43,44,72,199,200
Saito et al. наблюдали распределение Li в катодах от мощных литий-ионных элементов. 199 Авторы сообщили о градиенте Li по направлению к поверхности для разряженного состояния и наоборот, что было приписано медленной диффузии Li как в электроде, так и в электролите. 199 Al-дефицитные области в NCA, как также сообщалось, образовались во время езды на велосипеде. 199 Takahara et al. провели обширные исследования не только катодов, но и анодов на основе графита, уделяя особое внимание росту SEI при циклическом старении. 26,43,44,200 Авторам удалось выполнить калибровку на основе конкретного случая исследования и добиться количественного распределения Li по графитовому аноду. Они также сообщили о более быстром и более точном профилировании анодов на основе графита по глубине с использованием газа Ar с дополнительным 1% H 2 . 200
GD-OES был применен к графитовым электродам от старых коммерческих ячеек 18650, где была достигнута корреляция с электрохимическими данными. 28 В исследовании проводилось различие между «поверхностным» и «объемным» Li с использованием данных глубинного профилирования GD-OES, откалиброванных для Li на основе результатов ICP-OES. 28 Исследование показало, что содержание Li на поверхности коррелирует с величиной потери емкости, что подразумевает важную роль побочных реакций на поверхности графита в деградации клеток. 28
GD-OES недавно был использован в посмертном анализе для обнаружения лития на графитовых анодах, 72 , что затруднительно или невозможно другими методами. По сравнению с анодами с SEI градиент Li и содержание Li значительно увеличиваются в случае гальванического покрытия Li. 72 Кроме того, было замечено, что большая часть металлического Li размещается на поверхности графитовых анодов, 72 , что согласуется с расчетами Хейна и Латца. 202
Из-за сравнительно короткого времени измерения, небольшого размера образца, высокой чувствительности и его возможности обнаруживать Li в профилях глубины через все электроды, метод GD-OES является многообещающим аналитическим инструментом для лучшего понимания Li — механизмы старения ионных аккумуляторов. Однако, чтобы получить полную картину механизмов старения, GD-OES необходимо сочетать с дополнительными методами.
Методы химического анализа для анализа объема электродов
Оптико-эмиссионная спектрометрия с индуктивно связанной плазмой
В посмертных анализах для определения элементного состава электродов используется оптическая эмиссионная спектрометрия с индуктивно связанной плазмой (ICP-OES). 16,18,19,28,151 В ИСП-ОЭС индуктивно связанная плазма используется для получения возбужденных ионов и атомов из образца, которые испускают электромагнитное излучение в видимом диапазоне (рис. 6f).Длины волн этого излучения характерны для конкретного элемента. Таким образом, ИСП-ОЭС может определить соотношение между элементами, присутствующими в образце. Преимущество этого метода заключается в том, что могут быть обнаружены элементы от диапазона ppm до основных элементов образца. Однако одним из недостатков является то, что ИСП-ОЭС не дает полного состава пробы, что требует использования дополнительных методов. ICP-OES часто сравнивают с EDX (см. Выше), однако ICP-OES имеет преимущество в обнаружении Li.Образцы полностью растворяются в кислотном растворе, а затем измеряются. Это означает, что исследованию подлежит не только поверхность, а большая часть образца. Кроме того, для ИСП-ОЭС требуются площади образца в диапазоне 2 см. Однако ICP-OES не может обеспечить профили по глубине, и материал приходится соскабливать с нескольких сантиметров 2 образцов электродов, что ограничивает его способность изучать местные явления.
В посмертном анализе полезны измерения ICP-OES для подтверждения растворения переходных металлов с катода путем обнаружения перемещенного материала на аноде. 16,18,19,28,151 Было показано, что это растворение способствует механизму старения анода 16,19,28,36,37 и вызывается HF. 36,37 Stiaszny et al. обнаружены концентрации переходных металлов в свежих и состаренных анодах LiMn 2 O 4 -NMC / графитовых ячеек с помощью ICP-OES. 19 Этот результат был подтвержден уменьшением высоты пика NMC в циклической вольтамперометрии. 19 Было обнаружено, что количество Mn на графитовых анодах, растворенных из смеси NMC / LiMn 2 O 4 катодов, увеличивается с температурой 16,28 и временем 28 с помощью ICP-OES.Klein et al. хранится LiFe 0,3 Mn 0,7 PO 4 / LiMn 1,9 Al 0,1 O 4 смешанных катодов с различными соотношениями в электролите в течение двух недель при 60 ° C. 203 Авторы наблюдали наименьшее растворение Mn в чистом оливине, тогда как оно было на два порядка выше для чистой шпинели. 203 Для всех смесей авторы обнаружили резко сниженное количество растворения Mn в электролите с помощью ICP-OES. 203
Рост толщины SEI является еще одним механизмом старения и был изучен с помощью ICP-OES, ограниченного элементами Li, P и Mn и поддерживаемого EDX. 16,28 Результаты согласуются с ростом SEI за счет разложения соли LiPF 6 на аноде | электролит интерфейс. 16,28,33 Было обнаружено, что потребление циклического Li на аноде, измеренное методом ICP-OES, напрямую коррелирует с уменьшением емкости, 12,28 — уменьшением Li в катоде и увеличением сопротивления элемента. 28 С другой стороны, обсуждается реакция электролита на поверхности анода, приводящая к высыханию элементов, что приводит к дальнейшему снижению емкости. 25,204
Спектроскопия ядерного магнитного резонанса
Спектроскопия ядерного магнитного резонанса (ЯМР) — мощный метод, позволяющий охарактеризовать материалы и химические соединения в твердом состоянии и разбавленных растворителями. 205,206 Он предоставляет не только химическую и структурную информацию, но также информацию о транспортных свойствах и подвижности ионов, электронных, магнитных, а также термодинамических и кинетических свойствах. 207–211
Образец помещают в магнитное поле и возбуждают радиочастотным импульсом (рис. 6j).Записанный спад свободной индукции (FID) обрабатывается с помощью преобразования Фурье для получения спектра ЯМР. Требуется одно или несколько ЯМР-активных ядер (ядерный спин 0), которые служат в качестве зонда для обнаружения изменений в их химическом окружении и их электронных свойствах. Оба типа образцов, жидких и твердых, также могут быть исследованы на месте с использованием специальных измерительных установок. 212 213 Несколько авторов представили обзорные статьи, касающиеся ЯМР-спектроскопии при разработке литий-ионных клеток и посмертном анализе. 214
ЯМР — полезный инструмент для облегчения разработки новых анодных материалов, основанных на результатах анализа посмертных исследований. Согласно Delpuech et al., Высокая необратимая потеря емкости анодов на основе Si в основном происходит из-за разложения карбонатных растворителей с последующим образованием нелитированных углеродных частиц в олигомерной или полимерной форме. 215 Грей и его сотрудники показали, что потеря емкости и саморазряд напрямую связаны со структурными изменениями кремниевых анодов, и их можно избежать путем правильного выбора связующих. 216 Перес-Висенте и его коллеги изучили Sn 4 P 3 как возможный новый анодный материал. 217
Хотя большинство исследований ЯМР, связанных с материалами литий-ионных элементов, состоят из измерений твердотельного вращения под магическим углом (MAS), в сообществе литий-ионных элементов часто игнорируется, что это также мощный метод исследования жидкости. образцы и решения. Современные жидкие электролиты, представляющие собой смеси огромного количества органических и неорганических соединений, обеспечивают большое количество ЯМР-активных ядер, таких как 1 H, 13 C (в органических молекулах), 7 Li, 31 P, 19 F (в LiPF 6 ), в случае недавно разработанных проводящих солей, таких как LiTFSI или LiFSI, даже 14 N или 15 N могут быть вариантами.Это огромное разнообразие активных ядер открывает множество возможностей для изучения явлений, связанных с электролитом, таких как старение, разложение электролита или образование SEI.
DeSilva et al. исследовали формирование SEI на LiNi 0,80 Co 0,2 O 2 катодах и MCMB- (1028) -углеродных анодах с помощью твердотельного MAS ЯМР 7 Li, 19 F и 31 P. 218 Их электролиты на основе LiPF 6 представляли собой растворители на основе карбонатов (EC, EMC), содержащие фторированные добавки, такие как 1-FEC, DTFEC (бис- (2,2,2-трифторэтилкарбонат), 2,2,2-трифторэтил метилкарбонат (TFEMC) и трифенилфосфат. 218 В случае катода авторы могли определить различные количества необратимых концентраций Li для одного и того же номинального электрохимического SOC после разборки ячейки. 218 Кроме того, они могли показать, что аддитивное разложение и осаждение также происходят на катоде. 218
С анодной точки зрения были обнаружены различные количества LiF, а также продукты разложения фторированных карбонатов. Дюпре с сотрудниками исследовали состаренные электроды из Li 4 Ti 5 O 12 и LiFePO 4 и определили их концентрации LiF. 219 Путем корреляции со снижением емкости циклического элемента, авторы смогли предложить несколько путей реакции для различных механизмов старения исследуемых электродов. 219
Лухт и его сотрудники предложили механизмы термического разложения из-за автокатализа и протонных примесей для нескольких карбонатных растворителей, часто используемых в современных электролитах, таких как DMC, EC и DEC. 220,221 Их исследование представляло собой комбинацию методов ЯМР ( 1 H, 13 C, 19 F, 31 P, DEPT, COSY и HETCOR), GC-MS и SEC и проводилось на модели. система, которая не содержала материала, собранного из старого литий-ионного элемента.Однако, помимо протонных примесей, таких как H 2 O или этанол, авторы смогли идентифицировать DEC как главную причину термического разложения LiPF 6 и наблюдали ряд продуктов разложения, которые также могут изменять материалы анода и катода во время жизнь литий-ионного элемента. 220 В ходе последующего исследования они исследовали взаимодействие нескольких катодных материалов с органическими электролитами при повышенных температурах. 221 Очевидно, Li 2 CO 3 , присутствующий на поверхности катодных частиц, способен ингибировать термическое разложение органических электролитов. 221 Кроме того, продукты разложения электролита, обнаруженные в этом исследовании, были аналогичны тем, которые наблюдались на катодах, удаленных из подвергшихся термическому воздействию литий-ионных элементов. 222
Подводя итог, можно сказать, что модельные исследования, включающие измерения ЯМР в сочетании с другими методами определения характеристик, могут быть полезны для интерпретации данных, полученных в результате посмертного анализа старых литий-ионных клеток.
Структурная характеристика
В посмертном анализе метод дифракции рентгеновских лучей (XRD) обычно применяется для структурного анализа активных материалов в электродах (рис. 6k).Как и все дифракционные методы, XRD применим только к материалам, атомы которых обладают определенной периодичностью. XRD широко используется при диагностике многих механизмов старения, поскольку он предоставляет важную информацию о структурных изменениях, которые кристаллические активные материалы могут претерпеть во время старения. 16,34,81,135,223
Кроме того, XRD предоставляет информацию об изменениях ориентации частиц и образовании пленки на электроде | электролит интерфейс. 34,135 Последнее явление можно увидеть на XRD по уменьшению интенсивности пика. 34,135 Аналогичным образом, Liu et al. показали, что уширение пиков XRD указывает на возникновение расслоения графита. 223 Кристаллические продукты разложения на поверхности анода наблюдаются дополнительными пиками. 122
XRD также позволяет обнаруживать химические реакции разложения / растворения по уменьшению объема решетки катодных частиц. Stiaszny et al. проанализировали коммерческую литий-ионную батарею со смешанным катодом LiMn 2 O 4 / NMC и графитовым анодом, циклически повторяемым при комнатной температуре. 18 Авторы измерили изменение параметров решетки состаренного активного материала NMC, на которое повлияло растворение переходных металлов в электролите, что вызывает снижение количества Li в катоде. 18,19
Зависимое от температуры изменение механизма старения (покрытие Li / рост SEI) в коммерческой ячейке 18650 с графитовым анодом и LiMn 2 O 4 / NMC-катод было видно в измерениях XRD по изменению постоянных решетки a и c NMC. 16 Константы решетки также коррелировали с содержанием Li, измеренным методом ICP-OES в состаренных анодах и катодах. 16 После сохранения осаждения Li путем химической реакции его поверхности с изопропанолом для идентификации Li 2 CO 3 использовали порошковую дифракцию рентгеновских лучей (PXRD). 150 Результаты согласуются с измерениями EDX и FTIR. 150
XRD — это распространенный метод определения постоянных решетки кристаллических активных материалов.Однако, поскольку XRD ограничивается измерениями в объеме электрода, его часто приходится комбинировать с другими методами, например поверхностно-чувствительный метод, такой как SEM или EDX.
Анализ электролита
Разложение электролита происходит из-за побочных реакций, которые приводят к образованию нерастворимых, растворимых и газообразных продуктов. 37,224,225 Идентификация таких продуктов имеет основополагающее значение для отслеживания побочных реакций, ответственных за старение батареи. Таким образом, во многих исследованиях реализованы методы анализа электролитов и газов, образующихся при старении батареи.Наряду с жидкостной ЯМР-спектроскопией (см. Выше) хроматографические методы оказались очень успешными для посмертной характеристики электролитов. Основной принцип хроматографии — разделение компонентов смеси и последующее обнаружение. Разделение, например, достигается за счет разного времени удерживания в адсорбированном состоянии на стенке при прохождении через капилляр (рис. 6l). Эксперименты с другими методами часто дополняют друг друга, особенно для анализа нерастворимых продуктов; эти методы описаны в других подразделах этой рукописи.Большинство исследований, касающихся методов хроматографии, сосредоточено на смесях растворителей на основе карбоната LiPF 6 (EC, PC, DMC, EMC и DEC), поскольку это наиболее распространенные электролиты, используемые в литий-ионных батареях.
Далее обзор литературы по посмертному анализу электролита и газа разделен на две основные группы: исследования с лабораторными ячейками, разработанными специально для сбора большого количества проб жидкости и газа, и исследования с коммерческими ячейками, обычно содержащими только небольшой избыток электролита, который трудно восстановить.
Анализ электролита и газа, выполненный на ячейках лабораторного масштаба
Ранние исследования 171 были выполнены после первой загрузки LiCoO 2 / графитовых элементов. Авторы применили жидкостную хроматографию в сочетании с инфракрасной спектрометрией Фурье (LC-FTIR) для анализа восстановленного электролита, который предоставил алкилдикарбонаты в качестве основного растворимого соединения, образующегося при разложении электролита. 171
Газовая хроматография (ГХ) в сочетании с детектором теплопроводности (GC-TCD) позволила наблюдать H 2 , CO, CO 2 , CH 4 , C 2 H 4 , C 2 H 6 и C 3 H 8 при первой зарядке.Чтобы понять механизм образования алкилдикарбонатов, Sasaki et al. провели посмертный анализ ГХ-масс-спектрометрии (ГХ-МС) электролита, извлеченного после циклирования литий / графитовых полуэлементов. 226 Присутствие алкилдикарбоната было подтверждено, и параллельное химическое моделирование показало, что алкоксиды Li могут запускать образование алкилдикарбоната. 226
Стремясь понять восстановительное разложение LiPF 6 -карбонатных растворителей, группа Ларуэля 227–229 провела посмертный анализ электролита и газа на ячейках лабораторного масштаба.Авторы использовали ионизацию электрораспылением в сочетании с масс-спектрометрией высокого разрешения (ESI-HRMS) и GC-MS для анализа электролитов, хотя другой набор капиллярных колонок был реализован для GC-MS анализов задержанного газа. Параллельное использование этих методов позволяет обнаруживать соединения в широком диапазоне масс, таким образом, был выяснен общий механизм разложения электролита. 228,229 Авторы обнаружили, что большинство соединений разложения, полученных в результате линейного восстановления карбоната, обеспечивают алкоксиды Li, которые дополнительно запускают этерификацию электролита, в то время как двухступенчатое восстановление EC было менее важным.
С другой стороны, следы H 2 O в LiPF 6 приводят к образованию POF 3 , HF и LiF. Этот механизм усиливается за счет температуры, следовательно, в литературе также можно найти тесты хранения электролита с использованием хроматографических анализов. 230–234 Terborg et al. 230 исследовали механизмы термического старения и гидролиза LiPF 6 с помощью ионной хроматографии (IC) в сочетании с ESI-MS. В этих исследованиях интересно отметить, что реализация IC позволяет обнаруживать HF в электролитах благодаря идентификации F — .Kraft et al. 234 изучили продукты разложения электролитов LP30 и LP50 при термическом старении, разработав методы разделения и сравнив надежность трех различных колонок IC. Более того, авторы объединили IC-ESI-MS-MS для идентификации новых фосфорорганических соединений. 234 Handel et al. исследовали термическое разложение смесей EC / DEC + LiPF 6 с загрязнением деионизированной водой, применяя ГХ-МС для анализов жидких электролитов и ГХ-МС над паром для анализов летучих соединений. 233 Также были выполнены дополнительный ЯМР и кислотное титрование. Авторы пришли к выводу, что старение электролита протекает с низкой скоростью, поскольку исключаются каталитические поверхности, окружающий воздух и протонные примеси. 233
Анализ электролита и газа, выполняемый на коммерческих элементах
Хроматографические методы также применялись для анализа электролита и газа, взятого из коммерческих литий-ионных аккумуляторов. 83,225,235–238 Задача состоит в том, чтобы применить знания, полученные в результате анализов на лабораторных ячейках, на коммерческих, чтобы указать пути разложения электролита, соблюдаемые в соответствии с заданным протоколом старения.Идентификация нежелательных продуктов реакции может указать на будущую оптимизацию коммерческих ячеек. Тем не менее, метод отбора проб имеет решающее значение, поскольку коммерческие ячейки обычно не имеют ни избытка электролита, ни газовых карманов.
Kumai et al. спроектировал сосуд «газовыделения» для отбора проб газа из графитовых ячеек LiCoO 2 после циклических испытаний, перезарядки и избыточного разряда. 235 CO 2 , CO, CH 4 , C 2 H 6 и C 3 H 8 были обнаружены путем проведения анализов GC-TCD и GC в сочетании с пламенно-ионизационным детектором (GC -FID).Terborg et al. восстановил электролит из промышленной ячейки после 1400 циклов промывкой сепараторов, анода и катодов в ПК. 237 Затем растворы подвергали анализу ГХ-МС. 237
Совсем недавно было представлено оборудование GC-FTIR-MS 236 как полезный метод для анализа газов из набухшей коммерческой ячейки. Улавливание газа проводили в перчаточном боксе, наполненном аргоном, путем прокалывания мешка для ячеек с герметичным шприцем. Авторы идентифицировали CO, CO 2 , CH 4 и C 3 H 8 на графике ГХ / FTIR Грама-Шмидта, тогда как хроматограмма ГХ / МС позволила обнаружить другие менее распространенные летучие соединения. 236 Дополнительные анализы электролитов с помощью ГХ-МС позволили обнаружить алкилдикарбонаты и более длинные карбонатные цепи, а также органофосфатные соединения, что указывает на то, что следы воды, присутствующие в коммерческой ячейке, играют роль в разложении электролита. 236
Grützke et al. восстановленный электролит из литий-ионных элементов на основе NMC 5 Ач, которые были испытаны в полевых условиях в HEV. 83 Клапан давления каждой ячейки был раздавлен, и электролит был собран. 83 Анализ ГХ-МС выявил компоненты электролита, а анализ ГХ-ПИД позволил оценить состав. 83 Авторы обнаружили виды F — и PO 2 F 2 — с помощью IC-ESI-MS при открытии ячеек в инертной атмосфере, тогда как HPO 3 F — и H 2 PO 4 — также были обнаружены при открытии во влажной среде. 83 В промышленных ячейках следы воды кажутся неизбежными из-за гигроскопичности LiPF 6 , тогда электрохимическое / химическое разложение электролита, которое приводит к этерификации растворителя, сопровождается образованием органофосфатов, поскольку продукты разложения реагируют с POF 3 . 228
Чтобы понять влияние добавок, необходимо провести посмертный анализ разложившихся электролитов и образовавшегося газа на ячейках 239 лабораторного масштаба, а также на более крупных прототипах. 225 Более того, методы посмертной хроматографии начинают применяться для оценки новых составов электролитов, предназначенных для приложений высокого напряжения, для которых окисление электролита и термическое разложение являются основными проблемами, которые необходимо решить.
Электрохимический анализ повторно собранных электродов
Реконструкция в полуэлементы
Посмертные электрохимические характеристики могут быть выполнены в ячейках путем реконструкции анодов или катодов вместе с металлическим Li в качестве противоэлектрода. 12,17–19,34,88 Для электродов с двусторонним покрытием необходимо удалить одно покрытие, например с помощью N-метилпирролидона 88 или с помощью лазерного гашения. 87,90
Электроды, извлеченные из свежих и старых клеток, изучаются по одним и тем же протоколам. Целями этих электрохимических испытаний являются (i) определение остаточной (или остаточной) емкости электродов (в мАч / см 2 ) и (ii) измерение обратимой емкости (в мАч / см 2 ), учитывая что элементы обязательно были разобраны на одном и том же SOC (часто 0% от SOC).
Для отрицательного электрода / литиевых ячеек первый электрохимический тест заключается в заряде для извлечения лития из анода (делитирование). В то время как для положительного электрода / литиевых ячеек первые испытания заключаются в разряде для введения лития в катод (литиирование). Соответствующие емкости являются остаточными емкостями электродов. Для получения обратимых емкостей отрицательный электрод в полуэлементе снова литируется, а положительный электрод — литиирован.
В электродах, извлеченных из свежих ячеек, остаточная емкость позволяет оценить начальную необратимость ячейки из-за образования слоя SEI на этапе формирования. 46 Реверсивная мощность раскрывает начальную балансировку. 12
В состаренных электродах изменение остаточной емкости указывает на потребление Li в побочных реакциях, а изменение обратимой емкости служит для отслеживания дисбаланса клеток при старении, а также для правильного определения механизма старения. 17,46
На рисунке 8 показан принцип определения емкости в монетных половинках. Следует отметить, что следует соблюдать осторожность при использовании указанного метода определения емкости, если в электроде присутствуют неоднородности из-за ошибок изготовления или старения.Один из способов преодоления этой проблемы — не учитывать среднюю поверхностную емкость (в мАч / см 2 ) образцов, а учитывать средний вес образцов, собранных со «свежих» электродов перед созданием плоских ячеек для расчета и сравните массовую емкость (в мАч / г).
Увеличить Уменьшить Сбросить размер изображения
Рис. 8. Процедура электрохимических измерений в собранных монетных полуячейках.
Kobayashi et al. предложили аналогичную процедуру определения емкости каждого электрода. 12 Они изучили LiMn 2 O 4 / LiNi 0,8 Co 0,15 Al 0,05 O 2 смешанный катод по сравнению с . — анодная система из угольного графита. 12 Остаточная емкость катодов, собранная из элементов после циклического или календарного старения (преобразованная в SOC катода в состоянии разряда), увеличилась по сравнению со значением, полученным для свежего элемента. 12 Авторы продемонстрировали, что существует взаимосвязь между сохранением емкости исследуемой ячейки и SOC катода в состоянии разряда (идентично с напряжением холостого хода полукруглого элемента, которое определяет состояние литиирования). электрода). 12 Это говорит о том, что ионы Li не только необратимо накапливаются на анодной стороне в начальном цикле образования SEI, но также постоянно накапливаются во время циклического или календарного старения. Аналогичные результаты были получены другими методами. 16,28
Aurbach et al. повторно собрал свежие и повторно использованные электроды из ячейки 18650 с угольными анодами и катодами из LiCoO 2 в ячейки с литиевыми противоэлектродами и электродами сравнения. 34 Авторы выполнили циклическую вольтамперометрию (CV) и спектроскопию электрохимического импеданса (EIS) с этими повторно собранными ячейками. 34 Из измерений CV и EIS они обнаружили, что кинетика состаренных анодов замедляется по сравнению со свежими анодами. 34 Причина — рост толстой поверхностной пленки при старении. 28,34,135,240
Такие базовые электрохимические испытания в конфигурации полуячейки позволяют получить изображение реального состояния литиирования каждого электрода полной ячейки в состоянии разряда. Оставшиеся емкости могут косвенно привести к необратимому накоплению ионов Li в аноде, что может быть подтверждено также химическим анализом анода с помощью ICP-OES. Обратимые емкости позволяют оценить эволюцию вводимой способности основной структуры каждого электрода и определить наиболее значимый фактор замирания емкости.Ключевым вопросом является согласованность повторно собранных ячеек, поэтому по крайней мере две ячейки должны быть построены из одних и тех же электродов.
Реконструкция в полные ячейки с электродами сравнения
Дополнительно к реконструкции анода или катода в полуячейки по сравнению с . Li, 12,34,81 можно построить полные ячейки, используя анод, катод и дополнительный электрод сравнения (RE). 75,87,241 УЭ позволяет получать потенциалы как анода, так и катода во время зарядки и разрядки. 87 Отметим, что измерения в полуячейках дают другой результат, так как взаимодействие между анодом и катодом отсутствует.
Стабильность во времени потенциала RE имеет фундаментальное значение и зависит от температуры испытания и природы электрохимической пары, выбранной в качестве RE (Li + / Li, 16,241–257,90 FePO 4 / LiFePO 4 , 258 двухфазные пары, такие как Li 4 Ti 5 O 12 / Li 7 Ti 5 O 12 , 258–261 Сплавы Li, такие как Li x Sn / Sn, 262,263 Li x Al / Al, 264 или Li x Bi / Bi 265 ).
Как известно, позиция RE особенно важна. Например, в водных системах хорошо зарекомендовал себя капилляр Луггина-Габера, который расположен (i) рядом с рабочим электродом и (ii) между закупоривающим и противоэлектродом. 266 Недавно Hogg и Wohlfahrt-Mehrens выполнили измерения в 4-электродных полных ячейках с двумя RE. 241 Авторы обнаружили, что положение УЭ между анодом и катодом также очень важно для правильного измерения анодных потенциалов в полностью литий-ионной ячейке. 241
Ramadass et al. повторно собранные Т-клетки против . Li с графитовых анодов и катодов LiCoO 2 от промышленных ячеек 18650 (800 циклов при комнатной температуре). 81 Повторно собранные элементы были встроены в перчаточный ящик с использованием сепаратора от разобранного старого элемента Sony и 1 M LiPF 6 в EC: DMC = 1: 1 в качестве электролита. 81 Графитовый анод и катод NCA из коммерческих высокоэнергетических ячеек типа 18650 были недавно повторно собраны в 3-электродные полные ячейки с дополнительным литиевым электродом сравнения. 87 Используя этот метод, можно было измерить потенциал анода против . Li / Li + и, следовательно, для определения условий осаждения Li. 87,90 Следовательно, могут быть разработаны оптимизированные процедуры зарядки для предотвращения осаждения лития в коммерческих элементах 18650 и значительного увеличения срока службы батареи. 87
В дополнение к измерениям электродных потенциалов с помощью RE, также возможно выполнять измерения импеданса как анода, так и катода, извлеченных из свежих и состаренных ячеек, одновременно при различных состояниях заряда. 261 Этот тип измерения требует оптимизации морфологии УЭ и его размещения внутри ячейки для получения надежных спектров импеданса.
Расположение RE важно для получения надежных значений потенциала и импеданса. 248 Dees et al. смоделировали распределение потенциала электролита внутри ячейки, чтобы найти наилучшее расположение RE внутри ячейки-пакета. 267 В других статьях 248 249 268 показаны искажения или артефакты смоделированных спектров импеданса комбинированных геометрических и электрических асимметрий в электродах.Эти артефакты часто представляют собой индукционные петли в низкочастотной области одного из электродов.
В любом случае очень важно держать электроды в строго контролируемых условиях. 75 Itou et al. повторно собраны электроды из циклически проработанных ячеек вместе со свежими электродами в новые ячейки с литиевым электродом сравнения, чтобы измерить увеличение сопротивления электродов. 75 Этим методом авторы обнаружили, что катод в основном отвечает за увеличение сопротивления во время циклирования при 60 ° C. 75 Последовательно, FIB / SEM и XAFS выявили трещины на границах границ зерен внутри частиц и локальные изменения в катоде LiNi 0,8 Co 0,15 Al 0,05 O 2 соответственно. 75
Следует отметить, что еще одна конфигурация для посмертных испытаний импеданса представляет собой симметричные круглые ячейки с электродами той же полярности. Это позволяет оценивать каждый электрод без влияния противоэлектрода или RE. 269,270 Недостатком этой конфигурации является то, что полное сопротивление может быть получено только в одном уникальном состоянии литирования, соответствующем SOC элемента перед разборкой.
Альтернативным решением может быть интеграция RE непосредственно в все еще функционирующую промышленную ячейку без разборки и реконструкции электродов. Этот подход сложен, поскольку необходимо обеспечить надлежащее повторное запечатывание ячеек.
Как подробно описано в разделе «Физико-химический анализ состаренных материалов после разборки литий-ионных элементов», каждый метод физико-химического анализа имеет свои определенные преимущества и недостатки, позволяя наблюдать только определенные аспекты механизма старения, не имея возможности охарактеризовать других.Например, поверхностно-чувствительные методы не могут получить доступ к объемным свойствам электродов. Напротив, методы, чувствительные к массе, смешивают свойства поверхности со свойствами массы электрода. Поскольку объем обычно намного больше поверхности, влияние поверхности на измерение часто незначительно. Методы профилирования по глубине обнаруживают как поверхность электрода, так и объем, однако они не обнаруживают морфологических или структурных изменений.
Требуемые возможности метода анализа сильно зависят от наблюдаемого механизма старения.На рисунке 9 показан схематический обзор механизмов разрушения электродов и материалов, который часто не упоминается в литературе. Деградация электродов включает рост пленок на поверхности электродов (разложение электролита 16,18,26,28,40,122 или осаждение лития 25,45,72 ), засорение пор электродов или сепаратора, 37 отслоение электрода. сепаратор, 42,95 трещины в покрытии электрода 32,42,223 или деформация электродов или сепаратора. 9,14,32
Деградация материала включает трещины частиц, 42,74,75,129,130 отслаивание, 37 изменения на поверхности частиц, 37,158 образование пленки на частицах, 37,271 растворение / миграцию переходных металлов, 36,37,272 разложение электролита, 175,228,229 или закрытие пор сепаратора (например, под действием приложенного давления). 134 273 Соответствующие рекомендуемые методы анализа показаны на Рисунке 9.
На рисунке 10 показан обзор возможностей методов анализа, подробно обсуждаемых в разделе «Физико-химический анализ состаренных материалов после разборки литий-ионных элементов». Зеленый, оранжевый и красный цвета указывают на хорошую, ограниченную и отсутствие возможности обнаружения конкретного механизма старения соответственно. Из рисунка 10 ясно видно, что возможности различных методов анализа широко распространены, но нет метода, который бы охватывал все механизмы старения. Поэтому мы даем четкую рекомендацию исследовать образцы с помощью различных дополнительных методов анализа, чтобы получить полное представление о механизмах старения в литий-ионных элементах.
Увеличить Уменьшить Сбросить размер изображения
Рис. 10. Обзор методов анализа и явлений, которые они могут обнаружить. Зеленый, оранжевый и красный цвета указывают на хорошую, ограниченную и отсутствие возможности обнаружения конкретного механизма старения соответственно.
Разборка литий-ионных аккумуляторов является обязательной для сбора образцов для определения механизмов старения и улучшения материалов, включая поэтапное улучшение современных материалов, а также разработку новых поколений материалов.
В данной статье рассматриваются современные процедуры посмертного анализа старых литий-ионных клеток. В частности, подробно рассматриваются методы разборки старых литий-ионных элементов, а также физико-химический анализ их компонентов.
Химически инертная среда во время открытия ячейки имеет решающее значение для обеспечения надежных результатов с образцами, чувствительными к воздуху, и безопасной работы экспериментатора. Для последующей обработки образцов рекомендуется промыть электроды в растворителе, который уже входит в состав электролита (например,грамм. DMC) для поддержания качества образцов. Однако на данный момент не совсем ясно, как промывка влияет на слои SEI на электродах. Опытный экспериментатор, использующий соответствующее оборудование для вскрытия ячеек, является обязательным для получения интерпретируемых результатов при анализе образцов, полученных из литий-ионных ячеек.
Были рассмотрены доступные методы физико-химического анализа для посмертного анализа литий-ионных аккумуляторов, включая микроскопию, химические методы, чувствительные к поверхностям электродов и объему электродов, а также методы анализа электролитов и реконструкцию электродов на половину и полные ячейки с электродом сравнения.В последнем случае существует значительная разница между реконструкцией в половинные и полные ячейки. Полуячейки с анодами или катодом против . Противоэлектрод Li обеспечивает емкость отдельных электродов. Напротив, реконструкция анодов и катодов в 3-электродные ячейки с дополнительным электродом сравнения содержит информацию о взаимодействии между анодом и катодом. Таким образом, 3-электродные ячейки позволяют получить представление об сопротивлениях электродов и потенциалах электродов, которые имеют решающее значение для основных механизмов старения (например,грамм. Покрытие Li для отрицательных анодных потенциалов).
Каждый метод физико-химического анализа позволяет наблюдать только определенные аспекты разложения литий-ионных аккумуляторов. Поэтому рекомендуется исследовать образцы с помощью ряда дополнительных методов анализа, чтобы получить полную картину механизмов старения. Комбинируя рассмотренные методы, можно охарактеризовать все соответствующие части ячеек (аноды, катоды, сепараторы и электролиты) с точки зрения их микроструктуры, кристаллографической структуры и химического состава.
Только благодаря детальному знанию механизмов старения современные материалы могут быть удовлетворительно улучшены и разработаны новые материалы, отвечающие требованиям и требованиям будущих применений использования батарей в форме повышенной мощности и плотности энергии. Таким образом, мы считаем процедуры, рассмотренные в этой статье, подходящими для разборки элементов будущих поколений батарей с повышенной мощностью и плотностью энергии, после небольшого изменения метода разборки, если это необходимо.
Этот обзор был написан в рамках проекта MAT4BAT (http://mat4bat.eu/) и получил финансирование от Седьмой рамочной программы Европейского сообщества (FP7 / 2007-2013) в рамках грантового соглашения № 608931.
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно.Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались. Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом.Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie. Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу.Чтобы предоставить доступ без файлов cookie потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в cookie-файлах может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта.Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
Функции полисахаридов клеточной стенки в составе и архитектуре выявлены посредством мутаций
Akamatsu T, Hanzawa Y, Ohtake Y, Takahashi T, Nishitani K и Komeda Y 1999 Экспрессия генов эндоксилоглюкан трансферазы в acaulis мутантов 48 Arabidops.Plant Physiol. 121, 715–721.
Google Scholar
Arabidopsis Genome Initiative 2000 Анализ последовательности генома цветкового растения Arabidopsis thaliana . Nature 408, 796–815.
Google Scholar
Ариоли Т., Пенг Л.К., Бетцнер А.С., Берн Дж., Виттке В. и др. 1998 Молекулярный анализ биосинтеза целлюлозы в Arabidopsis .Science 279, 717–720.
Google Scholar
Баскин Т.И., Бетцнер А.С., Хоггарт Р., Корк А. и Уильямсон, Р.Е. 1992 Мутанты по морфологии корней у Arabidopsis thaliana . Aust. J. Plant Physiol. 19, 427–437.
Google Scholar
Бенфей П. Н., Линстед П. Дж., Робертс Р., Шифельбейн Дж. В., Хаузер М. Т. и Эшбахер Р. А. 1993 Развитие корней у четырех мутантов Arabidopsis с резко измененным морфогенезом корня.Развитие 119, 57–70.
Google Scholar
Bonin CP, Potter I, Vanzin GF и Reiter WD 1997 Ген MUR 1 Arabidopsis thaliana кодирует изоформу GDP-Dmannose-4,6-дегидратазы, катализируя первый шаг в de novo. синтез GDP-L-фукозы. Proc. Natl. Акад. Sci. США 94, 2085–2090.
Google Scholar
Браммелл Д. А., Катала С, Лэшбрук, С. С и Беннет А. Б. 1997 А, закрепленный на мембране эндо-типа Е эндо-1,4- ? -глюканаза локализуется на Гольджи и плазматических мембранах высших растений.Proc. Natl. Акад. Sci. США 94, 4794–4799.
Google Scholar
Burget E G и Reiter W-D 1999 Мутант mur 4 Arabidopsis частично дефектен в синтезе de novo уридиндифосфо L-арабинозы. Plant Physiol. 121, 383–389.
Google Scholar
Carpita N C 1996 Структура и биогенез клеточных стенок трав.Анну. Rev. Plant Physiol. Завод Мол. Биол. 47, 445–476.
Google Scholar
Карпита Н. С. и Гибео Д. М. 1993 Структурные модели первичных клеточных стенок у цветковых растений: соответствие молекулярной структуры физическим свойствам стенок во время роста. Плант Ж. 3, 1–30.
Google Scholar
Carpita N и Vergara C 1998 Рецепт получения целлюлозы.Science 279, 672–673.
Google Scholar
Чен Л-М, Карпита Н. С., Рейтер В. Д., Уилсон Р. Х., Джеффрис С. и Макканн М. С. 1998 Экспресс-метод скрининга мутантов клеточной стенки с использованием дискриминантного анализа инфракрасных спектров с преобразованием Фурье. Плант Ж. 16, 385–392.
Google Scholar
Dolezal O и Cobbett C. S 1991 дефицитный по арабинозокиназе мутант Arabidopsis thaliana .Plant Physiol. 96, 1255–1260.
Google Scholar
Doong R L и Mohnen D 1998 Солюбилизация и характеристика галактуронозилтрансферазы, которая синтезирует пектиновый полисахарид гомогалактуронан. Плант Ж. 13, 363–374.
Google Scholar
Ford C. W. и Hartley R D 1989 ГХ / МС определение характеристик циклодимеров из п-кумаровой и феруловой кислот с помощью фотодимеризации — возможного фактора, влияющего на биоразлагаемость клеточной стенки.J. Sci. Продовольственное сельское хозяйство. 46, 310.
Google Scholar
Fleischer A, Titel C и Ehwald R 1998 Потребность в боре и свойства клеточной стенки растущих и стационарных культивированных в суспензии клеток Chenopodium album L. Plant Physiol. 117, 1401–1410.
Google Scholar
Флеминг А. Дж., Маккуин Мейсон С., Мандель Т., Кюлемайер С. 1997 Индукция зачатков листьев путем экспансии белка клеточной стенки.Science 276, 1415–1418.
Google Scholar
Фрай С. С., Йорк В. С., Альберсхайм П., Дарвилл А., Хаяши Т. и др. 1993 г. Однозначная номенклатура олигосахаридов, производных ксилоглюкана. Physiol. Растение. 89, 1–3.
Google Scholar
Гибо Д. М. и Карпита Н. С. 1994 Биосинтез полисахаридов клеточной стенки растений. FASEB J. 8, 904–915.
Google Scholar
Иияма К., Лам Т. Б. Т и Стоун Б. А 1994 Ковалентные поперечные связи в клеточной стенке.Plant Physiol. 104, 315–320.
Google Scholar
Исии Т., Мацунага Т. и Хаяши Н. 2001 Образование димера бората рамногалактуронана II в пектине определяет толщину клеточной стенки тыквенной ткани. Plant Physiol. 126, 1698–1705.
Google Scholar
Kemsley E K 1998 Дискриминантный анализ спектроскопических данных. Джон Вили и сыновья, Чичестер, Великобритания, 179 стр.
Google Scholar
Levy S, York W. S., Stuikeprill R, Meyer B и Staehelin L.A 1991 Моделирование статических и динамических молекулярных конформаций ксилоглюкана — роль фукозилированной боковой цепи в поверхностно-специфичном складывании боковой цепи. Плант Ж. 1, 195–215.
Google Scholar
Лолле С. Дж., Берлин Г. П., Энгстром Е. М., Кроликовски К. М., Рейтер В. Д. и Прюитт Р. Э. 1997 Регуляция развития клеточных взаимодействий у мутанта Arabidopsis fiddlehead 1: роль эпидермальной клеточной стенки и кутикулы.Devel. Биол. 189, 311–321.
Google Scholar
Lukowitz W, Mayer U and Jurgens G 1996 Цитокинез в эмбрионе Arabidopsis включает связанный с синтаксином продукт гена KNOLLE . Cell 84, 61–71.
Google Scholar
Макканн М. С. и Робертс К. 1991 Архитектура первичной клеточной стенки. В Цитоскелетная основа роста и формы растений.Эд. К. В. Ллойд. С. 109–129. Academic Press, Лондон.
Google Scholar
Макканн М.К., Хаммури М., Уилсон Р., Белтон П. и Робертс К. 1992. Инфракрасная микроскопия с преобразованием Фурье — это новый способ изучения клеточных стенок растений. Plant Physiol. 100, 1940–1947.
Google Scholar
Макканн М.К., Чен Л., Робертс К., Кемсли Е.К., Сене С., Карпита Н.С., Стейси Н.Дж. и Уилсон Р.Х. 1997 Инфракрасная микроспектроскопия: неоднородность образцов в составе и архитектуре клеточных стенок растений.Physiol. Растение. 100, 729–738.
Google Scholar
Nicol F, His I, Jauneau A, Vernhettes S, Canut H и Höfte H 1998 Связанный с плазматической мембраной предполагаемый эндо-1,4- ? -D-глюканаза необходима для нормальной сборки стенок и удлинения клеток в Arabidopsis . EMBO J. 17, 5563–5576.
Google Scholar
Nishitani K 1995 Эндо-ксилоглюкантрансфераза, новый класс трансфераз, участвующих в построении клеточной стенки.J. Plant Res. 108, 137–148.
Google Scholar
Pear J R, Kawagoe Y, Schreckengost W. E, Delmer D. P, Stalker D. M 1996 Высшие растения содержат гомологи бактериальных генов cel A, кодирующих каталитическую субъединицу целлюлозосинтазы. Proc. Natl. Акад. Sci. USA 93, 12637–12642.
Google Scholar
Потиха Т. и Делмер Д. П. 1995 Мутант Arabidopsis thaliana , демонстрирующий измененные модели отложения целлюлозы.Плант Ж. 7, 453–460.
Google Scholar
Ральф Дж., Граббер Дж. Дж. И Хэтфилд Р. Д. 1995 Сшивки лигнин-ферулат при активном в травах включении сложных эфиров ферулированных полисахаридов в лигнины райграса. Carbohydr. Res. 275, 167–178.
Google Scholar
Reiter W-D 1998 Arabidopsis thaliana в качестве модельной системы для изучения синтеза, структуры и функции клеточной стенки растений.Plant Physiol. Биохим. 36, 167–176.
Google Scholar
Рейтер В.Д., Чаппл С.С. и Сомервилл С.Р. 1993 Изменение роста и клеточных стенок у мутанта Arabidopsis с дефицитом фукозы. Science 261, 1032–1035.
Google Scholar
Reiter W-D, Chapple C и Somerville C. R 1997 Мутанты Arabidopsis thaliana с измененным составом полисахаридов клеточной стенки.Плант Ж. 12, 335–345.
Google Scholar
Reiter W-D и Vanzin G F 2001 Молекулярная генетика путей взаимного превращения нуклеотидного сахара в растениях. Завод Мол. Биол. 47, 95–113.
Google Scholar
Рейнольдс Дж. О., Эйссес Дж. Ф. и Сильвестр А. В. 1998 Баланс деления и расширения во время морфогенеза листа кукурузы: анализ мутанта, warty 1.Разработка 125, 259–268.
Google Scholar
Ричмонд Т. А. и Сомервилл С. Р. 2001 Интегративные подходы к определению функции Csl. Завод Мол. Биол. 47, 131–143.
Google Scholar
Рудалл П. Дж. И Кэддик Л. Р. 1994 Исследование присутствия фенольных соединений в стенках однодольных клеток с использованием УФ-флуоресцентной микроскопии. Аня. Бот. 74, 483–491.
Google Scholar
Сене С. Ф. Б., Макканн М. С., Уилсон Р. Х. и Гринтер Р. 1994 Рамановская спектроскопия с преобразованием Фурье и инфракрасная спектроскопия с преобразованием Фурье — исследование пяти клеточных стенок высших растений и их компонентов. Plant Physiol. 106, 1623–163.
Google Scholar
Смит Б.Г. и Харрис П.Дж. 1999 Полисахаридный состав клеточных стенок Poales: Клеточные стенки Poaceae не уникальны.Биохим. System Ecol. 27, 33–53.
Google Scholar
Smith L G, Hake S and Sylvester AW 1996 Запутанная мутация 1 1 изменяет ориентацию деления клеток на протяжении всего развития листа кукурузы без изменения формы листа. Разработка 122, 481–489.
Google Scholar
Стэхелин Л. А. и Мур И. 1995 Аппарат-структура Гольджи растения, функциональная организация и механизмы торговли.Анну. Rev. Plant Physiol. Завод Мол. Биол. 46, 261–288.
Google Scholar
Тейлор Н.Г., Шейбл В., Катлер С., Сомервилль С.Р. и Тернер С.Р. 1999 Локус НЕРЕГУЛЯРНЫЙ XYLEM 3 Arabidopsis кодирует целлюлозосинтазу, необходимую для синтеза вторичной клеточной стенки. Растительная клетка 11, 769–779.
Google Scholar
Тейлор Н.Г., Лори С. и Тернер С. Р. 2000 Для синтеза целлюлозы у Arabidopsis требуются множественные каталитические субъединицы целлюлозосинтазы.Растительная клетка 12, 2529–2539.
Google Scholar
Томпсон Дж. Э., Смит Р. С. и Фрай С. С. 1997 Ксилоглюкан подвергается интерполимерному трансгликозилированию во время связывания со стенкой растительной клетки in vivo : Данные двойного мечения C-13 / H-3 и изопикнического центрифугирования в трифторацетате цезия. Biochem J. 327, 699–708.
Google Scholar
Тернер С. Р. и Сомервилль С. Р. 1997 Фенотип коллапсированной ксилемы Arabidopsis идентифицирует мутанты, дефицитные по отложению целлюлозы во вторичной клеточной стенке.Растительная клетка 9, 689–701.
Google Scholar
Ванзин Г. Ф., Мэдсон М., Карпита Н. С., Райхель Н. В., Кигстра К., Рейтер В. Д. 2002 г. У мутанта mur2 Arabidopsis thaliana отсутствует фукозилированный ксилоглюкан из-за повреждения фукозилтрансферазы AtFUT1. Proc. Natl. Акад. Sci. USA 99, 3340–3345.
Google Scholar
Verica J A и Medford J I 1997 Экспрессия модифицированного MERI5 изменяет рост клеток в трансгенных растениях Arabidopsis .Plant Sci. 125, 201–210.
Google Scholar
Уиллатс В. Г. Т., Маккартни Л., Маки В. и Нокс Дж. П. 2001 Пектин: клеточная биология и перспективы функционального анализа. Завод Мол. Биол. 47, 9–27.
Google Scholar
Xu W, Purugganan M M, Polisensky D. H, Antosiewicz D. M, Fry S. C. и Braam J. 1995 Arabidopsis TCH 4, регулируемый гормонами и окружающей средой, кодирует ксилоглюкановую эндотрансгликозилазу.Растительная клетка 7, 1555–1567.
Google Scholar
Zablackis E, Huang J, Muller B, Darvill A G и Albersheim P 1995 Структура стенок растительных клеток. 34. Характеристика полисахаридов клеточной стенки Arabidopsis thaliana листьев. Plant Physiol. 107, 1129–1138.
Google Scholar
Zablackis E, York W. S., Pauly M, Hantus S, Reiter W-D, Chapple C. C. S., Albersheim P и Darvill A, 1996 г. Замена L-фукозы на L-галактозу в клеточных стенках Arabidopsis mur 1.Science 272, 1808–1810.
Google Scholar
Исторические корни расизма | Объединенная методистская церковь
«Свобода не просто появляется, мы ее ищем».
Эрин Хокинс, Генеральная комиссия по вопросам религии и расы
Историческая ратуша
За несколько дней до 4 июля 2020 года, дня, когда многие в Соединенных Штатах задумываются об истории нашей страны, Объединенная методистская церковь провела беседу из серии Демонтаж расизма: стремление к свободе .Многие борются с тем, как нам наиболее достоверно понять и истолковать реалии настоящего момента. Мы молимся и надеемся на еще нереализованное будущее, в котором жизнь черных будет иметь такое же значение для лиц, принимающих решения в нашей стране, и для каждого верующего человека, как и для нашего всемогущего Бога.
В разгар этой жизненно важной работы по расчету с настоящим и переосмыслению будущего мы приглашаем вас быть подобными птице Санкофа: оглядываться назад, даже когда мы движемся вперед.
Модератор Эрин Хокинс поделилась своими знаниями и увлечением, ведя беседу.
Д-р Лиза Деллинджер, пастор Миссионерской конференции индейцев Объединенной методистской церкви в Оклахоме и эксперт в области расизма, Доктрины открытий и истории коренных народов Америки, открывает беседу, рассказывая о том, как европейские поселенцы в Северной Америке относятся к коренным народам. народов и его отношение к христианской церкви.
Генеральный секретарь Объединенной методистской комиссии по архивам и истории преподобный Фред Дэй III затем сталкивается с гордостью и стыдом причастности методистской церкви к расовой динамике Соединенного Королевства.С. и почему важно, чтобы Объединенные методисты знали эту историю.
Преподобный доктор Бобби Макклейн, пастор объединенной методистской церкви и автор книги « чернокожих в методистской церкви» , затем разделяет принципы расизма, которые необходимо разрушить в церкви.
Доктор Элисон Грин, специалист по истории религии США, затем преподает теологии превосходства белых и расового насилия.
Спикеры рассказали, где Бог призывает Объединенную методистскую церковь действовать как единое целое и как отдельные члены.
Впереди еще ратуши
Это первый из того, что будет серией разговоров. Второй будет сосредоточен на межсекторальном активизме между поколениями, объединяющем старейшин Объединенного методистского движения и новых лидеров, чтобы исследовать подходы «старой школы» и «новой школы» к организации расовой справедливости в церкви и сообществе. Ожидается, что следующая ратуша состоится в июле / августе, а дата будет объявлена дополнительно. Мы опубликуем даты на UMC.org/EndRacism.
Присоединяйтесь к нам, чтобы увидеть полную серию!
Посмотреть другие панельные дискуссии
Панель
Эрин Хокинс, модератор
г-жаЭрин М. Хокинс — генеральный секретарь Генеральной комиссии по вопросам религии и расы. Она является ведущим должностным лицом деноминационного агентства, которое культивирует расовую интеграцию и полное участие всех человек в работе, свидетельстве и жизни UMC. GCORR дает возможность церковному духовенству и мирскому руководству использовать ценности инклюзивности, расового равенства и справедливости в преобразующей работе жизненно важных общин, чтобы созидать Тело Христово. Г-ж Hawkins работает на уроки акции в создании святого отношения с Богом, «держа в напряжении нашей способности к величию, что призывает нас, как христиане, упорствовать в борьбе на пути становления нашего лучше самих, и для борьбы наших худших тенденций расизма, сексизм и классизм.”
Две степени магистра г-жи Хокинс в области организационного развития (Американского университета в Вашингтоне, округ Колумбия) и государственной политики (Университета Индианы) позволили ей понять, как системные процессы могут увековечить грех расизма и распространиться от местного к глобальному. арена.
Преподобный доктор Бобби Макклейн
Доктор Макклейн задумал и возглавил комитет, который выпустил сборник гимнов Песней Сиона , который изменил состав христианских гимнов всех деноминаций.Совсем недавно он был сопредседателем комитета по сиквелу « песен Сиона: Сион все еще поет»! Для каждого поколения написал Приходи воскресенье: литургический компаньон песням Сиона и завершил Афроамериканская проповедь и Библия: проповедь Сиона , третью книгу в этой трилогии
В 1978 году доктор Макклейн основал и работал исполнительным директором Многонационального центра служения в Университете Дрю, Мэдисон, Нью-Джерси. Там он написал Путешествующий свет: христианские взгляды на плюрализм и является автором книги Чернокожих в методистской церкви: куда ты идешь и вместе с покойным доктором Др.Грант Шокли и доктор Карен Коллиер, Наследие и надежда: афроамериканское присутствие в методизме .
- Магистр и докторская степень, Школа теологии Бостонского университета
- BA, с отличием , Clark College, Атланта, Джорджия
Загрузите более полную биографию.
Доктор Лиза Деллинджер
Д-р Лиза Деллинджер (Чикасо и американка мексиканского происхождения) получила степень магистра богословия с отличием в 2008 году в Богословской семинарии Филлипса в Талсе, штат Оклахома.
Она также служила пастором на Индийской миссионерской конференции Объединенной методистской церкви в Оклахоме с 2015 по 2017 год. Она получила докторскую степень по теологии, истории и этике в Гарретт-евангелической семинарии в 2020 году.
Она написала главы для книг Coming Full Circle: Constructing Native Christian Theology и Feminist Praxis Against U.S. Militarism. Она принимает участие в панельных дискуссиях по вопросам расизма, Доктрины открытий и коренных народов Америки, феминизма и христианства.Д-р Деллинджер также является стипендиатом Объединенной методистской ассоциации цветных женщин.
- Докторантура, Гарретт-Евангелическая семинария
- MDiv, Богословская семинария Филлипса, Талса, Оклахома
Ред. Дэй III
Альфред Т. Дэй, с 2014 г., является Генеральным секретарем Генеральной комиссии по архивам и истории. Он является членом духовенства Ежегодной конференции Восточной Пенсильвании. Он также является редактором журнала Methodist History.
До своего нынешнего назначения Дэй служил местным церквям на Ежегодной конференции Восточной Пенсильвании, в том числе был окружным суперинтендантом, а в последнее время — пастором исторической церкви Св.Георгия, памятник объединенного методистского наследия.
Будучи пастором Исторического Св. Георгия, он организовал и участвовал в исторических службах воссоединения и разделял служения между Историческим Св. Георгием UMC, Матерью Вефилем AME, Африканской епископальной церковью Св. Фомы и Матерью Африканской Цоар (2009-2014).
Он работал адъюнкт-профессором исследования объединенных методистов в Принстонской духовной семинарии (2011-14).
- Аспирантура, Университет Дрю и Университет Темпл
- M Div, Духовная семинария Гордона Конуэлла, 1975
- BS, Houghton College, Houghton NY, 1972
Др.Элисон Грин
Доктор Элисон Коллис Грин преподает религиозную историю Соединенных Штатов, с интересом к американским религиям, поскольку они связаны с политикой, богатством и бедностью, расой и этнической принадлежностью, окружающей средой и современным сельским югом. В дополнение к ее роли в Candler, Грин является аффилированным преподавателем факультета истории в Колледже искусств и наук Эмори. Она является автором книги No Depression in Heaven: The Great Depression, the New Deal and the Transformation of Religion in the Delta (Oxford, 2016), а также ряда эссе и статей по современной религиозной истории США в обеих странах. научные и популярные издания.Грин — выпускница YTI в Candler в 1996 году и член Консультативного совета YTI. Она также является членом редакционной коллегии журнала « Journal of Southern Religion».
- доктор философии, Йельский университет, 2010 г.
- MA и MPhil, Йельский университет, 2008 г.
- Бакалавр, Университет Северной Каролины в Чапел-Хилл, 2001 г.
Посмотреть резюме доктора Грина.
Регулирование состава и разборки фокальных комплексов кальций-зависимой протеазой кальпаином | Журнал клеточной науки
Чтобы изучить, как кальпаин модулирует организацию и разборку участков адгезивного контакта, мы исследовали эффекты ингибирования кальпаина на локализацию различных компонентов фокальной адгезии.В отличие от других изученных компонентов (зиксин, FAK, паксиллин, интегрин, талин и винкулин), актин-связывающий белок α-актинин показал сниженную локализацию в местах контакта с адгезивом после ингибирования кальпаином в клетках CHOK1 (рис. 5). α-Актинин представляет собой актин-связывающий белок, который перекрестно связывает актиновые филаменты и интегрин-содержащие адгезивные комплексы и локализуется как вдоль актиновых стрессовых волокон, так и в очаговых адгезиях (Otey et al., 1993). Используя как окрашивание антителом для α-актинина, так и экспрессию слитого белка EGFP-α-актинин, мы обнаружили, что α-актинин показал сниженную локализацию в сайтах адгезивных комплексов после ингибирования кальпаина с помощью ALLN или кальпастатина (рис.5). После ингибирования кальпаина мы обнаружили локализацию α-актинина по периферии клетки. В исследованиях живых изображений с использованием клеток CHOK1, котрансфицированных EGFP-α-актинином и контрольным вектором или кальпастатином, ингибирование кальпаина подавляло динамику и локализацию α-актинина в сайтах адгезивных комплексов (рис. 5). В контрольных клетках EGFP-α-актинин был очень динамичным и локализовался в сайтах адгезивных комплексов в центре и на периферии клетки. Напротив, экспрессия кальпастатина индуцировала периферическое распределение α-актинина и ингибировала динамику α-актинина.Интересно и то, что в отличие от α-актинина, зиксин, который обычно локализуется как в фокальных комплексах, так и в актиновом цитоскелете, присутствует преимущественно в фокальных комплексах после ингибирования кальпаина, а его совместная локализация с α-актинином снижается в клетках CHOK1 (рис. 6). ). В контрольных клетках альфа-актинин показал 80% совместную локализацию с зиксином, но после обработки клеточно-проницаемыми ингибиторами кальпаина наблюдалась совместная локализация только 31%. Иммунофлуоресценцию или котрансфекцию использовали для характеристики структур, содержащих циксин.Мы обнаружили, что как в контрольных, так и в обработанных ALLN клетках комплексы зиксина содержали α5-GFP-интегрин (Laukaitis et al., 2001) (рис. 7a), винкулин (рис. 7b), паксиллин, FAK и талин (данные не указаны). показано). Примечательно, что локализация α5-интегрина и винкулина в периферических фокальных комплексах была более заметной в клетках, обработанных ALLN. Чтобы определить, приводит ли неспецифическое нарушение актинового цитоскелета к аналогичным изменениям в локализации α-актинина и зиксина, клетки обрабатывали препаратом, разрушающим актин, цитохалазином D.Мы обнаружили, что, в отличие от ингибирования кальпаина, цитохалазин D нарушал образование очаговых комплексов с потерей очаговых адгезий как в центре клетки, так и на периферии, и не приводил к специфическому нарушению совместной локализации α-актинина и зиксина (данные не показано). В совокупности эти находки подтверждают, что кальпаин регулирует состав фокальных спаек и специфическую локализацию α-актинина в участках адгезивных комплексов.
Инструктивный внеклеточный матрикс легкого: основной состав и изменения при хроническом заболевании легких
Часть A: легочный ECM
Эволюция сложных тканей у высших организмов сопровождается увеличением разнообразия белков и структурной организации в ECM [ 1], а также расширение разнообразия рецепторов ЕСМ на клетках.Как и во всех других системах органов, легочный ECM состоит из двух основных структурных типов: 1) базальные мембраны, которые представляют собой тонкие слои гликопротеинов, которые покрывают базальную сторону эпителия и эндотелия и окружают мышечные, жировые и периферические нервные клетки; и 2) интерстициальные матрицы, которые образуют рыхлую и похожую на фибриллы сетку, которая связывает структурные типы клеток в тканях и тем самым поддерживает трехмерную (3D) когезию и биомеханические характеристики легких [2].Как базальные мембраны, так и интерстициальные матрицы образуют тканеспецифические «ниши», которые влияют на стволовость и дифференцировку популяций клеток-предшественников / стволовых клеток, а также на правильную функцию дифференцированных типов клеток, специфичных для ткани / компартмента.
Распутывание матрисомы
На основании биоинформатического анализа доступного генома млекопитающих, число основных структурных компонентов всего ЕСМ млекопитающего, как предполагается, включает ~ 300 белков (основная матрисома) [3, 4].Кроме того, ЕСМ связывается и служит резервуаром для большого количества секретируемых белков, таких как факторы роста, модифицирующие ЕСМ ферменты или другие связанные с ЕСМ белки, которые не вносят вклад в структуру ЕСМ, но влияют на его функцию как обучающая «ниша» (белки, ассоциированные с матрисомой). Из-за их сложных биохимических свойств (нерастворимость, высокая молекулярная масса и высокая степень сшивки белков) подробный анализ белков ЕСМ был очень сложным [5]; однако недавний прогресс в области оборудования для масс-спектрометрии (МС) и алгоритмов количественной оценки впервые позволил провести всесторонний анализ протеомов ЕСМ из различных тканей [6–13].Количественная оценка белков без метки с помощью МС не только позволяет относительное сравнение белков по образцам и условиям эксперимента, но также позволяет оценить абсолютные количества [14]. Используя этот подход, мы проанализировали наш недавно опубликованный набор протеомных данных протеома легких мышей [12] и оценили относительное содержание белков коровой матрицы, что позволило получить углубленный профиль экспрессии ECM в легких здоровых мышей (рис. 1).
РИСУНОК 1Протеом внеклеточного матрикса (ЕСМ) взрослого здорового легкого мыши.а) Мы использовали наш недавно опубликованный набор протеомных данных протеомов легких мышей для расчета относительной массовой доли белка указанных категорий генов. Интенсивности МС белков в пределах изображенных категорий генов выражали в миллионных долях от общей интенсивности МС всех белков. б – г) Для определения относительного числа копий белков ЕСМ в протеоме легких интенсивность МС нормализовали к теоретическому количеству триптических пептидов (нормализация размера белка; iBAQ).Мы использовали эту оценку числа копий белка для ранжирования коллагенов (b), протеогликанов (c) и гликопротеинов (d) по их количеству. МС: масс-спектрометрия; iBAQ: абсолютное количественное определение на основе интенсивности.
Состав ECM
Коллагены составляют основную часть протеина в легких [15], и они сильно изменяются при многих заболеваниях легких (рис. 2). Фибриллярные коллагены (типы I, II, III, V и XI), которые имеют большую прочность на разрыв, но низкую эластичность, вносят вклад в общую архитектуру легкого [16, 17], тогда как большие эластичные волокна, которые характеризуются низкой прочность на разрыв и высокая эластичность, обеспечивают легкому необходимую податливость и упругую отдачу.Эластичные волокна состоят из двух различных компонентов: эластин белка ЕСМ находится в его сшитой форме во внутренней сердцевине эластичных волокон, тогда как внешняя периферия эластичных волокон содержит микрофибриллы размером 10-15 нм [18].
РИСУНОК 2Патологические изменения внутри интерстициального внеклеточного матрикса (ЕСМ) в пораженном легком. а) Обзор легкого и участков образования болезни. б) Здоровый интерстициальный ECM поддерживается активностью резидентных фибробластов и представляет собой рыхлую сеть коллагенов, эластина и фибронектина, прикрепленную к базальной мембране слоя эпителиальных клеток.c) При идиопатическом фиброзе легких (IPF) фибробласты трансдифференцируются в миофибробласты с высокой сократимостью, которые откладывают высокие уровни молекул ECM в интерстиции и резко увеличивают жесткость ECM за счет ферментативного ковалентного сшивания коллагена и эластина. г) Признаками патологических изменений внутри ВКМ при хронической обструктивной болезни легких (ХОБЛ) являются обширное разрушение эластических волокон ферментами, разрушающими ВКМ, высвобождаемыми воспалительными клетками, наряду с повышенными уровнями гиалуронана и тенасцина С и сниженным отложением декорина. .Разрушение слоя эпителиальных клеток приводит к увеличению воздушного пространства (эмфиземе). д) При легочной артериальной гипертензии (ЛАГ) ремоделирование внеклеточного матрикса в стенке артерии характеризуется увеличением эластиновых и коллагеновых волокон, фибронектина и тенасцина С и гиперплазией гладкомышечных клеток. е) При астме характерные изменения ВКМ происходят под эпителием бронхов и утолщенной базальной мембраной. Кроме того, гладкомышечные клетки подвергаются гиперплазии, и увеличивается отложение коллагенов, фибронектина, гиалуронана и декорина.ж) При раке опухоли на первичной и метастатической сторонах окружены обширной жесткой стромой, которая содержит сильно сшитые коллагены и высокие уровни фибронектина, тенасцина С и гиалуронана. з) Легенда, изображающая молекулы и типы клеток.
Основными структурными компонентами микрофибрилл являются крупные гликопротеины фибриллин-1, -2 и -3 [19]. Кроме того, другие белки, такие как гликопротеины, ассоциированные с микрофибриллами, фибулины, белки, расположенные на границе раздела микрофибрилл эластина (EMILINs) и члены семейства эластинсшивающих лизилоксидаз (LOX), связаны с микрофибриллами или самим эластином [18].Интересно, что связанный с микрофибриллами протеин 4 (MFAP4) был самым распространенным гликопротеином, обнаруженным в нашем наборе данных по матрисомам легких здоровых взрослых мышей (рисунок 1) [12]. У людей высокая экспрессия MFAP4 была показана не только для легких, но и для других высокоэластичных тканей, таких как сердце и кишечник [20]. Биологическая функция MFAP4 в значительной степени неизвестна; Хотя он был идентифицирован как сывороточный биомаркер гаптического фиброза, он не может служить биомаркером фиброза легких [21].
Интерстициальный ECM альвеол состоит из расслабленной сети, в значительной степени основанной на коллагенах I и III типов и эластине как важных коровых белках [22, 23].Трехмерное расположение этой сети переплетенных волокон допускает нелинейное поведение напряжения-деформации (гистерезис и вязкоупругость), которое является характерным свойством мягких соединительных тканей [24]. Во время дыхания рассеяние энергии, вызывающее гистерезис и вязкоупругость в паренхиме легких, обусловлено контактами волокна с волокном в ECM коллаген-эластин, хотя типы сократительных клеток и сурфактант на границе раздела воздух-жидкость также могут способствовать этим эффектам [17]. . Помимо своих биомеханических функций, компоненты эластичных волокон, такие как фибулины и EMILINs, регулируют адгезию клетка-ECM, взаимодействуя с гетеромерными трансмембранными интегриновыми рецепторами [25, 26].В отличие от EMILINs, которые связывают интегрины через их домен gC1q-1, фибулин-5 связывает интегрины через эволюционно консервативную последовательность RGD (аргинин-глицин-аспарагиновая кислота). Этот трипептид-связывающий мотив RGD характерен для других гликопротеинов ВКМ, включая фибронектин, витронектин, остеопонтин, коллагены, тромбоспондины, фибриноген и фактор фон Виллебранда, все из которых опосредуют адгезию клетки-ЕСМ через рецепторы интегрина [27]. Помимо фиброзных коллагенов и гликопротеинов, протеогликаны (PGs), которые состоят из основного белкового компонента, ковалентно связанного с сульфатированными полисахаридами или гликозаминогликанами (GAG), являются основными составляющими ECM [28].Благодаря высокому содержанию полисахаридов PG являются гидрофильными, что способствует образованию гидрогелей и способствует вязкоупругости легких. PG содержат составляющие базальной мембраны перлекан и агрин, а также hyalectans versican, aggrecan, нейрокан, бревикан и подкласс протеогликанов с низким содержанием лейцина, включая его наиболее выдающиеся представители, декорин, бигликан и люмикан [29]. Декорин, бигликан и люмикан были одними из самых распространенных протеогликанов в нашем наборе данных по матрисомам здоровых легких взрослых мышей (рис. 1) [12]. In vitro , как декорин человека, так и бигликан способны связываться с профибротическим цитокином, трансформирующим фактор роста-β1 (TGF-β1), но in vivo , только декорин, но не бигликан, ингибирует фиброгенный эффект TGF- β1 [30]. Кроме того, сообщается, что декорин взаимодействует с различными металлопротеазами и может действовать как опухолевый супрессор, ослабляя рост, миграцию и ангиогенез опухоли [31].
Посттрансляционные модификации компонентов ECM
Посттрансляционные модификации (PTM), e.грамм. Ферментативное и химическое сшивание , трансглютаминирование, гликозилирование и гликозилирование, окисление и цитруллинирование, как известно, влияют на структурное и / или функциональное разнообразие белков ЕСМ [32]. Например, ферментативное поперечное сшивание коллагена и эластина, существенный шаг во время их биосинтеза, который в основном осуществляется ферментами семейства лизилоксидаз (LOX, LOXL1-4) и членами семейства трансглутаминаз (TG1-7, FXIII- A), обеспечивает компоненты ECM их характерной прочностью на разрыв [33–35].Кроме того, ферментативное (гликозилирование) или неферментативное (гликозилирование) добавление сахаров к белкам также влияет на биомеханику и функцию ЕСМ. В частности, в настоящее время гликирование считается фактором фиброза, связанного со старением. Этот процесс вызывает образование и накопление конечных продуктов гликирования (AGE). Аберрантные процессы гликирования были связаны с повышенной жесткостью тканей, например. в экспериментальном образовании AGE в сухожилиях, которые, как было показано, уменьшают скольжение волокна при увеличении растяжения волокна [36].Карбамилирование, представляющее собой неферментативное присоединение мочевины к остаткам изоциановой кислоты в белках, отрицательно влияет на стабильность и конформацию тройных спиралей коллагена I, а также на их расщепление под действием ММП [37]. Окисление белков ЕСМ реактивными формами кислорода (ROS) дополнительно модулирует качества ЕСМ, изменяя его производство, оборот и модификации, а также оказывая влияние на взаимодействия клетка-ЕСМ [38]. Кроме того, было обнаружено, что сывороточные уровни маркеров окислительного стресса повышены у пациентов с ИЛФ [39] и в конденсатах дыхания пациентов с астмой [40].ECM также может модулироваться цитруллинированием, ферментативной реакцией дезаминирования аргинина, которая влияет на адгезию клеток [41]. Для более полного обзора PTMs в ECM и их роли в заболевании мы отсылаем читателя к этим дополнительным обзорам [32, 42–45]. Взятые вместе, структурные и функциональные свойства легочного ECM могут быть изменены с помощью PTM, которые считаются важным потенциальным источником заболевания легких, как подробно описано в следующих разделах.
Биомеханика и перекрестные помехи между клетками и ЕСМ
Все клетки временно или постоянно прикрепляются к той или иной форме ЕСМ через рецепторы клеточной адгезии , наиболее заметно семейство интегринов трансмембранных гетеродимерных рецепторов [46, 47].Клетки могут использовать интегрины для исследования биохимических и топографических характеристик своего окружения [47, 48]. Сигнальная трансдукция, исходящая от адгезии клетка-ECM, контролирует клеточное поведение, такое как подвижность и распространение, морфология, выживаемость, пролиферация и дифференцировка [1, 22]. Эти сигнальные события зависят от множества белков, которые рекрутируются в кластеры интегринов плазматической мембраны, которые вместе называются «адгезомами» [49]. Недавние протеомные исследования очертили общий белковый состав адгезомы и его динамику под влиянием механических сил [50–55].Когда клетки сталкиваются со своим внеклеточным субстратом, они реагируют на жесткость этого субстрата множеством способов, включая немедленные изменения формы и активности клеток, а также долговременную экспрессию генов и изменения идентичности клеток. Эта механо-реципрокность генерируется посредством обратных связей между адгезиями клетка-ECM и цитоскелетом, которые настраивают силу сократительных сил на равновесие между приложенной силой и прочностью на разрыв субстрата ECM [47]. В этом процессе структура и организация цитоскелета изменяются, и было показано, что это вызывает долгосрочные изменения экспрессии генов [56].Точная молекулярная природа многих элементов в этих обратных связях в настоящее время неизвестна, и также неясно, в какой степени равновесие механо-чувствительности и механо-реакции различается между типами клеток и как такие различия могут быть запрограммированы. Важно отметить, что хорошо известно, что механо-чувствительные сигнальные пути важны в патофизиологии фиброзных заболеваний и рака, оба из которых обнаруживают динамические изменения в составе внеклеточного матрикса и механических свойствах во время прогрессирования заболевания [57–61].В частности, патологически повышенное отложение ВКМ, сопровождающееся ковалентным перекрестным связыванием и ремоделированием ВКМ, вызывает резкое увеличение жесткости ВКМ, вызывая, таким образом, механический градиент между патологической и нормальной тканью. Градиенты жесткости внутри ЕСМ диктуют миграционное поведение клеток в процессе, называемом дуротаксисом. Дуротаксис, по-видимому, является довольно общим явлением, поскольку было показано, что многие типы клеток, включая фибробласты, мезенхимальные стволовые клетки (МСК), миобласты и раковые клетки, перемещаются по градиентам жесткости [62–65].Кроме того, жесткость ВКМ влияет на распространение, сократимость и дифференцировку фибробластов [66, 67]. При выращивании на мягких матрицах фибробласты и многие другие клетки обычно демонстрируют низкие сократительные силы вместе с уменьшенным распространением и пролиферацией [68, 69]. Жесткость ВКМ также влияет на дифференцировку МСК, поскольку остеогенные и адипогенные клоны появляются на жестких и мягких поверхностях, соответственно [66]. Первичные культуры клеток альвеолярного эпителия типа II (ATII), которые, как считается, обладают потенциалом стволовых клеток и способны к долгосрочному самообновлению во взрослом легком, изменяют свою морфологию и функциональные характеристики (синтез и секреция сурфактанта) в зависимости от субстрат ЕСМ, используемый в культуре [28].В исследовании in vitro первичные альвеолярные макрофаги человека, которые опосредуют фиброз легких, показали увеличение фагоцитоза и изменения транскрипции и фенотипа при культивировании на жестком ECM [70]. Более того, для монослоя эндотелиальных клеток было продемонстрировано, что их культивирование на более жестком ECM оказывает негативное влияние на целостность монослоя, таким образом подтверждая важность проницаемости барьера в отношении трансмиграции лейкоцитов [71].
Резервная функция ЕСМ
ЕСМ служит резервуаром для ряда факторов роста и цитокинов, которые имеют решающее значение для дифференцировки и пролиферации клеток [4, 22, 72].Связь таких факторов, как фактор роста фибробластов (FGF), фактор роста гепатоцитов, а также латентный и активный TGF-β1 (полный список мы отсылаем читателя к ранее опубликованной таблице [72]) с ECM имеет некоторые последствия: ECM -связанные факторы 1) могут быть латентными, замаскированными или иметь иную активность по сравнению с их растворимой формой, 2) могут образовывать иммобилизованные градиенты, имеющие решающее значение для миграции клеток, 3) могут, как отпечатки прежней клеточной активности, иметь функцию памяти для инструктирования клеток поведение, 4) облегчают клеточную адгезию и рост клеток, и 5) как запасенные факторы могут быстро активироваться протеолитическим высвобождением и генерировать локальные сигналы, которые не зависят от медленных процессов, таких как экспрессия генов [72, 73].Деградация ECM является ключевым событием в ремоделировании ткани, и в этом процессе цинк-зависимые протеиназы (метцинцины) играют решающую роль.
ECM-разрушающие ферменты
Это суперсемейство металлопротеиназ включает матриксные металлопротеиназы (MMP) и адамализины. ММП представляют собой семейство из 25 цинк-зависимых эндопептидаз. Эти ферменты способны расщеплять все компоненты ВКМ [74] и базальные мембраны [75]. ММП секретируются в высоко согласованном процессе вместе с их ингибиторами, тканевыми ингибиторами металлопротеиназ (ТИМП) [76].Адамализины включают дезинтегрин и металлопротеиназы (ADAM) и ADAM с мотивом тромбоспондина (ADAMTS). Благодаря способности ADAM разрушать цитокины, факторы роста и лиганд FAS, им приписывают потенциальное клиническое применение при фиброзе, раке, воспалении и нейродегенерации [77]. При фиброзных заболеваниях легких производство и секреция ММП и ТИМП связывается с макрофагами [78], фиброцитами, эндотелиальными клетками и фибробластами [79], а также сильная экспрессия ММП, особенно ММП-1 и ММР-7. наблюдается в бронхиолярных и альвеолярных эпителиальных клетках [80].Для более подробного обсуждения молекулярных игроков в деградации и ремоделировании ECM, читатель может обратиться к превосходному обзору Lu et al. [81].
Часть B: процессы ремоделирования ЭКМ при хронических заболеваниях легких
IPF
Патогенез и ремоделирование тканей
Хотя этиология IPF остается в значительной степени неизвестной, IPF затрагивает примерно пять миллионов человек во всем мире. До недавнего времени единственным эффективным методом лечения была трансплантация легких. Недавно было показано, что два препарата, одобренные Управлением по санитарному надзору за качеством пищевых продуктов и медикаментов (FDA), пирфенидон и нинтеданиб, которые оказывают плейотропное действие на клетки, включая ингибирование продукции ECM, замедляют скорость снижения функции легких [82, 83].При IPF повторяющееся повреждение легких и последующие механизмы восстановления приводят к продолжающемуся разрушению эластичной паренхиматозной ткани легкого, которая постоянно замещается жесткой рубцовой тканью, состоящей из крупных и конденсированных агрегатов клетки-внеклеточного матрикса, известных как фибробластные очаги в легочном интерстиции (рис. 2c). . Прогрессивное ремоделирование ткани приводит к тракционным бронхоэктазам, утолщению межлобулярных перегородок и субплевральным сотам, что можно визуализировать с помощью компьютерной аксиальной томографии (CAT) [84]. Все эти микроскопические и макроскопические изменения способствуют прогрессирующей потере газообмена с последующим ухудшением функции легких, одышкой и, наконец, дыхательной недостаточностью, ведущей к смерти [85].
Миофибробласты
Фибробластные очаги содержат активированные фибробласты и / или высококонтрактильные миофибробласты, которые в основном характеризуются неоэкспрессией α-гладкомышечного актина (α-SMA) и его включением в волокна сократительного стресса. Помимо сократительных свойств, миофибробласты продуцируют большое количество молекул ВКМ в ответ на профибротические стимулы, которые, как полагают, являются результатом несбалансированных механизмов восстановления альвеолярного и бронхиолярного эпителия после повторяющихся микротравм и воспалительных процессов [86] (рисунок 2c).Происхождение миофибробластов в фиброзных очагах все еще остается предметом серьезных дискуссий. В различных исследованиях было показано, что миофибробласты в фиброзном легком происходят из клеток-предшественников костного мозга [87], циркулирующих фиброцитов периферической крови [88, 89], альвеолярных эпителиальных клеток, претерпевающих эпителиально-мезенхимальный переход [90], резидентных фибробластов [ 91] и перициты [92]. Довольно спорно, применяя трассировку клонов, другое исследование, описало еще неустановленную популяцию резидентов стромальных клеток в фиброзных очагах в качестве источника миофибробластов, но исключить перициты и эпителиальные клетки в качестве источников [93].
Инструктивность
Поскольку миофибробласт считается отличительной чертой IPF, можно предположить, что ECM, продуцируемый миофибробластом, является поучительным компонентом заболевания, который способствует прогрессированию заболевания. Действительно, культивируя фибробласты, происходящие либо из IPF, либо из здоровых контрольных легких на фиброзной или здоровой децеллюляризованной легочной ткани, Parker et al. продемонстрировал, что аберрантное микроокружение ЕСМ в IPF преимущественно управляет изменениями экспрессии генов и белков, а не внутренними клеточными изменениями [94].Процесс децеллюляризации и успешной рецеллюляризации целых органов, в идеале с использованием аутологичных стволовых клеток или клеток-предшественников, был протестирован как потенциальный суррогат в медицине трансплантации легких [95–97]. Даже трупные легкие человека, взятые у людей с хроническими заболеваниями легких, были исследованы на предмет возможного использования в тканевой инженерии ex vivo [98]. Однако в свете текущих открытий, что децеллюляризованные каркасы ECM от пациентов с IPF изменяют транскриптом и трансатом пересеянных фибробластов, остается интригующим открытым вопрос, могут ли такие суррогаты, полученные из патологических тканей легких, вообще использоваться для трансплантации легких.
Механофизиология
Современные концепции в этой области предполагают важнейшие функции конкретных белков ЕСМ в тонкой настройке сигнальных путей за счет их способности модулировать передачу сигналов факторов роста [99–101] или важные эффекты жесткости тканей на прогрессирование заболевания. Жесткость ВКМ при фиброзных нарушениях увеличивается за счет ферментативного ковалентного сшивания коллагена и эластина членами семейства LOX и TG [34, 35]. Было показано, что при IPF экспрессия лизилоксидазоподобного 2 (LOXL2) и TG2 повышается в сыворотке и легких, соответственно, пациентов с IPF [102, 103].Жесткость децеллюляризованной ткани IPF действительно выше, чем обычно, как показывает атомно-силовая микроскопия [13], и, соответственно, активность немышечного миозина-II, который управляет сократимостью клеток, также выше в фиброзных областях пациенты с ИЛФ [57]. Таким образом, активность и выживание активированных фибробластов при фиброзных заболеваниях можно в значительной степени контролировать с помощью механических сигналов. Действительно, ингибирование опосредованной миозином-II сократимости клеток с использованием ингибитора Rho-киназы индуцировало апоптоз миофибробластов и улучшало экспериментальный фиброз легких [57].Чумперлин и его коллеги использовали измерения экспериментального фиброза в легких, поврежденных блеомицином, с помощью атомно-силовой микроскопии, чтобы выявить поразительное увеличение фиброзных зон с модулем упругости (который измеряет жесткость ткани в Паскалях), превышающим 2 кПа [68]. Интересно, что диапазон физиологической жесткости (0,2–2 кПа) удерживает фибробласты легких в состоянии покоя, тогда как субстраты с более высокой жесткостью (2–35 кПа), наблюдаемые в фиброзных легких, вызывают профиброгенный фенотип с высокой пролиферацией и синтезом ВКМ. ставки [68].Мы показали, что (мио) фибробластам требуется комбинация α5β1-интегрина и гетеродимеров семейства αV-интегрина для эффективного ответа на жесткость богатого фибронектином тканевого микроокружения, которое присутствует в тканевом фиброгенезе [50]. Интересно, что устранение интегринов αV-семейства снижает фиброз в нескольких органах, включая легкие, на экспериментальных моделях мышей с травмой [104], что может указывать на то, что эти интегрины участвуют в механо-чувствительной реакции [50] и / или активации TGF-β. [105].
Протеомные сигнатуры ECM
Будущие протеомные исследования острых по сравнению с хроническими моделями повреждения легких и последующего фиброза могут позволить идентифицировать компоненты внеклеточной ниши, которые выборочно приводят к патологии в постоянном и прогрессирующем фенотипе рубцевания, как видно на IPF. Мы провели протеомное исследование с временным разрешением на модели мышей с острым повреждением легких блеомицином и, основываясь на наблюдаемой временной кинетике изобилия белков, мы смогли предсказать функции отдельных белков на ранних стадиях восстановления ткани, таких как мобилизация стволовых клеток. или поздние стадии, такие как разрешение фиброза.Неожиданные белки ЕСМ, которые не были изучены в контексте повреждения по сравнению с восстановлением и фиброзом, такие как Эмилин-2 и коллаген-XXVIII, были обнаружены с высокой степенью активации через 2 недели после повреждения [12]. Используя сочетание метки стабильных изотопов, фракционирования растворимости децеллюляризованной легочной ткани и протеомного анализа, Декарис и его коллеги оценили кинетику синтеза и отложения ВКМ на модели блеомицина [106] и обнаружили, что скорость синтеза белка фибриллярных коллагенов (типы I, III и V) резко увеличивались как в растворимой, так и в нерастворимой фракциях во время позднего фиброзного ответа, тогда как небольшие богатые лейцином протеогликаны бигликан и декорин, а также фибронектин были повышены при раннем фиброзном ответе.Однако в другом анализе ECM на основе MS децеллюляризованных IPF легких авт. Идентифицировали обогащение гликозаминокликанами, матриксным Gla белком и ассоциированными с микрофибриллами белками в ECM легких IPF [13]. Безусловно, ECM пораженных легких человека может содержать специфическую для заболевания и прогрессирования комбинацию секретируемых белков и протеолитических фрагментов компонентов ECM («сигнатуры»), которая может иметь не только большую диагностическую и / или прогностическую ценность для точной медицины будущего. но также может дать новое понимание молекулярных механизмов патогенеза болезни.
ХОБЛ и эмфизема
Патофизиологические аспекты ХОБЛ, которая в основном вызвана воздействием дыма, включают обструкцию воздушного потока и гиперинфляцию. Ограничение воздушного потока при ХОБЛ вызвано тремя взаимосвязанными процессами: ремоделирование малых дыхательных путей, приводящее к утолщению стенок дыхательных путей, потеря мелких дыхательных путей и увеличение дыхательных пространств в альвеолах, состояние, называемое эмфиземой [17]. Следовательно, распространение транспульмонального давления нарушается, вызывая гиперинфляцию легких.Эти структурные изменения вызывают биомеханические изменения, которые проявляются в потере упругой отдачи легких. Поскольку ECM представляет собой основной компонент дыхательной системы, несущий нагрузку, потеря упругой отдачи при эмфиземе легких предполагает, что эластин, основной компонент эластичных волокон, является главной целью разложения (рис. 2d). На модели эмфиземы мышей как интратрахеальная инстилляция эластолитического фермента эластазы, так и дефицит α1-антитрипсина, основного ингибитора эластазы нейтрофилов, приводили к образованию эмфиземы [107].Дисбаланс протеазной и антипротеазной активности является широко принятой гипотезой о том, как разложение ткани может происходить при эмфиземе [108]. Высвобождение ферментов, разрушающих ECM, в значительной степени связано с воспалительными клетками, в основном макрофагами, нейтрофилами и Т-клетками [109]. Однако не только разрушение эластических волокон может быть ответственным за изменения биомеханики легких, поскольку есть также экспериментальные доказательства того, что ремоделирование коллагена в стенках альвеол вносит вклад в структурные и биомеханические изменения, обнаруживаемые при эмфиземе легких [110].Соответственно, утолщение коллагеновых фибрилл было продемонстрировано на ультраструктурном уровне у людей и на моделях мышей-грызунов [111]. Это довольно неожиданное открытие подтверждается данными in vivo и , свидетельствующих о том, что деградация эластазой компонентов ВКМ может запускать синтез коллагена [112], но описанный ответ также может быть результатом макрофагов, продуцирующих TGF-β1, которые стимулируют выработку коллагена эффекторными клетками. [111]. Однако аберрантное отложение коллагена при ХОБЛ по-разному влияет на разные отделы легких.В то время как электронные микрофотографии показали высокое содержание коллагена и эластина в стенках альвеолярной перегородки эмфизематозной ткани [113], более низкая экспрессия коллагена I была обнаружена в дыхательных путях пациентов с ХОБЛ [114]. Annoni et al. проанализировал подробный состав внеклеточного матрикса пациентов с ХОБЛ по сравнению со здоровым контролем в различных отделах легких и показал, что в образцах умеренной ХОБЛ наблюдались изменения в эластических волокнах, фибронектине, коллагенах, тенасцине-С и версикане во всех отделах легких. [114].В соответствии с этими выводами Roman et al. продемонстрировал индуцированное никотином усиление экспрессии фибронектина как in vitro, , так и in vivo [115]. Более того, in vitro , гладкомышечные клетки дыхательных путей (ASM), выделенные от пациентов с ХОБЛ, продуцировали более высокие уровни Col8a1, MMP1, MMP3 и MMP10 в ответ на экстракт сигаретного дыма [116]. Недавно было обнаружено, что мыши с дефицитом гликопротеина ЕСМ фибулина-4 имеют повышенную активность ММП и эластазы нейтрофилов в легких, что приводит к разрушению альвеол у новорожденных мышей и впоследствии к образованию эмфиземы [117].Было обнаружено, что при ХОБЛ уровни экспрессии остеопонтина, который представляет собой фосфорилированный кислый гликопротеин ВКМ с функциями клеточной адгезии и миграции, повышены в мокроте пациентов с ХОБЛ [118]. Повышенные уровни остеопонтина неспецифичны для ХОБЛ, поскольку аналогичная повышающая регуляция остеопонтина была описана в легких и жидкости бронхоальвеолярного лаважа пациентов с ИЛФ [119]. Было обнаружено, что AGE, которые в основном вызваны воспалительными процессами, накапливаются в коже пациентов с ХОБЛ.Это потенциально подразумевает роль AGEs и событий гликирования в ранних патологических фазах COPD, хотя прямое участие AGEs в ECM in vivo и в легких человека все еще остается неясным [120]. Кроме того, во время острых обострений ХОБЛ происходит ускоренный оборот белка ЕСМ, что приводит к продуцируемой протеазой фрагментации белков ЕСМ, таких как коллагены, версикан и эластины. Эти отпечатки пальцев белка ЕСМ, циркулирующего в крови, могут быть использованы в качестве мощных биомаркеров при прогнозировании прогрессирования ХОБЛ [121].
ЛАГ
ЛАГ — прогрессирующее заболевание с неблагоприятным прогнозом. Патофизиология ЛАГ характеризуется повышенной нагрузкой на правый желудочек сердца в результате нескольких процессов, таких как генетическая предрасположенность, воспаление, пролиферация клеток, сужение сосудов и ремоделирование сосудов. Легочные артерии подвержены стойкому сужению сосудов и ремоделированию сосудов, что приводит к снижению эластичности легочной артерии [122]. При ЛАГ вся трехслойная архитектура артериальной стенки претерпевает ремоделирование, что приводит к утолщению интимы, медиального и адвентициального каналов [123] (рис. 2а).Соответственно, исследования динамического включения с использованием радиоактивного индикатора показали быстрый синтез эластина и коллагена в легочных артериях во время гипоксической легочной гипертензии [124]. Кроме того, компонент ECM, тенасцин-C, был связан с прогрессированием заболевания. У пациентов с ЛАГ и на экспериментальных моделях заболевания тенасцин-C коррелировал с пролиферацией гладкомышечных клеток (ГМК) (рисунок 2e) и митогенным ответом на FGF2. Аналогичным образом, в легких мышей с фиброзом тенасцин-C отрицательно коррелировал с эластичностью легких, то есть большие количества этого белка были связаны с более жесткой легочной тканью [12].Между тем, положительная корреляция между экспрессией тенасцина-C и пролиферацией SMC была обнаружена у пациентов с врожденным пороком сердца и на модели грызунов с монокроталином-индуцированной ЛАГ [125]. Однако экспрессия эластина in vitro и обратно коррелировала с пролиферацией SMC [126]. Кроме того, в утолщенной интиме пораженных сосудов из тканей биопсии легких пациентов с ЛАГ были выявлены повышенные уровни фибронектина и тенасцина-C с помощью иммуногистохимии и гибридизации in situ и [125].Повышенное отложение ГАГ, таких как гиалуроновая кислота (ГК), также наблюдалось в реконструированных легочных артериях [127, 128]. Более того, патологическая модификация тяжелой цепи НА, которая, как предполагается, также способствует адгезии и активации лейкоцитов, была идентифицирована в ткани легких пациентов с ЛАГ [129].
Известно, что ММП играют решающую роль в ремоделировании легочных сосудов. In vitro и in situ исследования SMC легочной артерии выявили дисбаланс MMP и TIMP при идиопатической ЛАГ [130]; Было обнаружено, что MMP3 подвергается понижающей регуляции, тогда как TIMP1 активируется в пораженных SMCs [131].Nave et al. сообщил, что лизилоксидазы также участвуют в ремоделировании легочных сосудов при ЛГ, вызывая массивное перекрестное сшивание ECM [132]. Несмотря на гиперплазию и гипертрофию сосудистой стенки, обнаруженную в легких с легочной гипертензией, они были успешно децеллюляризованы и рецеллюляризованы мезенхимальными стволовыми клетками [133]. При успешной рецеллюляризации стволовыми клетками децеллюляризованные каркасы 3D ECM из больных легких могут представлять собой действительный источник в клиническом использовании для создания здоровых функциональных тканей.Однако предварительным условием является то, что процесс рецеллюляризации приводит к успешному ремоделированию больного гипертрофического ВКМ в децеллюляризованных каркасах, и, по крайней мере, при фиброзных заболеваниях наблюдается противоположный эффект [94].
Астма
Патология астмы характеризуется хроническим воспалением подслизистых областей малых дыхательных путей с ассоциированной гиперплазией бокаловидных клеток, повышенной секрецией слизи, гипертрофией гладких мышц и бронхоспазмом (рисунок 2f).На структуру легких влияет ремоделирование дыхательных путей, вызванное гиперплазией ASM, с сопутствующим измененным профилем ECM вокруг структур дыхательных путей. Эти структурные изменения проявляются в биомеханических дисфункциях. Соответственно, уменьшается упругая отдача легких, что может быть вызвано нарушением передачи упругой нагрузки между паренхимой легкого и дыхательными путями. Аберрантное накопление ВКМ ранее было обнаружено в легких, страдающих астмой, с его основной локализацией в подслизистых и адвентициальных областях как больших, так и малых дыхательных путей [134].Отложения ВКМ, которые возникают даже при легкой форме астмы, включают коллагены I, III и V в субэпителиальной области, а также фибронектин в ретикулярной пластинке бронхиального эпителия [135] (рисунок 2f). Кроме того, при астме со смертельным исходом сообщалось об увеличении эластических волокон, MMP-9 и MMP-12 в крупных дыхательных путях [136]. Интересно, что состав ВКМ центральных и дистальных дыхательных путей у пациентов со стойкими симптомами астмы, несмотря на терапию кортикостероидами (неконтролируемая астма), отличается от такового у пациентов с чувствительной к кортикостероидам (контролируемой) астмой.По сути, при неконтролируемой по сравнению с контролируемой астмой было обнаружено, что отложение коллагена увеличивается в альвеолярной паренхиме, тогда как версикан увеличивается в центральных дыхательных путях, а декорин и бигликан увеличиваются в обоих отделах [137]. Различия между дистальными отделами дыхательных путей и бронхиальным ремоделированием могут быть основаны на изменении статуса секреции ВКМ фибробластов, происходящих из этих разных областей [138]. Эти данные указывают на высокую клиническую значимость ремоделирования ВКМ при астме.Было показано, что в ASM присутствие растворимых форм фибронектина и коллагена I может уменьшать сократительную способность, но увеличивать пролиферативную способность ASM, тогда как культивирование клеток в присутствии только экзогенного растворимого ламинина не влияло ни на пролиферацию, ни на сократимость [ 139]. Однако механизм, объясняющий, как белки ЕСМ в их растворимых, не депонированных формах по-разному влияют на фенотип и сократительные функции в ASMs, остается неуловимым. Таким образом, изменения в ECM могут управлять механизмами, ведущими к усиленному росту ASM (рисунок 2f), что является ключевым событием в молекулярной патологии астмы [140].Более того, in vitro и исследований ASMs предполагают, что изменения в ECM также происходят на посттрансляционных уровнях при астме [140]. Аберрантное отложение компонентов внеклеточного матрикса по-разному влияет на биомеханические свойства легких. Отложение ВКМ вокруг дыхательных путей усиливает сужение дыхательных путей и вызывает изменения в жесткости тканей.
Рак легкого
Отличительными признаками рака у человека, которые были всесторонне описаны Ханаханом и Вайнбергом [141], являются многоступенчатые биологические возможности, которые включают устойчивую пролиферацию, уклонение от подавления роста, устойчивость к смерти, репликативное бессмертие, индуцированный ангиогенез, инициирование инвазия и метастазирование, нарушение регуляции клеточной энергетики и воспалительные процессы (рис. 2g).Поскольку ЕСМ по своим биохимическим и биофизическим свойствам является регулятором клеточных ответов, лежащих в основе признаков рака, изучение влияния ЕСМ на неопластическое прогрессирование становится важным [142]. Например, мелкоклеточный рак легкого (SCLC) окружен обширной стромой ECM, которая содержит высокие уровни фибронектина, ламинина, коллагена IV и тенасцина-C [143]. Более того, строма опухоли развитой склеротической опухолевой ткани богата коллагеном I, тогда как коллаген III очень распространен в менее зрелой строме [144].Трансляционные исследования показали, что у пациентов с опухолями SCLC, окруженными обширной стромой, время выживания было сокращено, поскольку связывание клеток SCLC с ECM защищает раковые клетки от апоптоза, вызванного химиотерапией [143]. Ремоделирование стромы опухоли сильно влияет на жесткость ткани, что, в свою очередь, влияет на механобиологию клеток и запускает клеточные реакции (рисунок 2g). Поскольку коллаген, как сообщается, вносит основной вклад в прочность на разрыв в архитектуре легких, метаболизм коллагена играет решающую роль в укреплении стромы опухоли.Помимо аберрантного отложения и обмена коллагена, при раке легких наблюдались изменения в перекрестном связывании коллагена. Совсем недавно Chen et al. сообщил, что строма опухоли демонстрирует высокие уровни поперечных связей коллагена, производных гидроксилизинового альдегида (HLCC), и более низкие уровни поперечных связей, производных лизинового альдегида (LCC) [145] (рисунок 2g). Механически этот переход от преимущественно LCC в здоровых к HLCC в опухолевой ткани легкого был связан с ферментом лизилгидроксилазой 2 (Lh3).Сопутствующее увеличение жесткости стромы, в свою очередь, способствовало инвазии опухолевых клеток и метастазированию [145]. В соответствии с этими открытиями известно, что опухолевые клетки экспрессируют высокие уровни LOX, и фармакологические вмешательства, направленные на это семейство ферментов, подавляли метастазирование у животных с опухолями [146]. Mouw et al. показали, что при раке молочной железы жесткость ECM индуцирует miR-18a для снижения экспрессии фосфатазы и гомолога тензина (PTEN), что, в свою очередь, приводит к прогрессированию опухоли [147]. Кроме того, повышенная жесткость ECM в опухолях возвращается к клеткам Rho-зависимым образом, увеличивая напряжение цитоскелета и стабилизируя фокальные спайки [148].Поскольку микросреда опухоли создается раком и стромальными клетками, активация стромальных клеток происходит аутокринным и паракринным образом. В частности, активированные фибробласты, так называемые фибробласты, ассоциированные с раком, играют важную роль в прогрессировании опухоли, существенно ремоделируя ECM опухоли, подавляя иммунный ответ и высвобождая факторы, способствующие росту опухоли [149]. Таким образом, опухолевый ЕСМ обеспечивает аберрантные сигналы микросреды, благоприятствующие пролиферации и метастазированию, а также ингибируя апоптоз опухолевых клеток.Соответственно, на первичных участках и участках метастазов инкапсулирующая строма опухоли может придавать устойчивость к химиотерапии. При SCLC было продемонстрировано, что адгезия опухолевых клеток через интегрин-β1 к белкам ECM является решающим событием в устойчивости клеток к химиотерапии [143]. Безусловно, патологические изменения в ECM при раке легких, такие как повышенная экспрессия коллагена, изменение перекрестного связывания коллагена и последующее увеличение жесткости тканей, напоминают изменения, также обнаруженные в IPF, который выступает за сходство между этими двумя заболеваниями.Интересно, что сообщается о многих патогенных аналогиях между раком и IPF: общие факторы риска ( например, курение, воздействие окружающей среды, вирусные инфекции и хроническое повреждение тканей), отсроченный апоптоз, активация специфических сигнальных путей, как эпигенетические, так и генетические изменения, измененная экспрессия микроРНК и аберрантная пролиферация и инвазия клеток [150–154]. Есть практические основания рассматривать IPF как рак, подобное заболеванию, что в конечном итоге может привести к новым клиническим испытаниям противораковых препаратов для IPF [154].
Модельные системы ECM
Состав ECM и его биомеханические свойства влияют на морфологию клеток [155], подвижность, распространение, жизнеспособность клеток и апоптоз, а также на пролиферацию и дифференцировку [5]. Следовательно, поскольку клеточное поведение и ECM сильно взаимосвязаны, необходимы системы культивирования клеток in vitro, , которые точно имитируют физиологические и патологические условия, обнаруженные in vivo, . В прошлом было разработано множество моделей in vivo на животных, отражающих патогенез заболеваний легких человека.Поскольку подробное обсуждение животных моделей, представляющих заболевания легких, далеко выходит за рамки этого обзора, мы отсылаем читателя к ряду отличных обзоров для дальнейшего чтения [156–162]. В частности, исследования мезенхимальных клеток, которые физиологически встроены в ЕСМ, могут быть полезны с использованием моделей in vitro, , in vivo, или ex vivo, , которые очень похожи по структуре и биомеханике интерстициального легочного ВКМ [163–165] . В прошлом применялись различные системы трехмерных культур клеток [166].Самая упрощенная модель — это пластик для тканевой культуры, покрытый молекулами ЕСМ. Благодаря упрощенной конструкции его можно использовать для изучения взаимодействия определенных молекул ЕСМ с клетками, хотя здесь клетки прикреплены к двумерной (2D) поверхности, а не встроены в ЕСМ. Более того, жесткость пластика клеточной культуры намного превосходит физиологически обнаруженный in vivo ; Согласно измерениям с помощью атомно-силовой микроскопии, модуль Юнга (измеренный в Паскалях) пластика клеточной культуры колеблется от 2 до 4 ГПа, тогда как нормальная ткань легких колеблется от 0.44 и 7,5 кПа, в зависимости от измеряемой области, а фиброзная ткань легкого человека у пациентов с ИЛФ в среднем составляет 16,52 кПа [13, 167, 168]. Однако механически настраиваемые 2D-модели с использованием синтетических матриц на основе акриламида или полиэтиленгликоля дают возможность индуцировать образование градиента и жесткий контроль жесткости ECM [166, 169]. Помимо этих искусственно синтезированных макромолекулярных структур, нативные и сконструированные биополимеры, состоящие из коллагенов [170], эластина [171], фибрина [172] или ламинина [173], широко используются в модельных системах in vitro и ECM [174].Гели экстракта базальной мембраны и децеллюляризованные матриксы на основе фибробластов также используются в настоящее время [175], и они более точно имитируют состав, трехмерную структуру и биомеханику физиологического внеклеточного матрикса. Однако ни одна из этих моделей не может имитировать уникальную пространственную геометрию легочной ткани. Чтобы обойти эти биомиметические ограничения, можно использовать нарезанные вибратомом матрицы ex vivo , взятые из различных моделей болезней животных, а также из пораженных тканей человека в их нативных или децеллюляризованных формах [13, 176–178]. Ex vivo модели , использующие децеллюляризованную легочную ткань, помогут дополнительно механистически определить влияние ECM на патологии легких. Наконец, в будущем технологии трехмерной биопечати или методы аддитивного производства могут открыть многообещающие и захватывающие новые способы создания биосовместимых каркасов ECM, которые биомеханически напоминают настоящую ткань. Такие изготовленные каркасы могут быть использованы либо в качестве настраиваемых моделей in vitro для изучения инструктивной природы ECM, либо, в конечном счете, при успешном пересеве с клетками, в качестве биоинженерной ткани, готовой заменить больные ткани или целые органы [179].
Заключение и перспективы на будущее
Хронические респираторные заболевания возникают в результате сложного взаимодействия факторов риска окружающей среды, эпигенетических и генетических влияний. Считается, что неблагоприятные воздействия окружающей среды во время жизни плода и младенца приводят к стойким изменениям в структуре и функции легких. Такое «программирование» на ранних этапах жизни могло бы ошибочно переделать ECM и, следовательно, проложить путь для развития хронических респираторных заболеваний во взрослом возрасте. Регулируемое ремоделирование внеклеточного матрикса, особенно в процессе заживления ран, его резидентными клетками является предпосылкой для поддержания гомеостаза тканей.В патологических событиях качества ECM, такие как его состав и топологические особенности, биомеханические свойства, PTM или роль резервуара для секретируемых медиаторов, действуют как поучительные сигналы, которые влияют на поведение фибробластов и других типов клеток. Как только гомеостаз ткани нарушается повторным повреждением и восстановлением, аберрантные механизмы обратной связи между клетками и их реконструированным ECM запускают порочный круг, который приводит к прогрессированию заболевания. Во время прогрессирования заболевания взаимодействие между клетками и ECM может быть очень сложным, может включать различные типы клеток и может происходить в пространственно различных компартментах.Например, успешное метастазирование требует раннего ремоделирования ВКМ в метастатической нише, которая пространственно удалена от локальной ниши первичной опухоли, чтобы помочь циркулирующим опухолевым клеткам прижиться в отдаленных органах [180]. Следовательно, определение молекулярного состава ЕСМ в пораженных легких с помощью современных технологий, таких как протеомика, управляемая МС или секвенирование следующего поколения, является важным шагом на пути к идентификации диагностических биомаркеров прогрессирования заболевания и одинаковая терапия.Будущие попытки разработать методы для понимания сложной функциональной и физической топологии ECM in situ на системном уровне будут ключевыми для понимания его роли в тканевой (пато) физиологии. Чтобы увидеть прорывы в разработке новых терапевтических вмешательств, важно понимать сложные взаимодействия и лежащие в основе механизмы обратной связи, которые происходят между основной матрисомой, факторами, связанными с матрисомой, и клетками [168, 181]. Потенциальные терапевтические средства для лечения фиброзных заболеваний включают ферменты, которые воздействуют на жесткость ECM путем разрушения аномального ECM, а также антитела, которые ингибируют активность ферментов сшивания ECM, таких как лизилоксидазы, а также активность MMP и TGF-β1 [73].Кроме того, ингибиторы клеточных рецепторов, таких как рецептор эпидермального фактора роста, или лекарства, нацеленные на механизм механотрансдукции через интегрины , в настоящее время тестируются в качестве терапевтических средств при таких заболеваниях, как рак легких и фиброзные расстройства [73]. Кроме того, молекулы и пути, участвующие в реакции легких на окислительный стресс, обсуждаются как потенциальные терапевтические мишени [38]. Также растет интерес к использованию клеток-предшественников, таких как мезенхимальные стволовые клетки, которые могут обладать регенеративным потенциалом, в качестве усилителей восстановления тканей [168].Однако на данный момент трансплантация легких остается единственным действенным методом лечения терминальной стадии заболеваний легких, но нехватка донорских легких и необходимость интенсивной иммуносупрессии для предотвращения отторжения аллотрансплантата ограничивают их клиническое применение.