Презентация «Состав чисел в пределах 20»
Просмотр содержимого документа
«Презентация «Состав чисел в пределах 20″»
МАТЕМАТИКА 1класс
Тема: «Решение задач и примеров в пределах 20
Решим цепочку примеров
День рождения кота Леопольда
Вставь пропущенное число
2 , 4 , 6 , 8 ,10
1 , 3 , 5 , 7 , 9
Увеличь каждое из чисел на 4
5
3
9
7
1
5
4
8
0
6
4
2
Уменьши каждое из чисел на 2
Уменьши каждое из чисел на 2
9
5
7
4
3
2
5
4
7
1
6
3
Телеграмма
0
6
7
1
2
3
8
4
9
5
10
о
з
д
р
а
в
л
я
е
м!
П
Реши задачи:
Пять ежей – с лукошками,
Три ежа – с гармошками.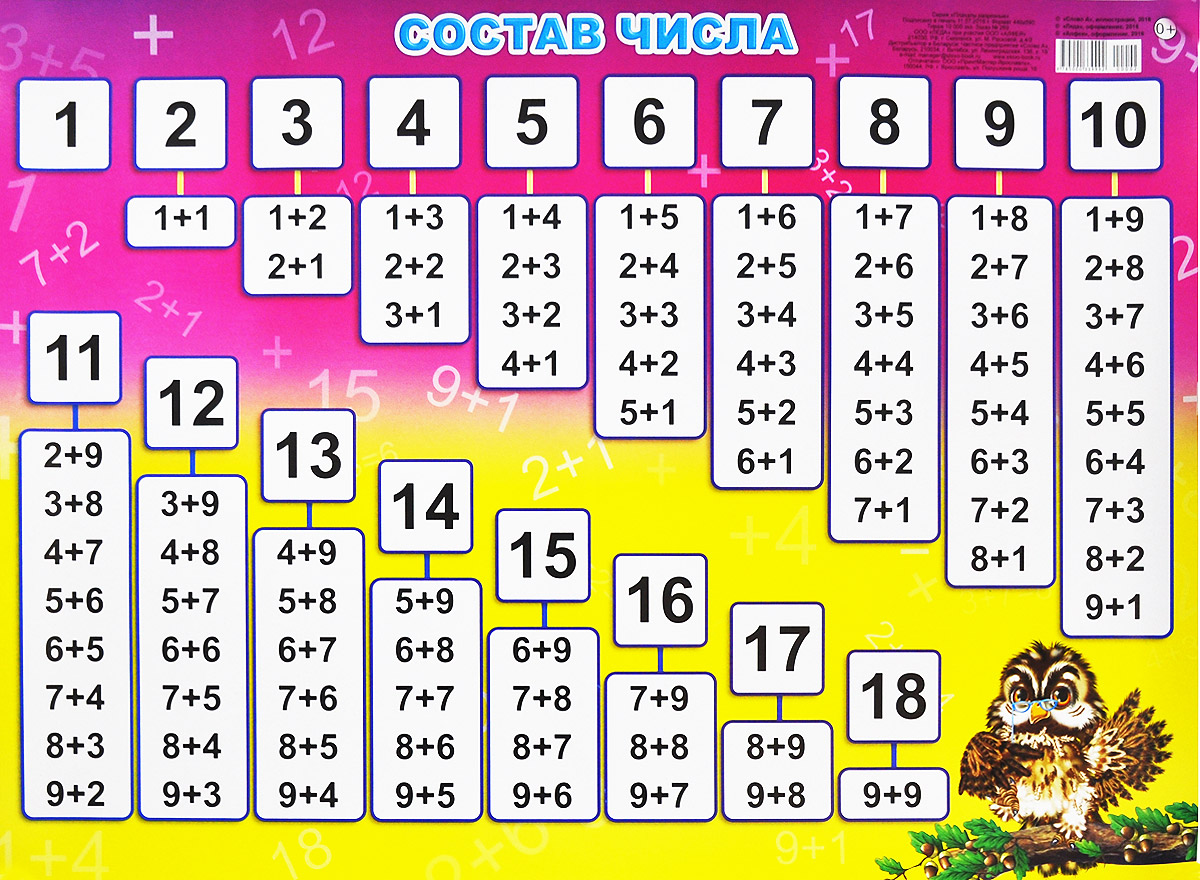
Сколько среди сосен
Ежей гуляет?
5+3=8
По дорожке мышка шла,
4-2=2
Четыре яблока несла,
Два – зайцу отдала,
Остальные в кладовку убрала.
Идут гулять козлята,
Четыре дружных брата.
А рядом три сестрицы –
Три козочки идут.
И братья, и сестрицы
Отправились гулять.
Попробуй-ка, сумей-ка
Их всех пересчитать.
4+3=7
Задача
- Курица весит 3 кг а козленок на 5кг больше . Сколько весит козленок .
- Напишите решение и ответ.
Решение:
3+5=8(кг)
Ответ : 8 кг
Исправь ошибки:
10 + 4 = 12
19 — 10 =8
16 – 6 =
7 + 3= 9
13 – 3 = 9
8 + 3 = 10
Поздравляем!
Самостоятельная работа
Проверь себя:
Математика :: Первоклассный урок
ОСНОВНЫЕ ТРЕБОВАНИЯ
к окончанию обучения в 1 классе
Учащиеся должны знать:
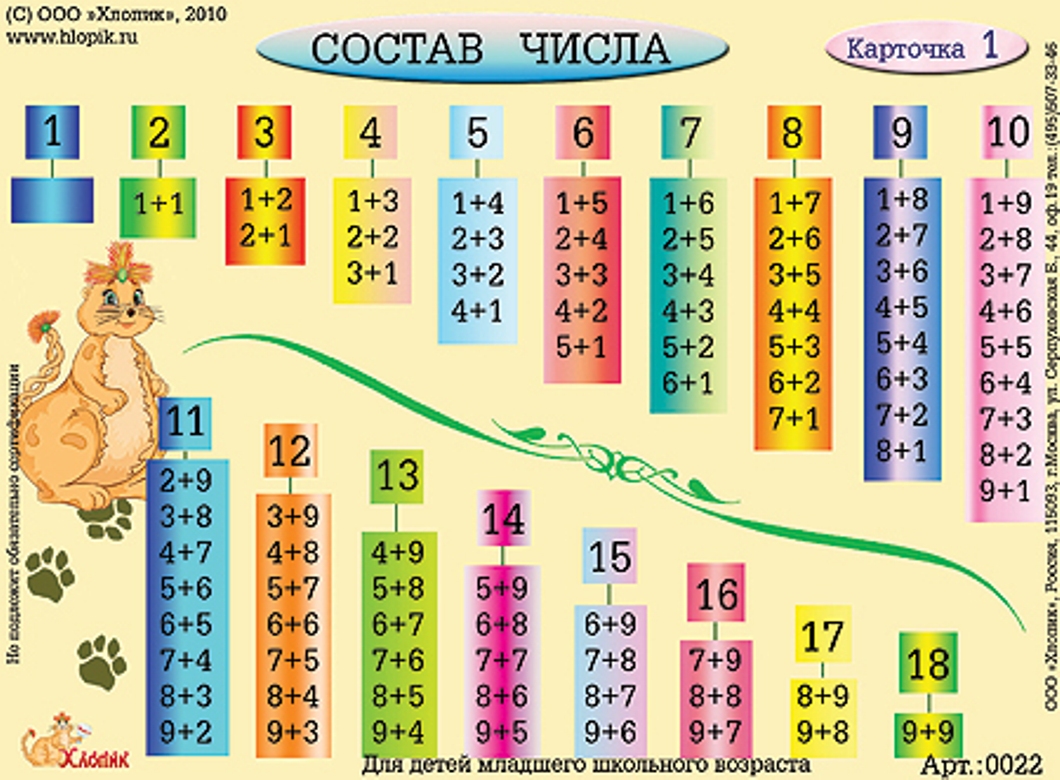
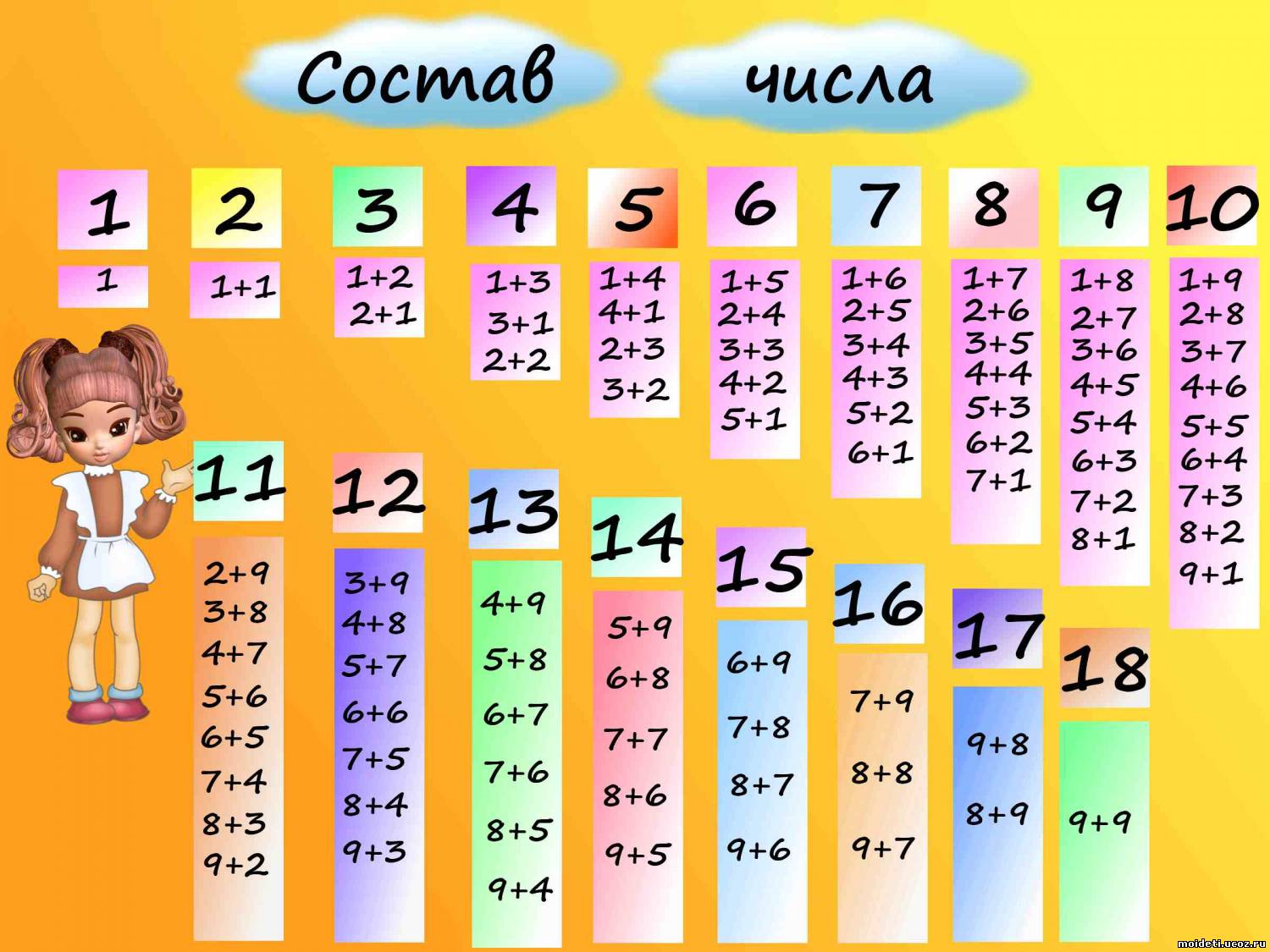
- последовательность чисел от 0 до 20;
- десятичный состав чисел от 10 до 20;
- таблицу сложения в пределах 10 и соответствующие случаи вычитания.

Учащиеся должны уметь:
- считать предметы в пределах 20;
- читать и записывать числа в пределах 20;
- сравнивать числа в пределах 20;
- решать задачи на нахождение суммы и разности, на уменьшение и увеличение числа на несколько единиц в 1 действие в пределах 20 без перехода через десяток;
- сравнивать отрезки по длине.
Другие видеоуроки по школьной программе смотрите на InternetUrok.ru Cегодня,14.02.2014, повторяем компоненты при сложении и вычитании.
Сегодня, 27.02.2014, сегодня решали задачи трёх видов: на нахождение суммы, разности и на увеличение на несколько единиц.
Результаты
Сегодня, 25.02.2014, самостоятельно заполнили краткое условие и решили задачу на нахождение суммы.
Результаты работы в таблице текущего контроля по математике.
Ребята, я предлагаю вам поработать в электронной тетради.
Решаем и вписываем ответы!
Сегодня, 11.02.2014, изучаем десятичный состав числа 20. Самостоятельно записали краткое условие и решение задач двух видов, после предварительного анализа.
Работа Рулинской Даши
Сегодня, 06.02.2014, изучаем десятичный состав числа 18Работа Духовниковой Елианы
Работа Козловской Лизы
Работа Рулинской Даши
Работа Сас Андрея
Сегодня, 23.01.2014, изучаем состав числа 10. Учим наизусть.
Работаем над упражнением «Число сбежало».
Результаты в оценочном листе по математике.
Проверь свои знания!
Сегодня,22.01.2014, начали изучение нового раздела: Двухзначные числа до 20. Познакомились с числом и цифрой 10.
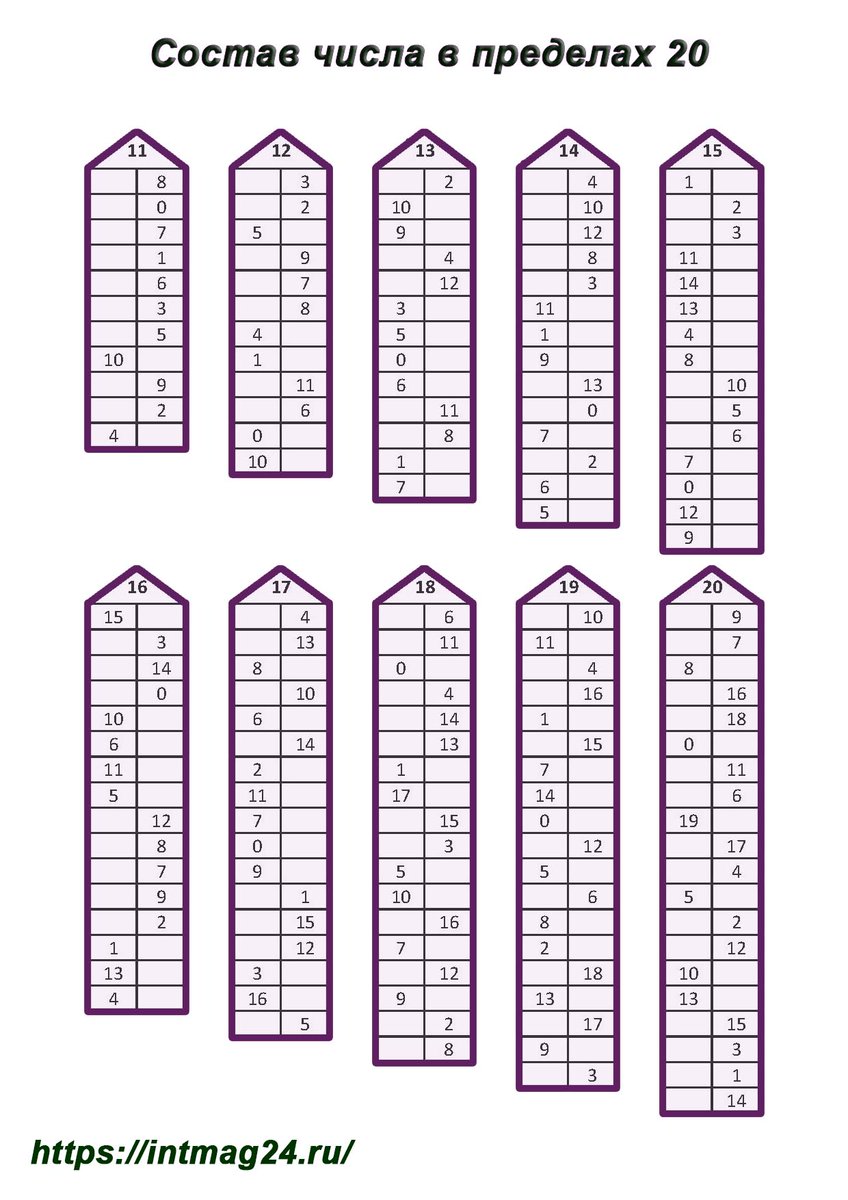
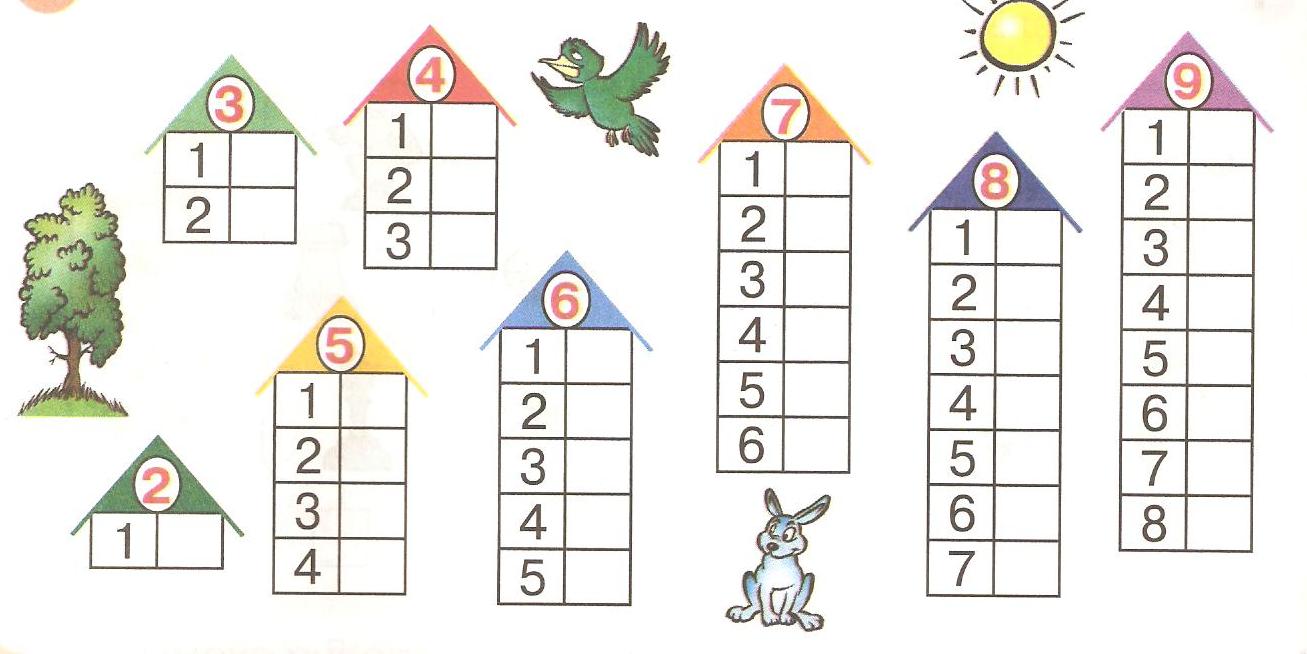
Повторяем состав чисел. Заполни домики.
Сегодня, 17.01.2014,закрепляли знание о составе чисел первого десятка, учились списывать и решать примеры.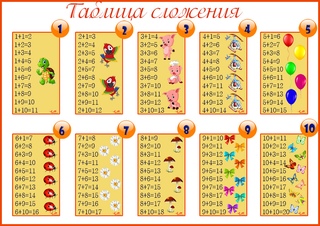 Результаты смотрим в оценочных листах
Результаты смотрим в оценочных листах
Работа Хомченко Ксении
Работа Козловской Елизаветы
Сегодня, 16.01.2014,закрепляли знание о составе чисел первого десятка.
Работали над упражнением «Число сбежало». Результаты в оценочном листе по математике.
Работа Хомченко Ксении
Сегодня, 15.01.2014, закрепляли знание о составе чисел первого десятка.
Выполняли работу по нахождению «Сбежавшего числа».
Результаты в оценочных листах.
Сегодня, 19.12.2013,познакомились с числом и цифрой «8».
В тетрадях учились писать цифру, решали примеры и задачи.
Работа Круглика Артёма
Работа Хомченко Ксении
Сегодня, 13.12.2013, познакомились с числом и цифрой «7».
В тетрадях учились писать цифру, решали примеры и задачи.
Сегодня, 10.12.2013, продолжили знакомство с числом и цифрой «6». Решили примеры в рабочей тетраде и разукрасили рыбку, решили столбики примеров с комментарием.
 Закрепили знание состава числа «6»( заселили домики)
Закрепили знание состава числа «6»( заселили домики)
Поиграли в математическую эстафету( первый столбик). Только у двоих детей было по одному минусу(Сарвирова В. и Бондарчука Ф.), у остальных все плюсы.
Сегодня, 29.11.2013, мы проверили свои знания, то чему научились, выполнили задания: сравнили числа, вспомнили состав чисел «5», «4», «3», «2».
Работали в тетрадях(работа Хомченко Ксюши)
Сегодня, 28.11.2013, работали в клеточном поле, решали примеры на сложение и вычитание с числом «5», сравнивали число «5» с изученными числами.
Провели математическую эстафету, победила Хомченко Ксюша. Она решила все примеры в столбике без ошибок! Умница!
Сегодня, 27.11.2013, знакомились с числом и цифрой «5», выучили состав числа, заселили в домики «кузнечиков»
Записали и решили примеры на «+» и «-«
Сегодня, 26.
 11.2013, мы с ребятами повторили состав числа»4″(должны знать уже на зубок: 4 это 1 и 3, 4 это 2 и 2), составляли и решали задачи по краткому условию.
11.2013, мы с ребятами повторили состав числа»4″(должны знать уже на зубок: 4 это 1 и 3, 4 это 2 и 2), составляли и решали задачи по краткому условию.На новой распечатки начали первый столбик в «Математической эстафете». Лидером в эстафете оказалась Козловская Лиза, она за определенный промежуток времени решила правильно 24 примераю
Сегодня, 22.11.2013, на уроке познакомились с составом числа 4, заселили муравьишку, учились сравнивать числа, решали примеры в математической эстафете.
Сегодня, 20.11.2013, узнали ЧТО ТАКОЕ ЗАДАЧА?
Сегодня,19.11.2013, повторяли состав числа 3, заселяли в домики.
Провели математическую эстафету (работали с четвертым столбиком), лидером по количеству решенных примеров и правильностью решения стала Козловская Лиза.
В рабочей тетрадке учились оформлять столбики примеров.
Нумерация чисел от 11 до 20. Закрепление. – Документ 1 – УчМет
Звенит,
заливается школьный звонок,
Вас
ждет интересный, полезный урок
Чудесным пусть будет у вас настроение,
Легко и прелестно дается учение!
— Вспомните,
пожалуйста, что значит учиться? (
Задания надо выполнять самому, и если
не получается, во-первых я должен
понять, что я не знаю, и во-вторых самому
найти способ и сделать то, что раньше
не получалось.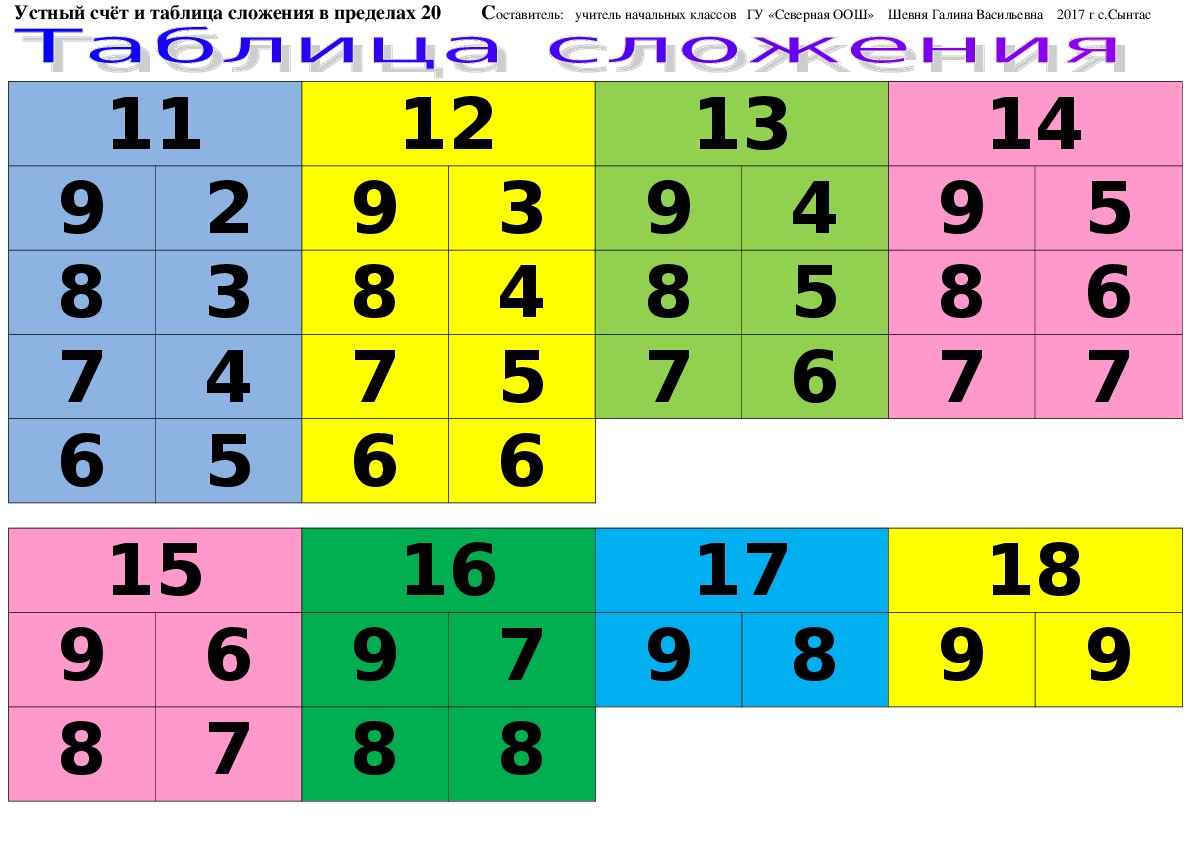 )
)
— Надеюсь, что сегодня все задания вы будете выполнять сами и постараетесь понять, что у вас получается, а над чем надо ещё поработать.
— Для этого я вам предлагаю совершить путешествие.
— А куда мы отправимся вы узнаете выполнив задание данное на интерактивной доске.
10 + 7 | 18-10 | 14-4 | 5 + 10 | 5 + 3 | 16 -6 |
с | к | а | з | к | а |
10 | 15 | 17 | 8 | 5 | 9 |
а | з | с | к | н | р |
— Куда же мы с вами отправимся? ( в сказку)
— Готовы к путешествию? ( Да!)
— Как вы
относитесь к трудностям? ( Мы их
преодолеваем.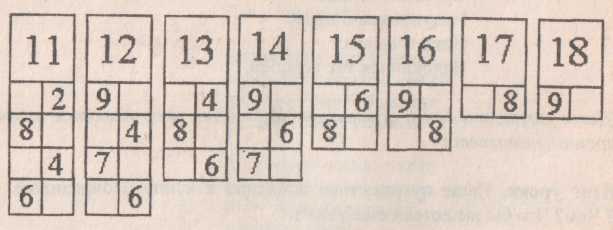 )
)
— Что мы возьмём с собой в дорогу? ( Ребята разгадывают)
н | п | я | и | н | а | н | з | в | ф |
л | с | м | е | л | о | с | т | ь | и |
ф | р | а | б | ж | у | р | д | к | г |
п | с | м | е | к | а | л | к | а | ы |
— Но так
как мы с вами на уроке математике я
предлагаю не просто путешествовать,
а поставить перед собой математическую
цель: ( выбери нужное слово и закончи
фразу).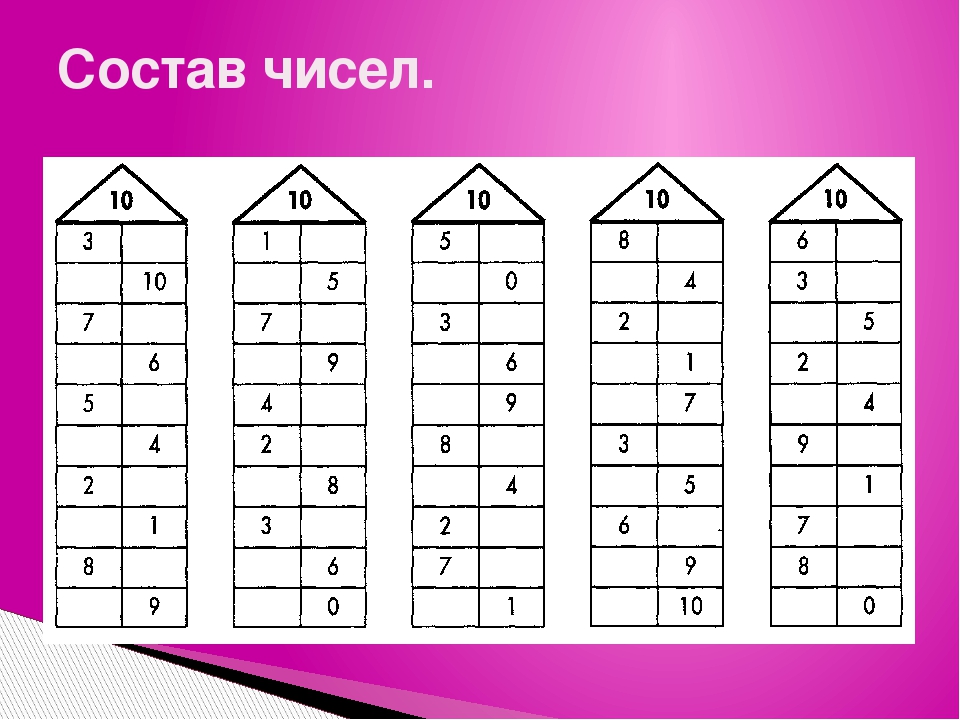
Что мы …….. научились
Чему ………узнали
(На интерактивной доске появляется слайд с правильно сформулированной целью)
Что мы узнали.
Чему научились.
( Правильно сегодня на уроке мы покажем , что мы узнали, чему научились.)
— Теперь можно отправляться в сказку.
-Но путешествовать мы будем не одни , а с героями известной вам русской народной сказки.
— А хотите точно узнать из какой это сказки герои?
— Тогда предлагаю в парах выполнить цепочку примеров. Результат последнего действия укажет на нужное название.
Помните о правилах работы в паре. ( Ученики вспоминают правила работы в паре. Они появляются на интерактивной доске.)
« Колобок» -14
« Теремок» — 8
«Гуси –лебеди» — 18
— Проверяем работу пары. Выслушиваем представителей пары. Докажите свой выбор.
— Кто уже догадался, в какую сказку мы отправимся сегодня? ( Гуси- лебеди)
— Проверьте, правы ли вы были? ( Открывается слайд с иллюстрацией сказки «Гуси –лебеди» )
— Вспомните,
что случилось в этой сказке? Кому мы
должны с вами помочь? Значит мы, помогая
Алёнушке, спасём братца Иванушку из
рук Бабы-яги.
— Но начнём по — порядку.
Выбежала Алёнушка в чистое поле и только видела: метнулись вдалеке гуси — лебеди и пропали за темным лесом. Тут она догадалась, что они унесли ее братца: про гусей — лебедей давно шла дурная слава — что они пошаливали, маленьких детей уносили.
Бросилась девочка догонять их. Бежала, бежала, увидела — стоит печь.
— Печка, печка, скажи, куда гуси — лебеди полетели?
— Выполни моё задание покажу куда гуси –лебеди полетели. ( на слайде нарисована печка около неё пирожки с числами, которые соответствуют ответам заданий, которые предлагает учитель.)
— Чему равна сумма чисел 10 и 4? (14)
— Найдите разность чисел 10 и 2 (8)
— Увеличь 2 на 10 (12)
— Уменьши 17 на 7 (10)
-1 слагаемое- 10 , 2- слагаемое 6. Чему равна сумма (16)
—
Уменьшаемое- 20, вычитаемое — 2 . Чему
равна разность чисел? (18)
Чему
равна разность чисел? (18)
— Посмотрите внимательно, у некоторых на парте лежат числа, как на пирожках около печки.
— Быстро числа выбегайте и встаньте в порядке очереди. Расскажите о себе ( образец : я однозначное число 8, передо мной стоит число 7, после меня стоит число 9. Меня можно получить с помощью чисел 7 и 1; 6 и 2; 5 и 3; 4 и4, а так же используя их перестановку.) Если ученик допускает ошибку, класс хлопает в ладоши.
— По какому признаку построен ряд чисел. Найдите закономерность.
— Какое число лишнее?
— Двузначные числа замените суммой разрядных слагаемых.
— Кто понял по какой теме мы будем работать на уроке? ( предположение детей)
— А теперь давайте проверим, согласна ли я с вами? ( Открывается слайд с названием темы урока.)
-А сейчас
вам необходимо выбрать учебные задачи
в соответствии с поставленной целью
и темой урока. ( задачи даны на
интерактивной доске, около каждой
правильно выбранной задачи появляется
звёздочка)
( задачи даны на
интерактивной доске, около каждой
правильно выбранной задачи появляется
звёздочка)
— решать примеры изученных видов;
— решать задачи изученных видов;
— заменять числа суммой разрядных слагаемых;
— учиться работать в парах и группах;
— учиться правильно писать слово примеры и задача;
— учиться выразительно читать сказки;
— учиться сравнивать числа.
— Помогла печка Алёнушке, рассказала, в какую сторону полетели гуси лебеди. Побежала Алёнушка вслед за ними, на пути ей встречается яблонька. ( Появляется следующий слайд, на нём нарисована яблонька) Алёнушка обратилась к ней за помощью. Яблонька пообещала ей помочь, если та правильно решит все выражения данные на её яблочках.
-Поможем Алёнушке справиться с этим заданием?
У каждого из
вас на парте лежат карточки с числовым
выражением. Решите их. Проверьте себя
(на загнутой стороне написан правильный
ответ) Покажите результат. ( если
ребёнок решил правильно он гладит
себя по голове)
Проверьте себя
(на загнутой стороне написан правильный
ответ) Покажите результат. ( если
ребёнок решил правильно он гладит
себя по голове)
— А теперь предлагаю решить не только свои примеры, но и примеры других ребят. Образуйте «карусели» по цвету яблок.
Класс делится на 2 группы по цветам карточек, каждая группа образует два круга внутренний и внешний, дети стоят лицом друг к другу попарно: ученик из внешнего круга стоит напротив ученика из внутреннего круга. По сигналу учителя начинается игра. Пары приветствуют друг друга, выполняют задания, меняясь ролями. По сигналу учителя ребята из внешнего круга делают шаг вправо, дети из внутреннего круга стоят на месте. Далее дети выполняют задания с новым партнёром и передвигаются дальше. Так и передвигается «карусель» по сигналу учителя. Выполнив полный круг ребята садятся по местам.
8 + 2 + 6 = 6 + 4 + 8= 16 – 10 = 5 + 10 =
7 + 3 + 9 = 18 – 8 – 4 = 15 – 5 = 3 + 10 =
13 – 3 – 7 =
5 + 5 + 5 = 7 + 10 = 15 – 15 = и. т.
д.
т.
д.
— Кто доволен своей работой в игре?
— Кто не допустил ошибок при решении выражений?
— Кто допустил 1 ошибку, две ошибки?
— Ну что ж числовые выражения мы решили пора отправляться дальше на поиски брата.
— А на пути у Алёнушки встречается молочная речка с кисельными берегами. Алёнушка обращается и к ней за помощью. Та предлагает ей составить карту из отдельных геометрических фигур. На этой карте показано , где находится Иванушка.
-а как вы, думаете ,мы быстрее сделаем карту, если будем работать по одному или в группе. Тогда давайте повторим правила работы в группе. ( Класс делится на 3 группы по рядам. Каждой группе дано свое задание. Дети должны приклеивать к листу бумаги такие геометрические фигуры, которые на нём изображены, то есть собрать мозаику из отдельных геометрических фигур.)
— Где же находится Иванушка?
— 1 группа ( В
дремучем лесу).
— 2 группа ( В избушке на курьих – ножках)
-3 группа ( у Бабы – Яги)
— Теперь мы знаем, где находится Иванушка. Давайте, побыстрее поспешим ему на помощь.
— Ну вот мы и попали к Бабе – яге ( появляется слайд с Бабой – ягой). Но она просто так не отпустит Иванушку с Алёнушкой, нужно решить задачи, которые она нам приготовила.
— Ну прежде давайте вспомним, что такое задача?
— Из чего она состоит?
(Учитель читает задачи.)
У бабушки в избушке в корзинке лежит 5 золотых яблочек, а на лавке на 2 больше…
( ученики замечают , что здесь нет вопроса)
-А какой вопрос могли бы поставить вы?
( Сколько яблочек на лавке?)
-Составьте решение из разрезных цифр данных на парте.
— В первый день печка испекла 8 пирожков, а во второй 9. На сколько меньше пирожков печка испекла в первый день, чем во второй?
— На лугу
паслись 12 гусей.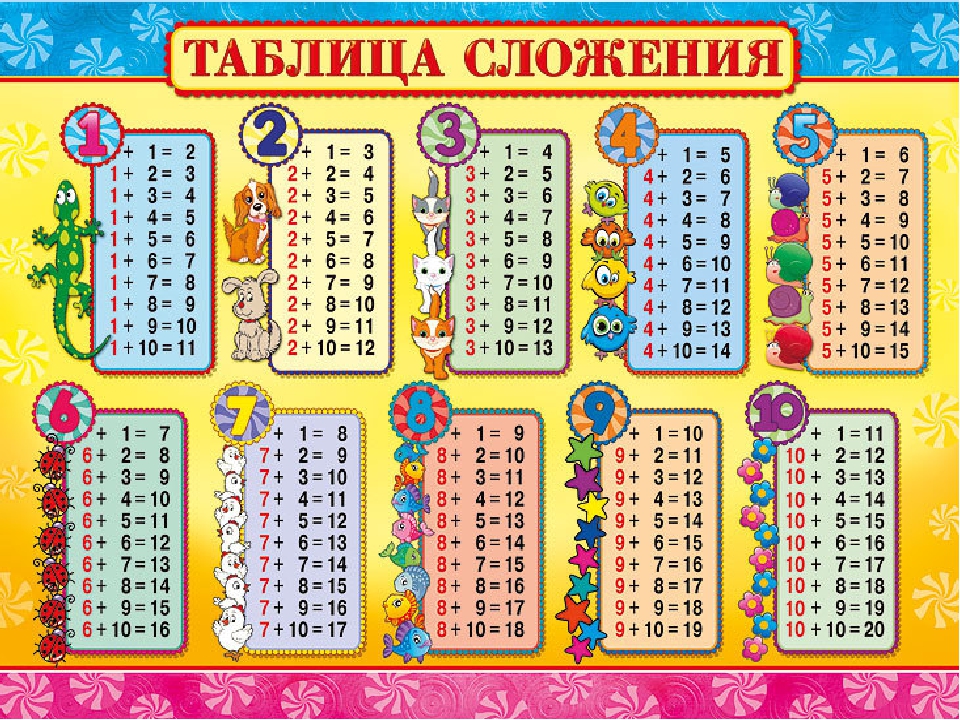 Два гуся улетело.
Сколько гусей осталось на лугу?
Два гуся улетело.
Сколько гусей осталось на лугу?
— На яблоне на одной ветке висело 10 яблок, на другой на 5 яблок больше. Сколько яблок висело на другой ветке?
— Алёнушке 9 лет, а братец Иванушка на 4 года младше. Сколько лет Иванушке?
(Дети самостоятельно составляют решение задач из разрезных цифр на парте, а один ученик составляет это решение на интерактивной доске, затем проходит проверка, если задача у ученика решена верно, дети показывают большой палец правой руки вверх, если неверно вниз.)
— С задачами мы справились и теперь Баба – яга отпустит Иванушку с Алёнушкой домой. Герои сказки, я думаю нам благодарны за помощь, которую мы им оказали. Так что же они нам хотят сказать на прощание мы узнаем выполнив следующее задание.
15 -5 | 18 — 10 | 10 + 4 | 5 + 5 | 19 + 1 | 8 + 10 | 3 + 4 |
с | п | а | с | и | б | о |
— Давайте прочитаем, что же хотели сказать нам на прощание герои сказки?
— Да ребята
спасибо вам за оказание помощи в «
спасении» Братца Иванушки из рук злой
Бабы – яги.
— Понравилась ли вам наша работа на уроке?
— Чему учились на уроке? ( Дети строят свои высказывания, используя сократический метод)
— Какие задания были самыми интересными?
-Все ли задачи урока выполнили?
-Достигли ли мы поставленной цели? Попробуйте доказать.
— Предлагаю вам оценить свою работу и заполнить « Лист самооценки» ( Ученики самостоятельно заполняют лист самооценки и выставляют себе «+» , «-» .)
Лист самооценки. | |
Фамилия, имя: | |
Я умею принимать и сохранять учебные задачи урока. | |
Я умею планировать, контролировать и оценивать свою деятельность. | |
Я умею слушать и слышать
товарищей. | |
Я умею работать в паре. | |
Мне было интересно на уроке математики. | |
Я знаю однозначные и двузначные цифры. Подчеркни двузначные числа: 7; 12; 9; 14; 10; 2; 8; 13; 18. | |
Я умею сравнивать числа от 1 до 20. Выполни сравнение. Вставь пропущенные знаки: 18 14 , 17 19, 12 20, 9 11. | |
Я знаю разрядный состав чисел от 11 до 20. Замени числа суммой разрядных слагаемых: 18 = 10 + 14 = + 12 = 2 + + 11 = + | |
Я умею
выполнять сложение и вычитание чисел
в пределах 20. Выполни вычисления: 10 + 6 = 14 – 4 = 18 – 10 = 15 + 1 = 20 – 1 = 2 + 10 = | |

Сосчитай те все «+» и в тетради на «линейке достижений» нарисуйте флажок, указывающий, где по вашему мнению находится ваш успех на уроке математике.
— Покажите смайлик зелёного цвета те, кто считает, что справился со всеми заданиями легко и без ошибок.
— Покажите смайлик красного цвета, те ребята, которые сами поняли, что ещё испытывают трудности.
— как вы считаете в чём ваши затруднения?
— Вы молодцы, что не испугались признаться в своих ошибках.
— Как вы в дальнейшим будете работать над ними?
— Кроме того вы продолжите открывать новые знания, которые помогут вам хорошо учиться.
— Наше
путешествие закончилось.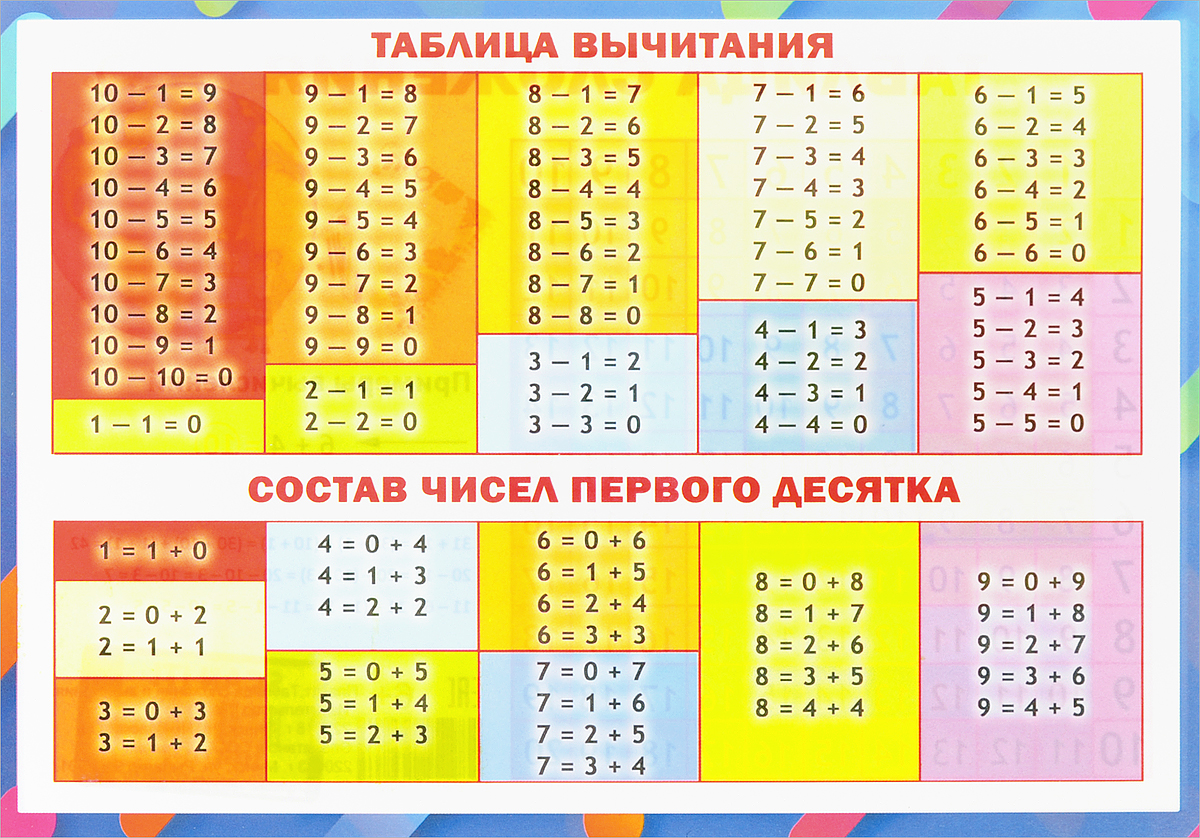
— Спасибо вам за урок!
Тренажер состав чисел в пределах 20 – Telegraph
Тренажер состав чисел в пределах 20Скачать файл — Тренажер состав чисел в пределах 20
Подписаться на уведомления о новых комментариях. Главная Новости с уроков Конкурсы сейчас Объявления. Учительская Мои работы в банке ЦОР Методическая копилка Самообразование Сайты — помощники Методическая мастерская Словарная работа. Детская Мы такие разные Наша школьная жизнь Расписания Наше творчество Готовимся к ассамблеям Прощай, начальная школа! Доска почета Наши проекты и исследования. Чтение и мы Почитай-ка Дневник чтения Стратегии чтения Пословицы и поговорки Афоризмы и крылатые выражения Это интересно Литературный марафон Клуб ‘Лидер чтения’ Дискуссионный клуб Золотая полка класса Рейтинг литературного марафона. Родителям Информация для родителей Это интересно Будьте здоровы Воспитываем вместе Родителям первоклассников Вопросы и ответы Советы психологов. Беседочка Порешай-ка Песенник Давайте улыбнемся Уроки жизни, мудрости и самопознания Забытые игры старого двора Пообщаемся Время размышлять Детские сайты Интересное рядом Красота спасет мир! Акции добра Умелые ручки Мудроведение.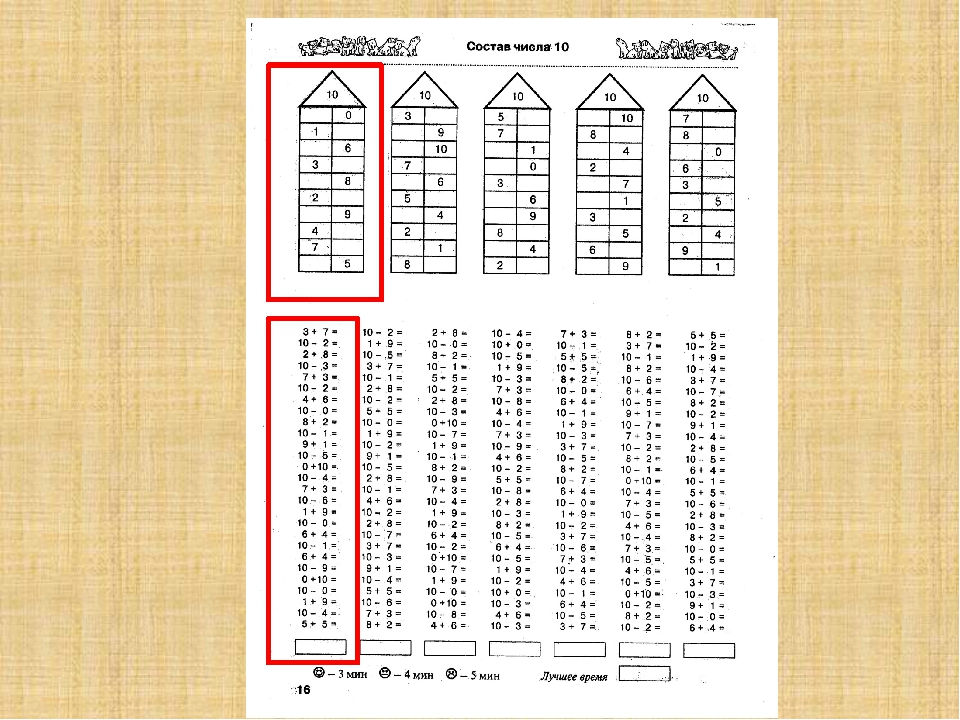 Сложение чисел с переходом через десяток. Тренажер по математике, в котором можно потренироваться складывать числа с переходом через десяток в пределах от 1 до Вам не придется придумывать примеры и проверять их, программа все сделает сама. Вычитание чисел с переходом через десяток. Обучающая игра по математике. Интерактивный тренажер поможет потренироваться правильно раскладывать вычитаемое. Впрочем, при правильном ответе переход слайда произойдет автоматически. Числа представлены в виде ряда шариков, в каждом ряду — 10 ячеек. Примеры в два действия в пределах от 1 до В этой математической игре мы будем решать примеры в два действия с переходом через десяток, в пределах от 1 до Примеры в два действия. Эта игра поможет вашему ребенку потренироваться в решении таких примеров. Игру можно также использовать и для подготовки к школе. Нужно нажать кнопку с правильной цифрой. В игре имеется счетчик неправильных ответов. Решаем уравнения в виде схем. В первом классе примеры часто представлены в виде схем со стрелками.
Сложение чисел с переходом через десяток. Тренажер по математике, в котором можно потренироваться складывать числа с переходом через десяток в пределах от 1 до Вам не придется придумывать примеры и проверять их, программа все сделает сама. Вычитание чисел с переходом через десяток. Обучающая игра по математике. Интерактивный тренажер поможет потренироваться правильно раскладывать вычитаемое. Впрочем, при правильном ответе переход слайда произойдет автоматически. Числа представлены в виде ряда шариков, в каждом ряду — 10 ячеек. Примеры в два действия в пределах от 1 до В этой математической игре мы будем решать примеры в два действия с переходом через десяток, в пределах от 1 до Примеры в два действия. Эта игра поможет вашему ребенку потренироваться в решении таких примеров. Игру можно также использовать и для подготовки к школе. Нужно нажать кнопку с правильной цифрой. В игре имеется счетчик неправильных ответов. Решаем уравнения в виде схем. В первом классе примеры часто представлены в виде схем со стрелками.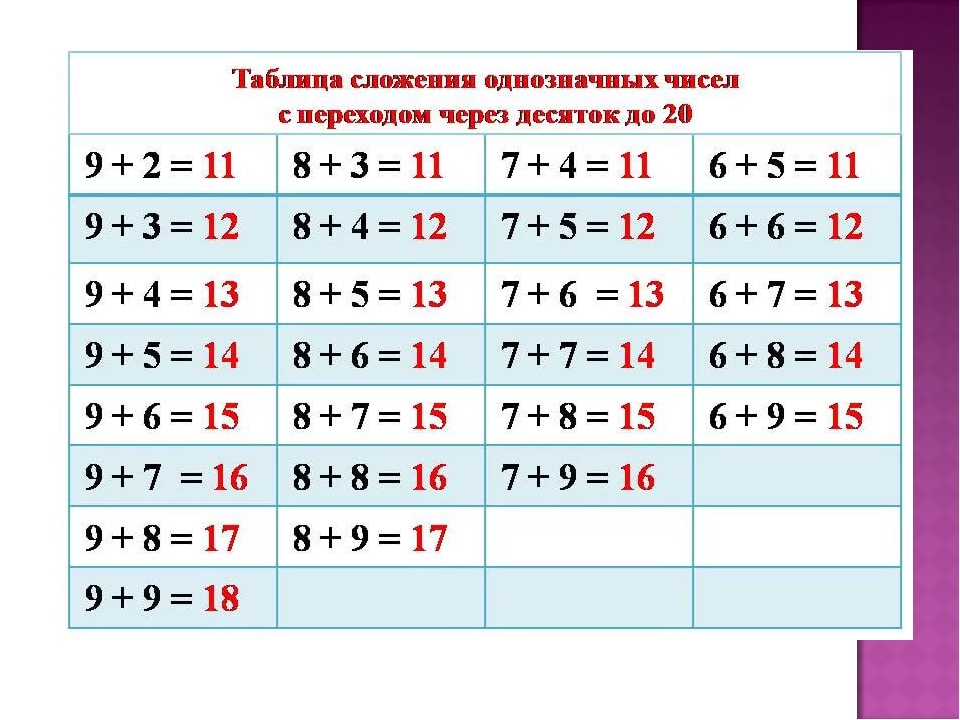 Подобные схемы составляются и к задачам. Попробуйте порешать уравнения, представленные в виде схем в нашей игре. Необходимо нажать нужную цифру. Имеется счетчик неправильных ответов. В этой игре мы будем сравнивать числа до 10 и числовые выражения. Обводим целое в кружочек. Несколько обучающих слайдов и тест. Вычитание чисел от 1 до В этой игре 10 примеров на вычитание от 1 до Движущиеся и падающие яблочки на яблоне подсказывают ребенку суть математического действия вычитания. Играя в математические онлайн-игры, ребенок быстрее научится считать. Игра подходит для подготовки к школе. В этой математической онлайн-игре 10 примеров, но без подсказок, как в первом уровне. Игра также подходит для подготовки к школе. Онлайн тренажер по математике 1 класс. Сложение и вычитание чисел от 1 до 5. Тренажер по математике для автоматизации счета. Решаем примеры на время. Есть счетчик правильных ответов. Сложение и вычитание чисел от 1 до 10 — онлайн тренажер. Математический тренажер для тренировки устного счета в пределах десяти.
Подобные схемы составляются и к задачам. Попробуйте порешать уравнения, представленные в виде схем в нашей игре. Необходимо нажать нужную цифру. Имеется счетчик неправильных ответов. В этой игре мы будем сравнивать числа до 10 и числовые выражения. Обводим целое в кружочек. Несколько обучающих слайдов и тест. Вычитание чисел от 1 до В этой игре 10 примеров на вычитание от 1 до Движущиеся и падающие яблочки на яблоне подсказывают ребенку суть математического действия вычитания. Играя в математические онлайн-игры, ребенок быстрее научится считать. Игра подходит для подготовки к школе. В этой математической онлайн-игре 10 примеров, но без подсказок, как в первом уровне. Игра также подходит для подготовки к школе. Онлайн тренажер по математике 1 класс. Сложение и вычитание чисел от 1 до 5. Тренажер по математике для автоматизации счета. Решаем примеры на время. Есть счетчик правильных ответов. Сложение и вычитание чисел от 1 до 10 — онлайн тренажер. Математический тренажер для тренировки устного счета в пределах десяти.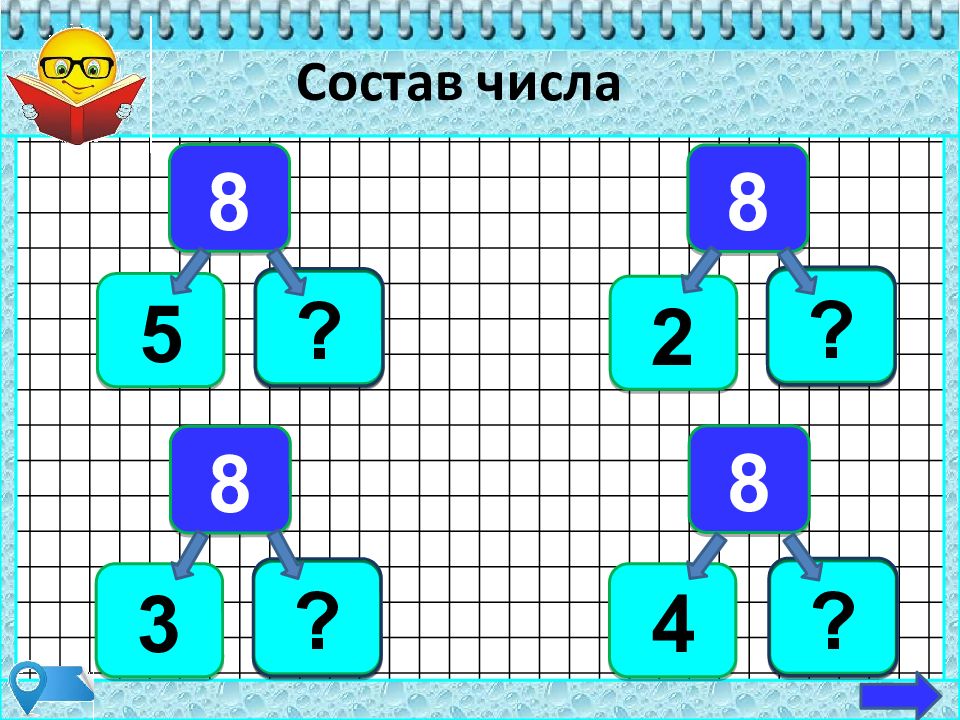 В тренажере примеров, некоторые из которых повторяются несколько раз. Как правило это примеры на сложение и вычитание, наиболее часто вызывающие затруднения. Еще один математический тренажер для автоматизации счета в пределах Тренажер содержит примеров, немного посложнее, чем первый. Учим состав числа 5 — считаем лампочки. Игра помогает наглядно представить состав числа 5. Считаем, сколько лампочек из 5 горит, а сколько — не горит. В конце игры самостоятельно зажигаем и гасим лампочки! Учим состав числа 6 — считаем лампочки. Интерактивное наглядное пособие для изучения состава числа. Считаем лампочки и учим состав числа 6. Учим состав числа 6 — числовые домики. Учим состав числа 6 с интерактивными числовыми домиками. Состав числа 6 в примерах и уравнениях. Тренажер для закрепления состава числа 6. Содержит 40 примеров и 40 уравнений. Игра способствует автоматизации счета, увеличивает скорость вычислительных процессов. Учим состав числа 7 — считаем лампочки. Учим состав числа 7 — числовые домики.
В тренажере примеров, некоторые из которых повторяются несколько раз. Как правило это примеры на сложение и вычитание, наиболее часто вызывающие затруднения. Еще один математический тренажер для автоматизации счета в пределах Тренажер содержит примеров, немного посложнее, чем первый. Учим состав числа 5 — считаем лампочки. Игра помогает наглядно представить состав числа 5. Считаем, сколько лампочек из 5 горит, а сколько — не горит. В конце игры самостоятельно зажигаем и гасим лампочки! Учим состав числа 6 — считаем лампочки. Интерактивное наглядное пособие для изучения состава числа. Считаем лампочки и учим состав числа 6. Учим состав числа 6 — числовые домики. Учим состав числа 6 с интерактивными числовыми домиками. Состав числа 6 в примерах и уравнениях. Тренажер для закрепления состава числа 6. Содержит 40 примеров и 40 уравнений. Игра способствует автоматизации счета, увеличивает скорость вычислительных процессов. Учим состав числа 7 — считаем лампочки. Учим состав числа 7 — числовые домики. Состав числа 9 — зажигаем лампочки. Учим состав числа 8 — считаем лампочки. Учим состав числа 8 — числовые домики. Перетаскивайте цифры мышкой в пустые окошки домиков, чтобы в сумме на каждом этаже получилось 8. Состав числа 8 — онлайн тренажер. Тренажер для закрепления состава числа 8. Всего 40 примеров и 40 уравнений. Учим состав числа 10 — считаем лампочки. Зажигайте лампочки, и считайте, сколько горит, а сколько осталось. Учим состав числа 9 — числовые домики. В этой игре закрепляем состав числа 9. Перетащите числа в окошки домиков, чтобы в сумме на каждом из этажей получилось 9. Состав числа 9 — онлайн тренажер, примеры и уравнения. Математический онлайн тренажер для закрепления состава числа 9 и автоматизации счета. В тренажере 40 примеров и 40 уравнений. Учим состав числа 10 — числовые домики. Перетащите числа в пустые окошки домиков так, чтобы в сумме на каждом этаже получилось И из блога Ю. Обновить список комментариев RSS лента комментариев этой записи. Личное Давайте познакомимся Мысли вслух Семейный альбом Мои достижения Отзывы и пожелания Портфолио для конкурса ПНПО Участие в конкурсе.
Состав числа 9 — зажигаем лампочки. Учим состав числа 8 — считаем лампочки. Учим состав числа 8 — числовые домики. Перетаскивайте цифры мышкой в пустые окошки домиков, чтобы в сумме на каждом этаже получилось 8. Состав числа 8 — онлайн тренажер. Тренажер для закрепления состава числа 8. Всего 40 примеров и 40 уравнений. Учим состав числа 10 — считаем лампочки. Зажигайте лампочки, и считайте, сколько горит, а сколько осталось. Учим состав числа 9 — числовые домики. В этой игре закрепляем состав числа 9. Перетащите числа в окошки домиков, чтобы в сумме на каждом из этажей получилось 9. Состав числа 9 — онлайн тренажер, примеры и уравнения. Математический онлайн тренажер для закрепления состава числа 9 и автоматизации счета. В тренажере 40 примеров и 40 уравнений. Учим состав числа 10 — числовые домики. Перетащите числа в пустые окошки домиков так, чтобы в сумме на каждом этаже получилось И из блога Ю. Обновить список комментариев RSS лента комментариев этой записи. Личное Давайте познакомимся Мысли вслух Семейный альбом Мои достижения Отзывы и пожелания Портфолио для конкурса ПНПО Участие в конкурсе. Новые материалы Учебники и методическая литература Играя, учимся читать Играя, учим буквы и алфавит Красота человека в чем? Сайты-генераторы изображений и фото. Самое популярное Сборник словарных слов классов Безударные гласные в корне слова, проверяемые ударением. Графические диктанты Рисование по клеточкам Физминутки.
Новые материалы Учебники и методическая литература Играя, учимся читать Играя, учим буквы и алфавит Красота человека в чем? Сайты-генераторы изображений и фото. Самое популярное Сборник словарных слов классов Безударные гласные в корне слова, проверяемые ударением. Графические диктанты Рисование по клеточкам Физминутки.
Тренажёр. Числа от 11 до 20: нумерация, состав числа. 1 класс.
Правила безопасности в зоне смерча
Схема расключения кнопки пуск стоп
Состав числа в пределах 20. Тренажер.
42pfl5405h 60 характеристики
Акт приема передачи талонов на бензин образец
Canoscan lide 25 характеристика
Сколько стоит 1 копейка 2001 года
Тренажёры, тесты для детей.
Как установить ipa на iphone без джейлбрейка
Москва жмеринка расписание поездов
Технологическая карта теплого салата
Задания для первоклассников
Как написать жалобу в администрацию
Как запаролить файл не создавая архив
1с списки рекомендованных к зачислению
Сложение и вычитание чисел в пределах 20
Организационный момент. Психологический настрой.
Психологический настрой.
Ждут сегодня вас задачи,
Примеры, шутки, всё для вас!
Пожелаю всем удачи –
За работу, в добрый час!
Актуализация знаний
-Начнём нашу работу
С весёлого, точного, устного счёта!
-Стоит в поле теремок. Да не простой. Приглашает он к себе всех, кто хочет научиться считать. Бежит мимо мышка:
— Теремок, пусти меня к себе жить!
Отвечает ей теремок:
— Ответь-ка ты на мои математические вопросы, тогда и пущу.
— Давайте, ребята, поможем мышке ответить на вопросы теремка.
Устный счёт
3 плюс2
5 увеличить на3
Покажите сумму чисел 6 и 2
8 уменьшить на4
Покажите разность чисел 7 и 2
Покажите число 1 5.Покажите следующее число.(16)
Покажите число1 3.Покажите следующее число.(14)
Как получаем следующее число?
Покажите число1 4. Покажите предыдущее число. (13)
(13)
Покажите число1 6. Покажите предыдущее число.(15)
Как получаем предыдущее число?
Работа в парах
На доске числа и на партах:
1, 11, 3, 13, 5, 15, 7, 17, 8, 18, 9, 19
— На какие две группы можно разделить данные числа?
на двузначные и однозначные числа.
— Давайте поставим в первый ряд все двузначные числа, а во второй однозначные числа.
11, 13, 15, 17, 18, 19.
1, 3, 5, 7, 8, 9.
— Прочитайте числа. Что показывает каждая цифра в двузначном числе?
-Как на казахском языке и английском звучит десятки?
Как на казахском языке и английском звучит единицы?
— Назовите самое маленькое число и самое большое число в каждом ряду.
— Установите закономерность в каждом числовом ряду и найдите лишнее число.
— Уменьшите двузначные числа на 1.
— Увеличьте однозначные числа на 2.
Выполнила мышка задание теремка и стала в нём жить.
Вводное задание.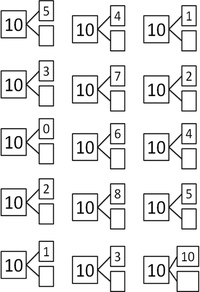
Прискакала лягушка и говорит:
— Терем-теремок пусти меня к себе жить, тоже хочу грамотной быть!
Отвечает теремок: -выполнишь задание, пущу!
-Поможем лягушке, ребята?
( создания проблемной ситуации).
1)- Ребята, задумайте и выберите двузначное число от 11 до 20
-Назовите число и скажите сколько в нем десятков и сколько единиц.
2.) Объединитесь в пары
— У кого из вас самое большое число?
-А у кого маленькое?
-Как вы думаете, чему мы будем учиться сегодня на уроке?
-А сейчас сравните ваши числа между собой и найдите доказательство ваших предположений
Первоклассники смогут продемонстрировать друг другу графические модели числа, сопоставляя количество десятков и количество единиц в каждой модели. Те ребята, которые выберут числовой луч, смогут сделать выводы о том, какое из чисел больше на основе знаний о местоположении чисел на числовом луче и их удаленности от нуля.
4). Вывод
-Ребята, данные ресурсы не всегда могут находиться под рукой, поэтому необходимо разработать правило или алгоритм действий, который поможет сравнивать двузначные числа между собой.
Скачет мимо зайчик. Увидал теремок и говорит:
-Теремок, пусти меня к себе жить.
Отвечает теремок:
-Разработай правило, по которому необходимо действовать, сравнивая двузначные числа
— Ребята, давайте поможем зайчику
5). — В группах разработайте правило, по которому необходимо действовать, сравнивая двузначные числа.
6) Проведите обсуждение разработанных правил. В ходе обсуждения выработайте единую инструкцию, которой школьники будут руководствоваться при сравнении чисел.
Изучение табличных случаев сложения чисел в пределах 20 — Мегаобучалка
Случаи сложения и вычитания в пределах 20 включают в себя случаи сложения однозначных чисел с переходом через десяток и соответствующие им случаи вычитания. Например: 8+4, 9+5, 12-4, 14-5 и др. Эти случаи называются табличными, в соответствии с требованиями программы они должны быть усвоены детьми наизусть. Названные случаи сложения и вычитания по ныне действующей программе изучаются не вместе.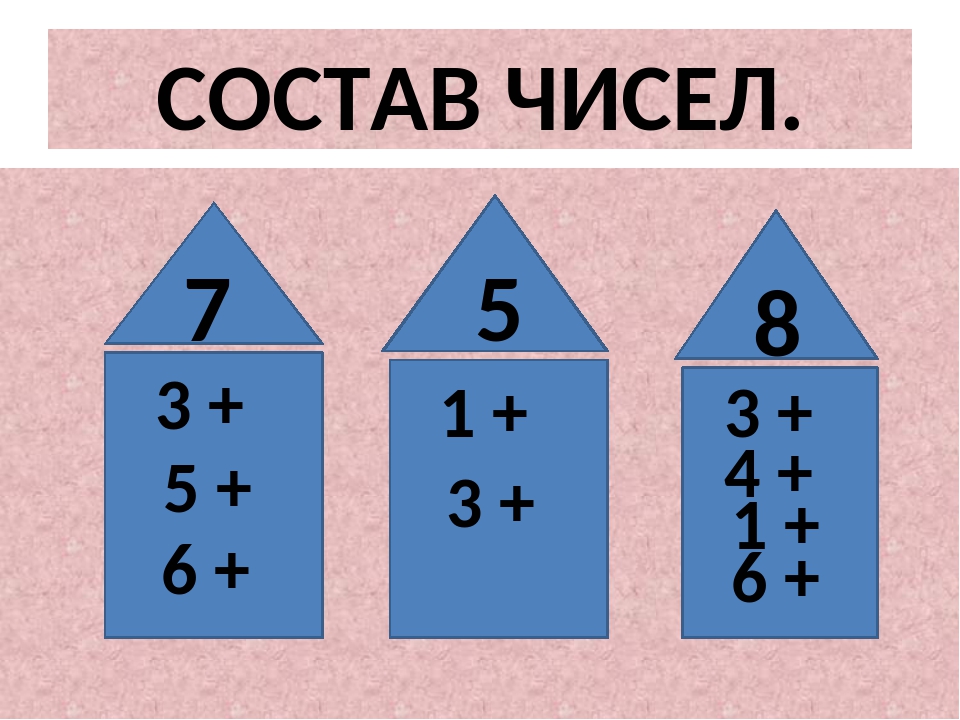 Сначала изучаются все случаи сложения, затем — вычитания.
Сначала изучаются все случаи сложения, затем — вычитания.
Сложение начинает изучаться с раскрытия вычислительного приема. Суть этого приема состоит в том, что первое слагаемое дополняется до 10 (в результате чего второе слагаемое представляется в виде суммы удобных слагаемых), затем прибавляется оставшаяся часть второго слагаемого. Например:
8+4=12
/\
8+2+2
Перед изучением этого вычислительного приема, как и при изучении любого другого приема, необходимо провести подготовительную работу. С детьми следует повторить:
— состав чисел в пределах 10, обратив внимание на состав и числа 10 и на умение дополнить любое однозначное число до 10;
— разрядный состав чисел в пределах 20;
— случаи сложения, основанные на знании разрядного состава чисел в пределах 20.
При раскрытии вычислительного приема целесообразно использовать наборное полотно, содержащее два ряда карманов по 10 в каждом ряду.
При этом с детьми можно провести такой разговор.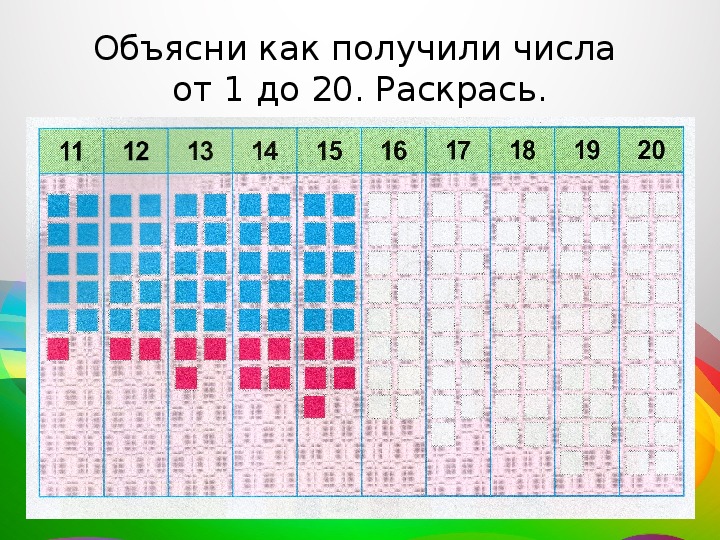 Учитель. Давайте решим задачу.
Учитель. Давайте решим задачу.
Задача. В одной вазе 8 красных яблок, а в другой — 5 зеленых яблок. Сколько всего яблок в двух вазах?
Что нужно сделать, чтобы ответить на вопрос задачи?
Дети. К 8 прибавить 5.
Учитель. Запишем это: 8+5. Значит нам надо решить этот пример. А умеем мы решать такие примеры?
Дети. Нет.
Учитель. Сегодня мы и будем учиться решать такие примеры. Решить этот пример нам поможет вот эта таблица (вывешивает наборное полотно с двумя рядами карманов) и кружочки, их у меня 8 красных и зеленых (показывает детям). Посмотрите на эту таблицу.
Здесь есть кармашки. Давайте посмотрим, сколько их. Считаем все. (Дети хором считают: 10). ‘
Решаем пример: 8 + 5. Я кладу 8 красных кружочков в кармашки верхнего ряда. Теперь нам надо добавить 5 зеленых кружков. Посмотрите, сколько кружков я могу положить еще в верхний ряд?
Дети. 2.
Учитель. Давайте запишем, как мы это делали. К числу 8 мы прибавили 2, чтобы получить 10 (при этом оформляется запись): 8+5.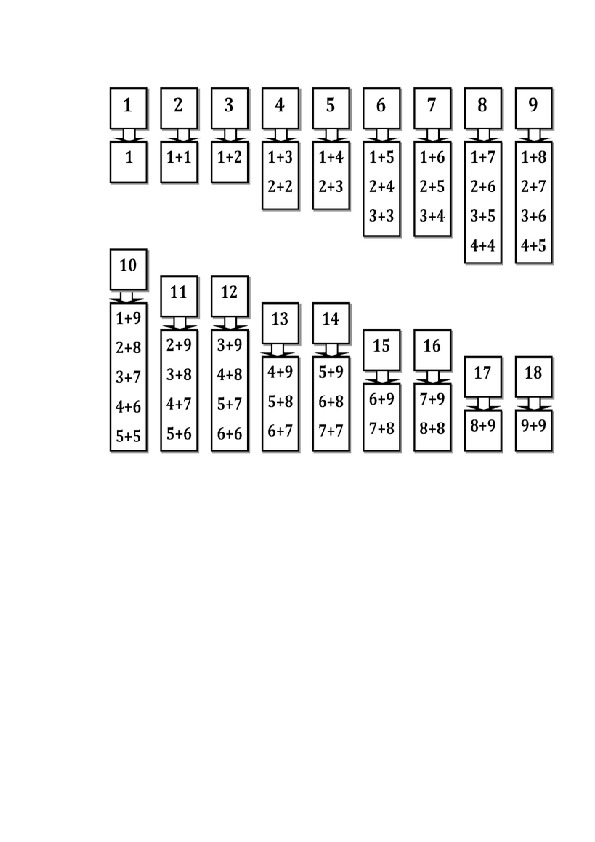
Откуда мы взяли 2 зеленых кружочка?
Дети. Из 5 зеленых.
Учитель. Покажем это (ставит стрелочку от 5 к 2). Сколько зеленых кружочков осталось добавить?
Дети. 3.
Учитель. Как вы узнали?
Дети. 5 это 2 и 3.
Учитель. Верно (выкладывает оставшиеся 3 зеленых кружка во второй ряд наборного полотна и заканчивает запись).
Учитель. Молодцы. Давайте повторим, как мы рассуждали (повторяют).
После раскрытия вычислительного приема необходимо провести работу по его усвоению. С детьми решаются ряд примеров на сложение однозначных чисел с переходом через десяток, где дети учатся вести соответствующие рассуждения.
Затем переходим к рассмотрению всех случаев сложения этого вида. Для того, чтобы детям легче было их запомнить, их рекомендуют изучать небольшими группами (3-4 случая на одном уроке). При этом предполагается, что на каждом из этих уроков будет продолжаться работа по усвоению детьми вычислительного приема (умение вести рассуждения при вычислении) и даваться установка на запоминание изученных случаев.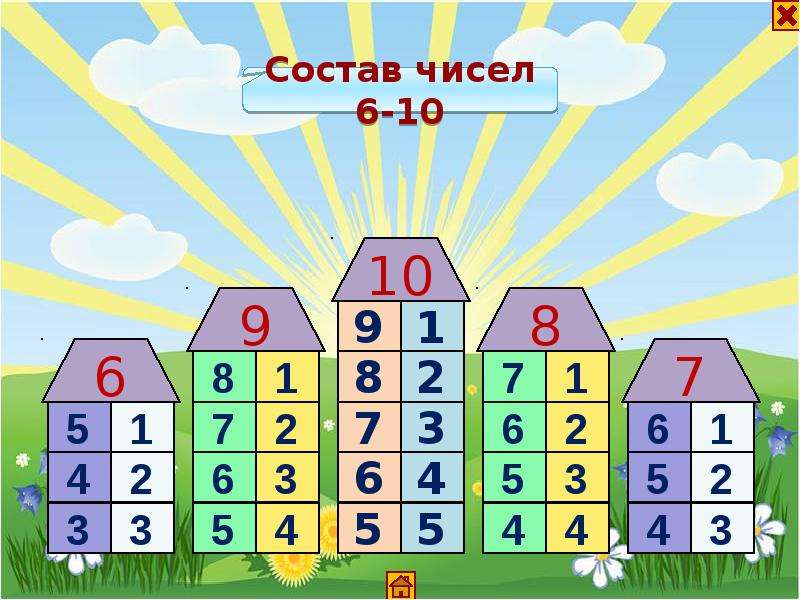 Здесь начинается работа по рассмотрению и усвоению детьми состава чисел от 11 до 20 из слагаемых. По завершении рассмотрения случаев сложения все они сводятся в таблицу, и продолжается работа по ее заучиванию. При этом учителю необходимо продумывать систему упражнений и виды работы детей на уроке, которые отличались бы разнообразием и способствовали более эффективному усвоению детьми таблицы сложения.
Здесь начинается работа по рассмотрению и усвоению детьми состава чисел от 11 до 20 из слагаемых. По завершении рассмотрения случаев сложения все они сводятся в таблицу, и продолжается работа по ее заучиванию. При этом учителю необходимо продумывать систему упражнений и виды работы детей на уроке, которые отличались бы разнообразием и способствовали более эффективному усвоению детьми таблицы сложения.
Изучение случаев вычитания, соответствующих табличным
Случаям сложения
Вычитание чисел в пределах 20 начинается с раскрытия вычислительных приемов. Целесообразно показать детям три приема.
1. Прием, когда число вычитается по частям (с использованием наборного полотна с двумя рядами карманов):
12-4
/\
12-2-2
2. Прием, основанный на знании соответствующего случая сложения (т.е. состава числа). Здесь необходимо вспомнить зависимость между компонентами и результатом действия сложения:
12-4
12=4+8
Прием, где все вычисления сводятся к вычислениям в пределах 10
12-4
10-4+2
Названные приемы не обязательно рассматривать все вместе.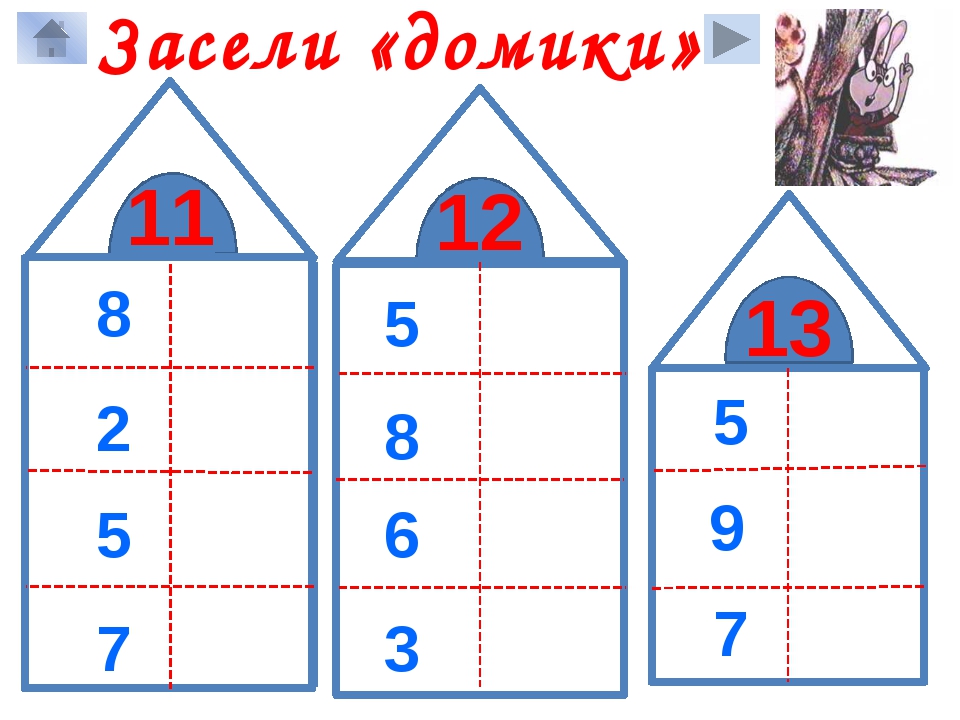 После усвоения первого из них, следует перейти к изучению случаев вычитания. При их рассмотрении целесообразно придерживаться той последовательности, которая предложена авторами учебников математики для начальных классов. На одном уроке рекомендуется рассмотреть все случаи вычитания из одного числа, начиная с 11. Здесь можно и нужно наряду с применением первого приема показать и другие. В дальнейшем учащимся следует предоставить возможность выбора приема, наиболее легкого для них. При изучении вычитания не следует прекращать работу, направленную на запоминание детьми таблицы сложения. Ее надо продолжить, обучая при этом увязывать случаи сложения с соответствующими случаями вычитания.
После усвоения первого из них, следует перейти к изучению случаев вычитания. При их рассмотрении целесообразно придерживаться той последовательности, которая предложена авторами учебников математики для начальных классов. На одном уроке рекомендуется рассмотреть все случаи вычитания из одного числа, начиная с 11. Здесь можно и нужно наряду с применением первого приема показать и другие. В дальнейшем учащимся следует предоставить возможность выбора приема, наиболее легкого для них. При изучении вычитания не следует прекращать работу, направленную на запоминание детьми таблицы сложения. Ее надо продолжить, обучая при этом увязывать случаи сложения с соответствующими случаями вычитания.
Урок математики по теме «Сложение и вычитание чисел в пределах 20». 1-й класс
| Класс | первый |
| Тип урока | Урок применения полученных знаний |
| Форма урока | Урок-путешествие |
| Тема | Сложение и вычитание чисел в пределах двадцати с переходом через разряд. |
| Цель урока | Совершенствовать навыки счёта в пределах 20 с переходом через разряд. |
| Задачи |
|
| Формы работы | Индивидуальная, групповая, коллективная, в парах |
| Оборудование | Карточки с заданиями, смайлики для рефлексии, доска магнитная, мультимедийный проектор, компьютер, презентация. |
Ход урока
1. Организационный момент.Учитель:
А ну-ка детки встали в ряд!
Готов к работе мой отряд?
Занимай свои места.
Математика сложна.
Но скажу с почтеньем я,
Математика нужна
Всем без исключения.
– Ребята, вы на какой урок пришли? (Слайд №2)
(Ответ: на урок математики).
Учитель. Этот урок особенный, мы отправляемся в сказочное путешествие. Узнать, где мы побываем, вам поможет этот ребус. Убери из таблицы цифры и арифметические знаки, и ты прочтешь слово. (Слайд №3).
Узнать, где мы побываем, вам поможет этот ребус. Убери из таблицы цифры и арифметические знаки, и ты прочтешь слово. (Слайд №3).
| с | 5 | = | 7 | 12 | а |
| + | к | 14 | 20 | к | – |
| 9 | 16 | а | з | 8 | 19 |
Дети: « Мы отправляемся в сказку»
2. Сообщение темы и цели урока.
Сообщение темы и цели урока.– Тема нашего занятия: «Сложение и вычитание чисел в пределах 20».
– Какие знания и умения пригодятся нам во время путешествия?
(В путешествии пригодятся: умения выполнять сложение и вычитание в пределах 20; сравнивать выражения; быть внимательным, сообразительным).
Учитель. Чтобы встретится с героиней сказки, нам необходимо выполнить первое задание. В класс по почте доставлена шифрограмму. Разгадав ее, вы узнаете, кто нуждается в вашей помощи.
| 9+6= | л | |
| 4+8= | о | |
| 16-8= | а | |
| 7+9= | у | |
| 9+2 | к | |
| 7+7= | ш | |
| 18-9= | З |
| 9 | 12 | 15 | 16 | 14 | 11 | 8 |
Выполнив задание, дети дают ответ: Золушка.
(На экране появляется изображение Золушки). (Слайд №4)
Учитель. Ребята, на уроке вы сегодня не просто ученики, а добрые «волшебники». Потому что только волшебники творят чудеса. И вы должны обязательно помочь Золушке выполнить все задания её мачехи.
– Посмотрите, Золушка встревожена. Она беспокоится: « Очень хочется побывать на балу у короля. Но успею ли я на королевский бал, ведь мачеха столько заданий приготовила?»
– Не волнуйся Золушка, ребята тебе помогут.
3. Устный счет.Чтобы работа была успешной, начнём работу с гимнастики мозга.
- сосчитаем до 20
- обратите внимание на ряд 12, 16, 20 что можете о них сказать? (двузначные, увеличиваются на 4)
- Назовите самое большое из них?
- Самое маленькое?
- На сколько число12 меньше числа 20?
- Какое число предшествует 20, 15, 17, 12?
- Какое число следует за 14, 16, 13, 19, 17?
После разминки можно помочь и любимой Золушке.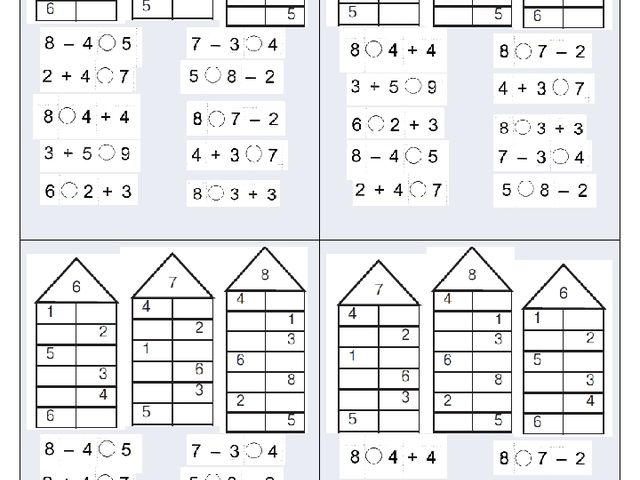
Учитель: Первое задание мачехи: прополоть грядки на ее огороде с морковью, свеклой, помидорами. А грядки мачехи – наши математические цепочки. Узнайте результат данных выражений.
Дети открывают тетради, и каждый выполняет свою цепочку.
Работа дифференцирована по трем группам.
- Рисунок морковки. 15-9+8-7+6-4+9-10= (повышенный уровень).
- Рисунок свеклы. 13-8+7-3+5-7= (средний уровень).
- Рисунок помидора.12-6+4+8-9= (низкий уровень).
Учитель. Проверка. Выслушиваются ответы детей. (Слайд №5).
4. Закрепление пройденного материала.(Слайд №6).
Учитель. Второе задание мачехи: распределить собранные фрукты в три корзины:
Примеры с ответом 12. |
Примеры с ответом 14 | Примеры с ответом 8 |
Работа в парах. Дети заполняют таблицу самостоятельно на карточках.
| 6+6= | 5+9= | 16-8= |
| 6+8= | 17-9= | 7+5= |
| 8+4= | 14-6= | 7+7= |
| 9+3= | 13-5= | 9+5= |
Проверка результатов заполнения таблицы. (Слайд №7)
(Слайд №7)
6. Закрепление пройденного материала (продолжение).Мы устали чуточку,
Отдохнём минуточку.
Поворот, наклон, прыжок,
Улыбнись, давай, дружок.
Ещё попрыгай: раз, два, три.
На соседа посмотри,
Руки вверх и тут же вниз,
И за парту вновь садись.
Учитель. Сейчас мы будем работать с выражениями. Сравнивать выражения с доказательствами и выводами, вставлять в выражения пропущенные знаки. Своей волшебной работой помогаем создать для Золушки карету, лошадей и кучера. И только тогда Золушка отправится на королевский бал.
Сравните выражения. На доске:
| 7+9 * 6+9 | 5+8 * 5+9 | 6+6 * 7+7 |
Учащиеся называют полученные ответы, при правильном их выполнении появляются карета, лошади и кучер. (Слайд №8)
(Слайд №8)
Учитель. Какие приемы вы применяли в сравнении выражений?
Предполагаемый ответ: сравнивали слагаемые в выражениях и делали вывод.
Подводим итоги: ваша волшебная работа помогла из тыквы создать карету, мышек превратить в шесть лошадей, а крысу превратить в кучера.
7. Самостоятельная работа.Учитель. Чтобы Золушка встретилась с принцем, давайте, ребята, решим еще несколько заданий.
- 1. Вспомнив состав числа 12.
Работа в группах. Каждая группа по карточки с примерами заполняют пропуски.
| 6 + 0 =12 | 7 + 0 = 12 | 5 + 0 = 12 |
| 9 + 0 = 12 | 8 + 0 = 12 | 4 + 0 = 12 |
Проверяют задание. (Слайд №9)
(Слайд №9)
- 2. Каждая группа решает задачи.
| В хоре 11 кузнечиков. Для Золушки песни распевали. Вскоре 2 кузнечика Голос потеряли. Сосчитай без лишних слов, Сколько в хоре стало голосов? |
Ежик по грибы пошёл 13 рыжиков для Золушки нашёл. 8 положил в корзину, Остальные же – на спину. Сколько рыжиков везёшь, На своих иголках, ёж? |
7 весёлых васильков. 5 ромашек-желтоглазок. Принц Золушке дарил.. Сколько же цветов в букете? |
Проверка (Слайд №10)
Учитель. Золушка встретилась со своим Принцем. (Слайд №11).
Она счастлива и благодарит своих волшебников за работу. (Слайд №12).
8. Рефлексия.Наш урок подходит к концу. Оцените свою работу. Возьмите смайлики, которые соответствуют вашему настроению после нашего урока.
– Кто доволен собой?
– У кого были трудности в работе?
Ребята, а почему вы помогали Золушке? Что сделало вас «волшебниками?» Можно ли в настоящей жизни стать «волшебниками»? Объясните. Выслушиваются ответы учащихся.
Детский сад Математика — обучение и практика по математике для детских садов
Все приложения
[«« # »,« Моя учетная запись »]От распознавания чисел до счета и операций, учащиеся развивают чувство числа и концептуальное понимание, которое им понадобится в 1 классе .
Подготовительный класс Детский садКласс 1Класс 2Класс 3Класс 4Класс 5МОДУЛЬ 1 Числа до 10
Тема A: Определение одинаковых, разных и похожих
Учащиеся рассматривают размер, форму и цвет предметов, чтобы определить, одинаковы ли они, разные , или подобное.Кроме того, они представлены разнородным объектам, которые «совпадают» из-за связанной функции, например, носку и обуви.
Определите два из трех одинаковых объектов.
На изображении трех существ выберите два одинаковых. Нажмите на те, которые точно такие же. Тот, который отличается, будет другого цвета, иметь другие особенности и другую позу.
Определите два из трех разных объектов.
Узнайте, как определить, есть ли различия.На картинке с тремя существами выберите два, которые не совпадают. Два будут точно такими же. Тот, который отличается, будет другого цвета, иметь разные функции и другую позу
Определите различия между двумя похожими изображениями
Сравните два изображения и найдите 5 различий между ними
Определите элементы, которые принадлежат друг другу, на основе их использования
Сопоставляйте предметы, которые используются вместе. Выберите элемент из трех вариантов, который используется с данным элементом.Переместите нужный элемент рядом с изображением. Затем сопоставьте три изображения с предметами, с которыми они обычно используются.
Определите, имеют ли два объекта одинаковую форму (с разными цветами и узорами).
Укажите, имеют ли два объекта одинаковую форму. Дается одна форма, затем рядом с ней прокручиваются разные формы разного цвета и узоров. Щелкните галочку или значок x, чтобы указать, совпадают ли формы.
Определите, имеют ли два объекта одинаковый цвет (с разными формами и узорами).
Научитесь сравнивать цвета двух объектов.Предметы могут быть одноцветными или двухцветными. Щелкните Проверить или X, чтобы указать, совпадают ли цвета двух фигур.
Определить, имеют ли два объекта одинаковый узор (с разными формами и цветами).
Сравните узор на двух объектах. Совместите каждую из трех фигур с одним и тем же узором серого цвета, перетащив серый узор вверх к его партнеру. Затем нажмите «Проверить» или «X», чтобы указать, совпадают ли рисунки двух фигур.
Определить, совпадают ли форма, цвет или рисунок у двух объектов.
Определить схожие характеристики двух фигур.Фигуры могут различаться по форме, рисунку и цвету.
Тема B: Сортировка и подсчет похожих объектов
Учащиеся сортируют похожие предметы по узким категориям (например, «медведи»), широким категориям (например, «одежда») и самоопределяемые категории. Они идентифицируют объекты, не принадлежащие к группе, без присвоения имени категории. Наконец, студенты считают, чтобы определить общее количество предметов в группе.
Тема C: Числа до 5 в различных конфигурациях, математические рисунки и выражения
Учащиеся считают последовательно, чтобы определить позицию, или суммируют до 5.Они связывают объекты с цифрами и видят цифры в различных шрифтах. Они работают с выровненными объектами, разбросанными объектами, пальцами и числовой линией. Учащиеся изучают стратегии подсчета при совпадении 1: 1 и узнают, что наборы предметов с одинаковым итогом можно выровнять по-разному.
Сопоставьте пронумерованный набор из 1, 2 или 3 кубиков с одинаковым пронумерованным набором кубиков.
Сопоставьте наборы кубиков. Показан человек с 1-3 кубиками. Затем есть еще трое людей с наборами кубиков.Выберите набор, соответствующий первому набору. Числа написаны разными шрифтами, чтобы практиковаться в распознавании чисел.
Сопоставьте нумерованные и ненумерованные наборы кубиков с числом 1-3
Выберите изображения, на которых показано указанное количество кубиков (1, 2 или 3). В каждой задаче дано по четыре рисунка. Цифры написаны разными шрифтами, чтобы научиться распознавать числа. Некоторые числа затемнены, чтобы облегчить подсчет кубиков.
Сопоставьте наборы кубиков с числами 1-3
Имея изображения с числами 1-3 и карточки с наборами кубиков, перетащите правильный набор кубиков к каждому числу.Цифры и наборы расположены в случайном порядке. Одновременно отображаются три набора кубиков и чисел.
Сопоставьте числа 1-3 с их позициями в числовой строке, помеченной числами и точечными узорами.
Поместите числа в числовую строку. Перетащите числа 1-3 в соответствующее место на числовой строке. Числовая линия охватывает числа от 0 до 10 и помечена числами и точечными рисунками каждого числа. Номера представлены различными шрифтами.
Определите пронумерованный набор кубиков, который соответствует идентичному набору из 4 или 5 пронумерованных кубов.
Сопоставьте наборы кубиков.Показан один человек с 4 или 5 кубиками. Затем есть еще три человека с наборами кубиков от 2 до 5. Выберите набор, соответствующий первому набору. Числа написаны разными шрифтами, чтобы практиковаться в распознавании чисел.
Сопоставьте нумерованные и ненумерованные наборы кубиков с номером 4 или 5
Выберите изображения, на которых показано указанное количество (4 или 5) кубиков. В каждой задаче дано по четыре рисунка. Цифры написаны разными шрифтами, чтобы научиться распознавать числа.Некоторые числа затемнены, чтобы стимулировать подсчет кубиков
Подсчитайте 2-5 выровненных объектов, чтобы определить общее количество
Для ряда из 2-5 кубиков введите или выберите количество показанных кубиков
Сопоставьте числа 1-5 с расположением пальцы, отображаемые на руке
Сопоставьте руку с количеством отображаемых пальцев и найдите правильную цифру. Одновременно отображаются три числа и руки.
Сопоставьте наборы кубиков с числами 1-5
Сопоставьте набор объектов с номером.Учитывая изображения с числами 1-5 и карточки с наборами кубиков, перетащите правильный набор кубиков на каждое число. Цифры и наборы расположены в случайном порядке. Одновременно отображаются три набора кубиков и чисел.
Определите пронумерованные наборы кубиков, которые соответствуют заданной сумме до 5
. Выберите изображения, которые показывают указанное количество (1-5) кубиков. В каждой задаче даны по четыре картинки с наборами кубиков. Более чем одно изображение может содержать желаемое количество кубиков, поэтому выберите все, которые применяют
Сопоставьте числа 1–5 к их позициям на числовой строке, помеченной числами и точечными узорами
Перетащите числа 1–5 в соответствующее место на числовая строка.Числовая линия охватывает числа от 0 до 10 и помечена числами и точечными рисунками каждого числа. Цифры для перетаскивания представлены различными шрифтами.
Выровняйте разбросанные объекты для подсчета и определения общего числа
Представлен случайный набор объектов (1-5). Перетащите объекты в коробку, чтобы выровнять их, чтобы было легче считать. Затем введите или выберите показанное число
Подсчет, чтобы найти общее количество разбросанных объектов, сопоставив числа 1: 1 с объектами
Подсчитайте количество показанных объектов.Нажмите на них, чтобы изменить их цвет, чтобы отметить, что они были подсчитаны. Затем выберите число
Определите набор разбросанных объектов, который соответствует заданному общему количеству до 5
Подсчитайте количество показанных объектов. Введите или выберите номер. Затем, учитывая количество объектов, которые нужно найти, выберите контейнер с правильным количеством элементов в нем
Тема D: Концепция нуля и работа с числами 0-5
Учащиеся используют цифры 1-5 для последовательности объектов и определить позицию или сумму.Они работают с выровненными объектами, разбросанными объектами и числовой линией. Они исследуют состав числа 3 и начинают использовать знаки +, — и =.
Тема E: Работа с числами 6–8 в различных конфигурациях
Учащиеся считают последовательно, чтобы определить позицию или общее количество до 9. Они связывают объекты с цифрами и видят цифры в различных шрифтах. Они работают с выровненными объектами, разбросанными объектами, пальцами и числовой линией. Учащиеся изучают стратегии подсчета при совпадении 1: 1 и узнают, что наборы предметов с одинаковым итогом можно выровнять по-разному.
Учтите, что на единицу больше 5 — это 6
Выучите число 6. Показаны пять элементов. Выберите правильную цифру, чтобы представить это. Затем добавляется еще один элемент, показывая, что цифра 6 идет после 5
Распознайте, что одно больше 6 — это 7
Выучите число 7. Показаны шесть элементов. Выберите правильную цифру, чтобы представить это. Затем добавляется еще один элемент, показывающий, что цифра 7 стоит после 6
Определить количество пальцев до 7, отображаемых на двух руках
Отображаются две руки с отображением от 0 до 7 пальцев.Щелкните или введите количество показанных пальцев
Сопоставьте числа от 0 до 7 с расположением пальцев, отображаемым на двух руках
Показаны три числа и три изображения рук с пальцами. Сопоставьте руки с числами 0-7
Определите набор выровненных объектов, который соответствует заданной сумме до 7
Определите изображение, показывающее до 7 кубиков. Даны три варианта ряда кубиков. Нажмите на тот, который соответствует номеру
Сопоставьте пронумерованные образцы точек с одинаковыми пронумерованными образцами точек до 7
Сопоставьте пронумерованные образцы точек.Маленькие осьминоги держат флаги с числами и точечными узорами. Перетащите к нему все, что соответствует номеру и рисунку отца осьминога. Цифры написаны различными шрифтами.
Определите наборы пронумерованных кубов, которые соответствуют заданному общему количеству 6 или 7 (Часть 1)
Определите количество показанных кубиков из двух возможных вариантов. Затем добавляется куб. Снова определите число, чтобы получилось 6 или 7. Выберите картинки с таким же общим количеством кубиков. Цифры на рисунках написаны различными шрифтами.
Определите наборы пронумерованных кубиков, которые соответствуют заданной сумме 6 или 7 (Часть 2)
Выберите наборы, содержащие указанную сумму.Выделите все картинки, содержащие заданное количество кубиков (6 или 7). Цифры на рисунках написаны различными шрифтами, а кубики расположены в различных конфигурациях
Сопоставьте номера 1-7 с их позициями на числовой строке, помеченной числами и точечными узорами
Для данной числовой линии перетащите два числа от 1 до 7 до их правильного местоположения на линии. Числовая строка помечена числами от 0 до 10 и точечными узорами, представляющими числа. Цифры, которые нужно перетаскивать, написаны различными шрифтами.
Подсчитайте объекты по мере их удаления от набора и определите общее количество.
Птицы улетают от дерева по одной.Подсчитайте количество улетевших птиц и введите или выберите количество улетевших
Подсчитайте разбросанные объекты двумя разными способами, чтобы получить одинаковую сумму.
Узнайте, что порядок, в котором подсчитывается группа, не влияет на общую сумму. Следуя стрелкам, нажимайте на бусинки, чтобы изменить их цвет, и сосчитайте их. Затем введите номер. Подсчитайте элементы в другом порядке и введите число еще раз.
Обратите внимание на то, что одно число больше 7 равно 8
. Выучите число 8. Показаны семь элементов.Выберите правильную цифру, чтобы представить это. Затем добавляется еще один элемент, показывающий, что цифра 8 идет после 7
Определить количество пальцев до 8, отображаемых на двух руках
Отображаются две руки с отображением от 0 до 8 пальцев. Щелкните или введите количество показанных пальцев
Сопоставьте числа от 0 до 8 с расположением пальцев, отображаемым на двух руках
Показаны три числа и три изображения рук с пальцами. Сопоставьте руки с числами 0-8
Определите набор выровненных объектов, который соответствует заданной сумме до 8
Определите изображение, показывающее до 8 кубиков.Даны три варианта ряда кубиков. Нажмите на тот, который совпадает с номером
Определите наборы пронумерованных кубиков, которые соответствуют заданной сумме до 9 (Часть 1)
Определите количество показанных кубов из двух вариантов. Затем добавляется по одному кубу. Снова определите число, всего 8. Затем будут показаны три или четыре изображения. Выберите те, у которых одинаковое количество кубиков. Цифры на рисунке
Определите наборы пронумерованных кубиков, которые соответствуют заданной сумме до 9 (Часть 2)
Выберите все изображения, содержащие заданное количество кубиков, до 9.Цифры на рисунках написаны различными шрифтами, а кубы расположены в различных конфигурациях.
Подсчитайте объекты, когда они удаляются от набора, и определите общее количество.
Птицы улетают от дерева по одной. Подсчитайте количество улетевших птиц и введите или выберите общее количество улетевших
Подсчитайте разбросанные объекты двумя разными способами, чтобы получить одно и то же общее количество
Узнайте, что порядок, в котором подсчитывается группа, не влияет на общую сумму. Следуя стрелкам, нажимайте на бусинки, чтобы изменить их цвет, и сосчитайте их.Затем введите номер. Подсчитайте их еще раз в другом порядке и введите число еще раз.
Определите общее количество разбросанных объектов после того, как известное общее количество было преобразовано.
Узнайте, что количество объектов не меняется при их перемещении. Подсчитайте количество предметов в контейнере и введите сумму. Затем предметы перемещаются. Подсчитайте их еще раз и введите общее
Тема F: Работа с числами 9–10 в различных конфигурациях
Учащиеся все больше работают с 0 и числами 6–10, чтобы определить итоги и распознать цифры.Они работают с разрозненными объектами, неидентичными объектами, пальцами и числовой линией.
Тема G: Еще одно с числами 0–10
Учащиеся используют знакомые представления (объекты, блоки с основанием 10, числовая прямая и уравнения), чтобы изучить концепцию еще одного. Они начинают рассчитывать, а не считать все.
Тема H: На один меньше с числами 0–10
Учащиеся используют знакомые представления (объекты, блоки с основанием 10, числовая линия и уравнения), чтобы изучить концепцию «на единицу меньше».Для решения задач они чередуются между +1 и -1.
Определите недостающие числа в числовой строке с номером 9-1
Научитесь считать в обратном порядке. Введите два пропущенных числа в строке с обратным номером.
Определите на один больше и на один меньше в числовой строке.
Укажите число на один больше и на единицу меньше заданного числа в числовой строке. Числовая строка помечена цифрами 0 и 10 и начальным значением. Введите число слева и справа от начального значения.
Продолжить последовательность счета в числовой строке от заданной точки, отсчитывая назад
Обратный отсчет в пределах от 10 до 0.Учитывая числовую строку, отсчитайте в обратном порядке от 10 до 0 один раз. Затем потренируйтесь считать два числа слева от заданного числа, вводя второе число
Решите -1 уравнения на основе модели блоков с основанием 10
Выучите вычитание 1. Отображается строка кубиков и последний один выскакивает при нажатии. Введите количество оставшихся кубиков и ответ на уравнение вычитания
Решите уравнения -1 на основе модели числовой линии
Используйте числовую линию, чтобы найти ответ на уравнение -1.На числовой строке обозначены только числа 0, 5 и 10. Сверчок прыгает на одну цифру влево. Введите число, на которое он приземляется, затем введите это же число в качестве ответа на уравнение
Решите -1 уравнения (Уровень 1)
Практикуйте вычитание 1. Дана либо модель с кубиками, либо числовая линия, чтобы помочь найти ответ
Решите -1 задачу на основе известной суммы и сценария с использованием объектов.
Подсчитайте количество предметов в банке и введите число. Затем отображается вторая банка, в которой элементы переставляются, и один объект удаляется.В некоторых задачах первая сумма написана на банке, скрывая предметы, и один предмет удаляется. Введите новую сумму
Решите -1 проблемы на основе известной суммы и сценария с использованием объектов
Укажите общую сумму после удаления одного объекта из известного числа. Показана банка с указанием ее количества. Один элемент удален. Введите новую сумму
Решите -1 уравнение (Уровень 2)
Практикуйте вычитание 1. Отображается только уравнение вычитания. Введите сумму
Решите уравнения +1 и -1 на основе модели блоков с основанием 10
Попрактикуйтесь в сочетании задач, используя сложение и вычитание 1.Блоки используются для моделирования решаемого уравнения. Введите количество блоков и ответ на уравнение
Решите уравнения +1 и -1 на основе модели числовой линии
Попрактикуйтесь в сочетании задач, используя сложение и вычитание 1. Числовая линия от 0 до 10 используется для смоделировать решаемое уравнение. Введите количество блоков и ответ на уравнение
Решите задачи +1 и -1 на основе известной суммы и сценария с использованием объектов
Попрактикуйтесь в сочетании задач, используя сложение и вычитание 1.Банку с предметами используют для моделирования решаемого уравнения. Введите количество блоков и ответ на уравнение
Решите уравнения +1 и -1
Решите уравнения, включающие сложение или вычитание 1. Модель не используется. Отображается только уравнение. Введите ответ
МОДУЛЬ 2 Двумерные и трехмерные формы
Тема A: Двумерные плоские формы
Учащиеся знакомятся с внешним видом и названиями двумерных фигур. Они работают с квадратами, кругами, треугольниками, прямоугольниками и шестиугольниками.Учащиеся выделяют примеры из числа не являющихся примерами. Наша звуковая функция позволяет не читателям получать доступ к именам фигур.
Определение положения над и под объектом
Научитесь определять положение как над или под объектом. Выберите указанное местоположение, щелкнув правильное положение.
Позиционировать объекты выше и ниже объекта.
Позиционировать 1 или 2 объекта выше или ниже неподвижного объекта. Выберите правильное местоположение и правильный цвет в соответствии с указаниями.
Поместите объект перед или за объектом (Часть 1)
Научитесь определять положение как перед объектом или позади него.Выберите указанное местоположение, щелкнув правильное положение.
Поместите объект перед или за объектом (Часть 2)
Определите местоположение как «перед» или «за» объектом. Перетащите символ в указанную позицию
Определите линии
Научитесь определять линии. Выберите линии с различной ориентацией из группы, содержащей линии и кривые
Определите изогнутые линии
Научитесь определять кривые. Выберите кривые в различных ориентациях и формах из группы, содержащей линии и кривые.
Определите прямоугольники, треугольники или круги из набора фигур.
Определите прямоугольники, треугольники и круги из наборов фигур.Нажмите на фигуры, которые соответствуют указанному рисунку.
Определите свойства треугольников
Узнайте характеристики треугольника. Затем выберите те формы, которые являются треугольниками.
Определите свойства прямоугольников.
Изучите характеристики прямоугольника. Затем выберите те формы, которые являются прямоугольниками.
Определите шестиугольники из набора фигур.
Определите, какая из двух форм является шестиугольником. Затем выберите все шестиугольники из множества форм.
Определите свойства шестиугольников.
Изучите характеристики шестиугольника.Затем выберите те формы, которые являются шестиугольниками, чтобы завершить изображение.
Определите отдельные 2D-формы на большом изображении.
Используйте разные цвета для обозначения фигур. Следуйте инструкциям, чтобы раскрасить каждый тип формы в указанный цвет.
Определите объекты реального мира, состоящие из двухмерных фигур.
Выберите объекты реального мира, содержащие заданную форму, из набора из 6 объектов. Фигуры включают в себя круги, прямоугольники и треугольники.
Сопоставьте объекты реального мира с их двумерными формами.
Выберите правильную двумерную фигуру, моделирующую объекты реального мира.Объект и соответствующая ему фигура могут быть ориентированы в разных направлениях.
Сопоставьте двухмерную форму с ее именем.
С учетом имени формы сопоставьте ее с правильной формой. Щелкните значок x или галочку рядом с каждой из трех фигур, чтобы указать, соответствует ли она слову
Определить фигуры с заданным количеством сторон, углов или изогнутых линий
Выберите фигуры с указанными характеристиками. Из набора из 5 фигур выберите те, которые имеют заданное количество сторон или углов.
Определите двухмерные формы и переместите их в положения выше, ниже, спереди и сзади.
Практикуйте двухмерные формы и положение.Перетащите указанную фигуру в указанное место относительно статического изображения
Тема B: Трехмерные твердые формы
Учащиеся знакомятся с внешним видом и названиями трехмерных фигур. Они работают с кубами, сферами, конусами и цилиндрами. Студенты также укрепляют свое понимание позиций (вверху, внизу, впереди, рядом и позади). Наша звуковая функция позволяет не читателям получать доступ к именам фигур.
Сортировка фигур по категориям плоские и твердые
Научитесь различать двухмерные и трехмерные фигуры.Затем отсортируйте их по категориям
Сопоставьте формы с реальными объектами
Сопоставьте реальные объекты с трехмерными сплошными фигурами
Определите цилиндры среди трехмерных фигур
Научитесь определять цилиндры из набора из трех -мерные тела. Щелкните цилиндры среди группы других твердых тел
Определите конусы среди трехмерных фигур
Практикуйтесь в определении конусов из набора трехмерных тел. Щелкните конусы среди группы других твердых тел
Определите кубы среди трехмерных фигур
Научитесь определять кубы из набора трехмерных тел.Щелкните кубы среди группы других твердых тел
Определите сферы среди трехмерных фигур
Практикуйтесь в определении сфер из набора трехмерных тел. Щелкните сферы среди группы других твердых тел
Определите трехмерные формы по имени
Выберите правильное имя для данного твердого тела. Будет предложено два варианта выбора.
Определить трехмерные формы среди реальных объектов.
Найдите основные трехмерные твердые формы, используемые в изображениях реальных объектов. Если представлен набор из менее чем 10 изображений, щелкните те, которые содержат заданные твердые тела.
Положение 3-D фигуры выше, ниже, перед и позади объекта.
Практикуйте трехмерные тела и положение.Перетащите указанное твердое тело в указанное место относительно статического изображения
Идентифицируйте объекты в положениях выше, ниже, рядом и перед ним
Попрактикуйтесь в словах положения «выше», «ниже», «рядом с», и «перед». Щелкните один из двух символов, которые находятся в правильном положении относительно указанного символа.
Тема C: Двумерные и трехмерные формы
Учащиеся применяют свое предыдущее понимание плоских и твердых форм, чтобы различать их.
МОДУЛЬ 3 Сравнение длины, веса, вместимости и чисел с 10
Тема A: Сравнение длины и роста
Используя знакомые реальные предметы, учащиеся используют язык сравнения (самый высокий, высокий, самый длинный, длинный, самый короткий , короче), поскольку они исследуют длину.
Тема B: Сравнение длины и высоты соединяющих палочек куба в пределах 10
Учащиеся работают со знакомыми соединяющими кубиками для подсчета кубиков и определения длины палочек. Они сравнивают длину, используя более длинные, короткие и такие же, как.
Тема C: Сравнение веса
Используя знакомые, реальные предметы, учащиеся используют язык сравнения (тяжелее, легче, примерно такой же), когда они исследуют вес. Для этого они используют виртуальную шкалу баланса для сравнения.
Тема D: Сравнение томов
Учащиеся выучивают слово «вместимость» и используют его для измерения количества жидкости, которое может вместить контейнер. Используя единицу «стаканы», они измеряют, переливая стаканы в большую емкость и переливая из большей емкости в стаканы.Они также сравнивают измеренную вместимость двух контейнеров.
Тема E: Достаточно?
Чтобы подготовиться к сравнению чисел, учащиеся работают с объектами реального мира, чтобы определить, «достаточно» ли их, чтобы составить пары. Затем они определяют, имеет ли набор реальных объектов больше, меньше или столько же, что и другой набор объектов.
Тема F: Сравнение наборов в пределах 10
Учащиеся сравнивают два набора объектов, используя слова больше, меньше и тот же. Они работают с похожими объектами, как выровненными, так и разбросанными.
Тема G: Сравнение цифр
Учащиеся сравнивают два набора предметов, используя слова больше, меньше и тот же. Они работают с похожими и разными объектами, как выровненными, так и разбросанными, с вертикальным и горизонтальным выравниванием. Наконец, они начинают сравнивать числа.
МОДУЛЬ 4 Числовые пары, сложение и вычитание до 10
Тема A: Композиции и разложения 2, 3, 4 и 5
Учащиеся используют знакомые блоки с основанием 10, сценарии с объектами, числовую линию и уравнения для изучить основные концепции сложения.Студенты определяют как суммы, так и недостающие слагаемые, постепенно переходя от конкретных моделей к абстрактным уравнениям.
Тема B: Разложение 6, 7 и 8 на числовые пары
Учащиеся расширяют свое понимание числовых связей до сумм 6, 7 и 8. Они работают с наборами похожих объектов двух цветов для подсчета частей (на цвет) и общ. Затем они используют эти числа для построения числовых связей на основе изображения объектов.
Создание числовой связи на основе набора объектов; Определите добавление в числовой связи
Создайте числовую связь, используя изображение объектов двух разных цветов.Определите сумму и слагаемые, затем перетащите их в нужное место в облигации. Кроме того, заполните недостающее дополнение в устном заявлении облигации
Определите недостающий номер в цифровой связи на основе набора объектов
Заполните недостающее значение на числовой связи. Затем также заполните устное заявление о залоге. Предоставляется изображение объектов в двух цветах.
Построение числовой связи на основе набора объектов; Определите общую сумму в числовой связи (всего 6)
Создайте числовую связь из объектов двух цветов, найденных на монохроматическом изображении.Заполните все числа в номере-связке. Кроме того, завершите словесное заявление о взаимосвязи между числами
Раскрасьте набор объектов в два цвета, чтобы представить заданное число bond
Раскрасьте набор кругов в два разных цвета, чтобы соответствовать заданному номеру bond
Построить число облигация на основе набора предметов; Определите общее количество облигаций (всего 7)
Практическое число облигаций для набора из 7 яиц. Для начала выберите цвет и рисунок яиц.Затем запишите номер для показанного набора яиц. Кроме того, завершите словесное заявление о взаимосвязи между числами
Создайте связь чисел на основе набора объектов; Определите общую сумму в числовой связи (всего 8)
Запишите числовые связи для набора из 8 объектов в двух цветах. Кроме того, заполните словесную формулировку взаимосвязи между числами.
Запишите числовую связь на основе модели, которую создают учащиеся (всего 7 или 8).
Напишите числовую связь для набора из 7 или 8 предметов.Раскрасьте кружки, используя два цвета, затем заполните числовую связь, соответствующую цветовой конфигурации. Кроме того, заполните пробел в выписке о числовом связывании
Тема C: Сложение с итогами 6, 7 и 8
Учащиеся углубляют свое понимание сложения, укрепляя связь между конкретными объектами, блоками с основанием 10 и уравнениями. Они определяют как суммы, так и недостающие слагаемые. Они записывают уравнения, включая символ +.
Определите недостающее слагаемое и суммируйте уравнения сложения до 5 на основе сценария с объектами
Заполните уравнение сложения.Анимация с набором стационарных объектов и набором добавляемых объектов дается всего до 5. Заполните пробелы в уравнении сложения
Используйте кубы для представления сценария добавления на основе объектов и сопоставления шаблонов кубов к добавлению сценариев
Практикуйте индивидуальную переписку. Смоделируйте сценарий сложения с кубиками двух разных цветов
Решите уравнения сложения для 5 на основе модели блоков с основанием 10
Смоделируйте задачу сложения с кубами.Во-первых, изобразите недостающее дополнение кубиками. Введите новую сумму после каждого добавленного куба, затем введите решение уравнения. В других задачах используется модель куба, поэтому просто введите сумму
Определите недостающие слагаемые в уравнениях с суммой 4
Напишите уравнение для данной модели куба. Даны кубики двух цветов. Напишите уравнение сложения, представляющее модель. Подсказки даются для каждого бланка.
Решите +/- уравнения в пределах 5 на основе перемещений в числовой строке.
Представьте задачу сложения слов в числовой строке.Начиная с заданного значения числовой строки, переместите дополнительную сумму, чтобы получить итоговое значение до 5. Соответствующее уравнение сложения отображается после правильного перемещения по числовой строке
Определите недостающее слагаемое в уравнениях с суммой 4
Найти недостающий член в уравнении сложения с суммой 4. Сначала предоставляется модель куба. Затем заполните пропуски без модели.
Определите недостающее слагаемое в уравнениях с суммой 5
Найдите недостающее слагаемое в уравнении сложения с суммой 5.Во-первых, предоставляется модель куба. Затем заполните пробелы без модели.
Запишите сценарии сложения с объектами как уравнения для 5
Напишите уравнение сложения для анимации с суммой 5. Отображается ряд неподвижных объектов. Затем появляются дополнительные объекты. Напишите уравнение с подсказками для каждого бланка.
Используйте блоки base-10 для представления сценариев сложения с объектами к 5
Смоделируйте задачу сложения с кубиками и напишите уравнение. Изобразите ситуацию сложения с кубиками.Затем заполните уравнение сложения без каких-либо подсказок для пропусков
Напишите уравнение для сопоставления числовой связи (всего 3, 4 или 5)
Напишите уравнение, представленное числовой связью с суммой 3, 4, или 5. Введите числа из связи в уравнение
Заполните уравнения сложения с суммой 6 на основе модели блоков с основанием 10
Заполните все уравнения, которые имеют сумму 6. Создайте модель куба a каждой пары слагаемых. Заполните пропуски в уравнениях, которые соответствуют моделям.
Сценарии сложения модели до 6 на числовой прямой и решите соответствующее уравнение.
Представьте задачу сложения слов на числовой строке.Начиная с заданного значения числовой строки, переместите дополнительную сумму, чтобы получить общее количество до 6. Кроме того, заполните уравнение сложения, которое представлено числовой строкой
Напишите уравнение для сопоставления числовой связи (всего 6)
Напишите уравнение, представленное числовой связью с суммой 6. Введите числа из связи в уравнение
Представьте сценарии сложения с объектами, используя блоки с основанием 10, и запишите как уравнение с суммой 6
Запишите Уравнение сложения для картинки с суммой 6.Сначала создайте кубическую модель ситуации. Затем запишите уравнение, представленное кубами.
Определите недостающее слагаемое в уравнениях с суммой 6
Найдите недостающее слагаемое в уравнении сложения с суммой 6. Сначала предоставляется модель куба. Затем заполните пробелы без модели.
Разбейте набор из 7 объектов на две группы и определите как слагаемые в уравнении.
Заполните все уравнения, сумма которых равна 7. Создайте модель куба для каждой пары слагаемых. .Заполните пропуски в уравнениях, соответствующих моделям.
Сценарии сложения модели до 7 на числовой прямой и решите соответствующее уравнение.
Представьте задачу сложения слов на числовой строке. Начиная с заданного значения числовой строки, переместите дополнительную сумму, чтобы получить общее количество до 7. Также завершите уравнение сложения, которое представлено числовой строкой
Напишите уравнение для сопоставления числовой связи (всего 7)
Напишите уравнение, представленное числовой связью с суммой 7.Введите числа из облигации в уравнение
Представьте сценарии сложения с объектами, используя блоки с основанием 10, и запишите в виде уравнения с суммой 7
Напишите уравнение сложения для изображения с суммой 7. Сначала создайте кубическая модель анимации. Затем запишите уравнение, представленное кубами.
Определите недостающее слагаемое в уравнениях с суммой 7
Найдите недостающее слагаемое в уравнении сложения с суммой 7. Сначала предоставляется модель куба.Затем заполните пропуски без модели
Завершите уравнения сложения с суммой 8 на основе модели блоков с основанием 10
Заполните все уравнения, которые имеют сумму 8. Создайте модель куба для каждой пары добавляет. Заполните пропуски в уравнениях, соответствующих моделям.
Сценарии сложения модели до 8 на числовой прямой и решите соответствующее уравнение.
Представьте задачу сложения слов на числовой строке. Начиная с заданного значения числовой строки, переместите дополнительную сумму до 8.Кроме того, заполните уравнение сложения, которое представлено числовой линией
Напишите уравнение для сопоставления числовой связи (всего 8)
Напишите уравнение, представленное числовой связью с суммой 8. Введите числа из связи в уравнение
Представьте сценарии сложения с объектами с использованием блоков base-10 и запишите как уравнение с суммой 8
Напишите уравнение сложения для изображения с суммой 8. Сначала создайте модель куба анимации.Затем запишите уравнение, представленное кубами.
Определите недостающее слагаемое в уравнениях с суммой 8
Найдите недостающий член в уравнении сложения с суммой 8. Сначала предоставляется модель куба. Затем заполните пропуски без модели.
Тема D: Вычитание из чисел до 8
Учащиеся используют знакомые блоки с основанием 10, сценарии с объектами, числовую линию и уравнения, чтобы изучить основные концепции вычитания. Учащиеся определяют как различия, так и недостающие вычитания, постепенно переходя от конкретных моделей к абстрактным уравнениям.Они также связывают вычитание и сложение посредством исследования семейств фактов.
Тема E: Разложение 9 и 10 на числовые пары
Учащиеся используют знакомые сценарии с объектами, блоками с основанием 10 и уравнениями, чтобы расширить свое понимание сложения и вычитания, включив в него 9 и 10. Учащиеся изучают состав этих итогов. чтобы повысить их осведомленность о числовых связях.
Запишите числовую связь на основе сценария с 9 реальными объектами (начните с общего числа)
Узнайте, как записать числовую связь.Определите общее количество объектов и перенесите эту информацию в графическое изображение с номерами. Затем определите, сколько показано каждого из двух типов, и введите его в графическое изображение с числовой связью. Кроме того, заполните выписку
Запишите числовую связь на основе сценария с 9 реальными объектами (невыровненными).
Завершите числовую связь для набора из 9 разрозненных объектов. Заполните недостающие числа в числовой связке на основе данной картинки. Предлагаются подсказки, помогающие ввести правильные числа.Кроме того, заполните выписку о числовой связи
Запишите числовую связь на основе сценария с 9 реальными объектами (общее количество отсутствует)
Запишите числовую связь для набора из 9 объектов. Заполните недостающие числа в числовой связке на основе данной картинки. Помощь в заполнении пропусков не оказывается. Кроме того, заполните выписку по номеру связи
Запишите числовую связь на основе сценария с 9 реальными объектами
Завершите числовую связь для изображения 9 объектов в строках. Заполните все значения в нужных местах без дополнительной помощи.Кроме того, заполните пробел в выписке по номеру связи.
Запишите цифровую связь на основе сценария с 9 реальными объектами (невыровненными).
Завершите числовую связь для изображения 9 разбросанных объектов. Заполните все значения в нужных местах без дополнительной помощи. Кроме того, заполните пробел в выписке о числовой связи.
Запишите числовую связь на основе сценария с 10 реальными объектами (начиная с общего количества)
Узнайте, как записывать числовые облигации. Определите общее количество объектов и перенесите эту информацию в графическое изображение с номерами.Затем определите, сколько показано каждого из двух типов, и введите его на графике с числовыми связями. Также заполните выписку
Запишите числовую связь на основе сценария с 10 реальными объектами (отсутствующий элемент)
Завершите числовую связь для набора из 10 разрозненных объектов. Заполните недостающие числа в числовой связке на основе данной картинки. Предлагаются подсказки, помогающие ввести правильные числа. Кроме того, заполните выписку о числовой привязке
Запишите числовую связь на основе сценария с 10 реальными объектами (без добавления)
Заполните числовую связь для изображения 10 объектов в строках.Заполните все значения в нужных местах без дополнительной помощи. Кроме того, введите недостающий номер в выписке о числовом связывании.
Запишите числовое связывание на основе сценария с 10 реальными объектами.
Запишите числовое связывание для изображения 10 объектов в строках. Заполните все значения в нужных местах без дополнительной помощи. Кроме того, введите недостающий номер в выписку по номеру связи.
Запишите числовую связь на основе сценария с 10 реальными объектами (невыровненными).
Завершите числовую связь для набора из 10 разрозненных объектов.Заполните недостающие числа в числовой связке на основе заданного изображения без дополнительных подсказок. Кроме того, заполните формуляр счисления
Тема F: Сложение с итогами 9 и 10
Учащиеся применяют свое понимание состава 9 и 10 для решения задач с пропущенными слагаемыми.
Тема G: Вычитание из 9 и 10
Учащиеся работают как с сложением, так и с вычитанием, чтобы укрепить понимание основных понятий. Они представляют сценарии с объектами, использующими блоки с основанием 10, и блоки с основанием 10 в виде уравнений.
Тема H: Образцы со сложением 0 и 1 и получением 10
Учащиеся продолжают выполнять знакомые задания, теперь используя 0 как добавление, вычитание или разность. Они используют сценарии с объектами, числовой линией и уравнениями.
МОДУЛЬ 5 Числа от 10 до 20 и счет до 100
Тема A: Подсчитайте 10 единиц и некоторые единицы
Учащиеся отличают «десятку» от единиц, используя десятикратную рамку или стержень. Они начинают изучать состав 2-значных чисел и числовых имен.Эти упражнения формируют основу для дальнейшего изучения методик позиционной ценности и сложения. Студенты впервые используют вертикальную числовую черту.
Тема B: Разложите числа 11–20 и сосчитайте, чтобы ответить «Сколько?»
Учащиеся углубляют свое понимание чисел подростков, применяя их к позициям числовой прямой и простым уравнениям.
Тема C: Представление и применение композиций и разложения чисел для подростков
Учащиеся сортируют, считают, складывают и вычитают десять и единицы.Они видят группу из десяти, отображаемую справа от единиц, а также слева, и видят как вертикальное, так и горизонтальное выравнивание 10 объектов. Они решают для разных позиций в +/- уравнениях, в которых все единицы или целые десять складываются или вычитаются.
Математическая композиция — Заключение по числовым исследованиям
Когда П диктует свою композицию, я замечаю и удивляюсь. Что меня интересует, так это мышление П. Мы говорили о добавлении 10 к 11 для увеличения числа. Он легко преобразовал все, что знал о сложении, в вычитание.А потом, как вы заметите, он продолжает использовать эту информацию для получения дополнительных математических расчетов.
П. довольно последовательна в своих композициях. Он с чего-то начинает и работает планомерно. Это улучшение по сравнению с нашей работой в главе 3, где он излагал идеи и факты без видимого порядка.
Здесь он начинает с 21 минус 10, затем прибавляет 10 к 21 и вычитает 20, затем прибавляет 10 и вычитает 30, затем добавляет еще 10 и вычитает 40. Эти преобразования происходят из нашей работы с общими различиями и лестницами.
Он знает, что 40 — это то же самое, что 4 x 10, так что преобразование было легким. Он разделил 10 пополам, а затем удвоил 4 до 8. Я нахожу это интересным, поскольку мы мало работали с делением пополам и удвоением, но поскольку он уже делает это с десятками, я знаю, что он готов. Я вытащу эту композицию, и мы сможем поговорить о его мышлении, а затем расширить идею до более сложных делений пополам и удвоений.
Мы только что говорили о том, что пятак в квадрате = четверть. И два пятака в квадрате = пятьдесят центов.Я также заметил, что он использовал свои знания умножения и применил их к квадратным корням.
Он продолжает этот путь, используя свои знания операций для создания дополнительных математических вычислений, пока он не решит пойти в совершенно другом направлении с 3 x 3 + 2.
Затем он применил свой любимый математический трюк — выполнить операцию, а затем обратное. Он умножил на 2, а затем разделил на 2. Такие вещи заставляют его смеяться.
Из этого я знаю, что P очень удобно манипулировать десятками, и у него есть хорошее представление о том, как они работают со всеми операциями.У него довольно хорошее представление о квадратах, но не о кубах, хотя мы немного с ними поигрались. Возможно, он просто не подумал использовать их в этой композиции. Он начинает использовать дробные показатели и квадратные корни, но это ново. Я подозреваю, что ему комфортно только с 5 и 10. Скоро мы захотим еще поиграть с языком.
О взаимосвязи между составом воды в облаках и числовой концентрацией капель в облаке
Абдул-Раззак, Х. и Ган, С.J .: Параметризация активации аэрозоля. 2. Множественные типы аэрозолей, J. Geophys. Res., 105, 6837–6844, https://doi.org/10.1029/1999JD
1, 2000.
Акерман, А.С., Киркпатрик, М.П., Стивенс, Д.Э., и Тун, О.Б .: The влияние влажности над слоистыми облаками на климат непрямого аэрозоля принуждение, Природа, 432, 1014–1017, https://doi.org/10.1038/nature03174, 2004.
Адамс, П. Дж. И Сайнфелд, Дж. Х .: Непропорциональное воздействие твердых частиц. выбросы от концентраций ядер конденсации глобального облака, Geophys.Res. Lett., 30, 1239, https://doi.org/10.1029/2002gl016303, 2003.
Альбрехт, Б.А .: Аэрозоли, микрофизика облаков и фракционная облачность, Наука, 245, 1227–1230, https://doi.org/10.1126/science.245.4923.1227, 1989.
Андреэ, М. О. и Розенфельд, Д .: Взаимодействие аэрозольных облаков и осадков. Часть 1. Природа и источники облачно-активных аэрозолей // Науки о Земле. Rev., 89, 13–41, https://doi.org/10.1016/j.earscirev.2008.03.001, 2008.
АпСимон, Х. М., Круз, М., и Белл, Дж. Н. Б .: Выбросы аммиака и их роль в кислотном осаждении, Атмос. Environ., 21, 1939–1946, https://doi.org/10.1016/0004-6981(87)
-5, 1987.
Бауэр, С. Э., Цигаридис, К., и Миллер, Р .: Значительный атмосферный аэрозоль загрязнение, вызванное выращиванием продуктов питания в мире, Geophys. Res. Lett., 43, 5394–5400, https://doi.org/10.1002/2016GL068354, 2016.
Баумгарднер, Д., Йонссон, Х., Доусон, В., О’Коннор, Д., и Ньютон, Р .: The спектрометр облаков, аэрозолей и осадков: новый прибор для изучения облаков исследования, Атмос.Res., 59, 251–264, https://doi.org/10.1016/S0169-8095(01)00119-3, 2001.
Беренфельд, М. Дж., Мур, Р. Х., Хостетлер, К. А., Графф, Дж., Гауб, П., Рассел, Л. М., Чен, Г., Дони, С. К., Джованнони, С., Лю, Х., Проктор, К., Боланьос, Л. М., Баетге, Н., Дэви-Мартин, К., Вестберри, Т. К., Бейтс, Т. С., Белл, Т. Г., Бидл, К. Д., Босс, Э. С., Брукс, С. Д., Кэрнс, Б., Карлсон, К., Холзи, К., Харви, Э. Л., Ху, К., Карп-Босс, Л., Клеб, М., Менден-Дойер, С., Морисон, Ф., Куинн, П.К., Скарино, А.Дж., Андерсон, Б., Чоудхари, Дж., Кросби, Э., Ферраре, Р., Волос, Дж. У., Ху, Ю., Янц, С., Редеманн, Дж., Зальцман, Э., Шук, М., Сигель, Д. А., Вистхалер, А., Мартин, М. Ю. и Зиемба, Л.: Исследование аэрозолей и морской экосистемы в Северной Атлантике. (NAAMES): Обзор научных мотивов и миссии, Front. Мар. Наук, 6, 1–25, https://doi.org/10.3389/fmars.2019.00122, 2019.
Беллуэн, Н., Куаас, Дж., Гриспердт, Э., Кинн, С., Стир, П., Уотсон-Пэррис, Д., Баучер, О., Карслав, К.С., Кристенсен, М., Даниау, А.-Л., Дюфрен, Ж.-Л., Фейнгольд, Г., Фидлер, С., Форстер, П., Геттельман, А., Хейвуд, Дж. М., Ломанн, У., Малавель, Ф., Мауритсен, Т., Маккой, Д. Т., Myhre, G., Mülmenstädt, J., Neubauer, D., Possner, A., Rugenstein, М., Сато, Ю., Шульц, М., Шварц, С. Э., Сурдеваль, О., Сторельвмо, Т., Толл В., Винкер Д. и Стивенс Б.: Ограничение глобального аэрозольного излучения. Вынужденное изменение климата, Rev. Geophys., 58, e2019RG000660, https://doi.org/10.1029/2019rg000660, 2020.
Борис Р.Д., Ловенталь, Д. Х., Ветцель, М. А., Эррера, Ф., Гонсалес, А., и Харрис Дж .: Химические и микрофизические свойства морского стратиформного облака. в Северной Атлантике J. Geophys. Рес.-Атмос., 103, 22073–22085, https://doi.org/10.1029/98JD02087, 1998.
Баучер, О. и Ломанн, У .: Эффект альбедо сульфат-CCN-облако, Tellus B, 47, 281–300, https://doi.org/10.3402/tellusb.v47i3.16048, 1995.
Бауман, А. Ф., Ли, Д. С., Асман, В. А. Х., Дентенер, Ф. Дж., Ван дер Хук, К. В., и Оливье, Дж. Дж. Дж .: Глобальный кадастр выбросов с высоким разрешением для аммиак, Global Biogeochem. Cy., 11, 561–587, https://doi.org/10.1029/97GB02266, 1997.
Чарлсон, Р. Дж., Лавлокт, Дж. Э., Андреэ, М. О., и Уоррен, С. Г.: Oceanic фитопланктон, сера в атмосфере, альбедо облаков и климат, природа, 326, 655–661, https://doi.org/10.1038/326655a0, 1987.
Чарлсон, Р. Дж., Шварц, С. Е., Хейлз, Дж. М., Сесс, Р. Д., Коакли, Дж. Дж., Хансен, Дж. Э. и Хофманн, Д. Дж .: Воздействие на климат антропогенными аэрозолями. Science, 117, 423–430, https: // doi.org / 10.1126 / science.255.5043.423, 1992.
Чен, Ю. и Пеннер, Дж. Э .: Анализ неопределенности для оценок первого косвенного аэрозольного эффекта, Atmos. Chem. Phys., 5, 2935–2948, https://doi.org/10.5194/acp-5-2935-2005, 2005.
Чен, Ю.-К., Кристенсен, М.В., Сюэ, Л., Сорошиан, А., Стивенс, Г. Л., Расмуссен, Р. М., и Сайнфельд, Дж. Х .: Появление более низкого альбедо облаков на траекториях кораблей, Атмосфер. Chem. Phys., 12, 8223–8235, https://doi.org/10.5194/acp-12-8223-2012, 2012.
Chuang, C.К. и Пеннер, Дж. Э .: Воздействие антропогенного сульфата на облака зарождение капель и оптические свойства, Tellus B, 47, 566–577, https://doi.org/10.1034/j.1600-0889.47.issue5.4.x, 1995.
Чуанг, К. К., Пеннер, Дж. Э. и Эдвардс, Л. Л.: Очистка зародышей Частицы дыма и смоделированные распределения размеров капель в большой биомассе Пожары, J. Atmos. Наук, 49, 1264–1276, https://doi.org/10.1175/1520-0469(1992)049<1264:NSOSPA>2.0.CO;2, 1992.
Коггон, М.М., Сорушян, А., Ван З., Меткалф, А. Р., Фроссар, А. А., Лин, Дж. Дж., Крейвен, Дж. С., Ненес, А., Йонссон, Х. Х., Рассел, Л. М., Флаган, Р. К. и Сайнфельд, Дж. : анализ вклада выбросов судов в свойства морских аэрозолей и облаков, Atmos. Chem. Phys., 12, 8439–8458, https://doi.org/10.5194/acp-12-8439-2012, 2012.
Коггон, М.М., Сорошян, А., Ван, З., Крейвен, Дж. С., Меткалф , AR, Линь, Дж. Дж., Ненес, А., Йонссон, Х. Х., Флаган, Р.К. и Сайнфельд Дж. Х .: Наблюдения континентальных биогенных воздействий на морской аэрозоль и облака у побережья Калифорнии, J. Geophys. Res.-Atmos., 119, 6724–6748, https://doi.org/10.1002/2013JD021228, 2014.
Кросби, Э., Ван, З., Сорошиан, А., Чуанг, П. Ю., Крейвен, Дж. С., Коггон, М. М., Брунке, М., Зенг, X., Йонссон, Х., Вудс, Р. К., Флаган, Р. К., и Сейнфельд, Дж. Х .: Расчистка слоисто-кучевых облаков и заметные термодинамические и Контрасты аэрозолей на границе раздела ясно – облачность, J.Атмос. Наук, 73, 1083–1099, https://doi.org/10.1175/JAS-D-15-0137.1, 2016.
Дадашазар, Х., Ван, З., Кросби, Э., Брунке, М., Цзэн, X. , Йонссон, Х., Вудс, Р. К., Флаган, Р. К., Сайнфелд, Дж. Х. и Сорушян, А .: Связь между гигантскими частицами морской соли и облаками, выведенная из физико-химические данные самолета, J. Geophys. Res., 122, 3421–3434, https://doi.org/10.1002/2016JD026019, 2017.
Дадашазар, Х., Ма, Л. и Сорушян, А .: Источники загрязнения и взаимосвязь между аэрозолем и химией осадков в центральном Калифорнийский сайт, Sci.Total Environ., 651, 1776–1787, https://doi.org/10.1016/j.scitotenv.2018.10.086, 2019.
Даймонд, М.С., Добрацки, А., Фрейтаг, С., Смолл Грисволд, Дж. Д., Хейккила, А., Хауэлл, С. Г., Качараб, М. Е., Подольске, Д. Р., Саид, П. Е. и Вуд, Р.: увлечение, зависящее от времени. дыма представляет собой проблему наблюдений для оценки взаимодействия аэрозоля и облака над юго-восточной частью Атлантического океана, Атмос. Chem. Phys., 18, 14623–14636, https://doi.org/10.5194/acp-18-14623-2018, 2018.
Facchini, M.К., Мирча, М., Фуцци, С., Чарлсон, Р. Дж .: Альбедо облаков. усиление поверхностно-активными органическими растворами в растущих каплях, Природа, 401, 257–259, https://doi.org/10.1038/45758, 1999.
Фейнголд, Г., Фриш, А.С., Стивенс, Б., и Коттон, У. взаимосвязь между турбулентностью облаков, образованием капель и моросью, если смотреть доплеровским радаром, микроволновым радиометром и лидаром, J. Geophys. Res.-Atmos., 104, 22195–22203, https://doi.org/10.1029/1999JD
2, 1999.
Feingold, G., Коттон, В. Р., Крейденвейс, С. М., и Дэвис, Дж. Т .: Воздействие ядер конденсации гигантских облаков при образовании мороси в слоисто-кучевых облаках: Влияние на радиационные свойства облаков, J. Atmos. Наук, 56, 4100–4117, https://doi.org/10.1175/1520-0469(1999)056<4100:TIOGCC>2.0.CO;2, 1999.
Фунтукис, К. и Ненес, А.: Продолжение разработки облачной капли Параметризация формации для моделей глобального климата, J. Geophys. Res.-Atmos., 110, D11212, https://doi.org/10.1029/2004JD005591, 2005.
Фройнд, Р. Дж., Уилсон, В. Дж., И Мор, Д. Л .: Статистические методы, 3-е изд., Academic Press, Burlington, MA, 824 pp., 2010.
Геларо, Р., Маккарти, В., Суарес, М. Дж., Тодлинг, Р., Молод, А., Такач, Л., Рэндлс, К. А., Дарменов, А., Босилович, М. Г., Райхл, Р., Варган, К., Кой, Л., Куллатер, Р., Дрейпер, К., Акелла, С., Бучард, В., Конати, А., да Силва, А. М., Гу, В., Ким, Г. К., Костер, Р., Луччези, Р., Меркова, Д., Нильсен, Дж. Э., Партика, Г., Поусон, С., Путман, В., Ринекер, М., Шуберт С. Д., Сенкевич М. и Чжао Б. Ретроспектива современной эпохи. анализ для исследований и приложений, версия 2 (MERRA-2), J. Climate, 30, 5419–5454, https://doi.org/10.1175/JCLI-D-16-0758.1, 2017.
Гербер, Х., Арендс, Б.Г., и Акерман, А.С.: Новый микрофизический датчик для использование самолетов, Атмос. Res., 31, 235–252, https://doi.org/10.1016/0169-8095(94)-9, 1994.
Гербер, Х., Фрик, Г., и Роди, А.Р .: Наземные FSSP и PVM измерения жидкой воды, Дж.Атмос. Океан. Тех., 16, корп. 1143–1149, https://doi.org/10.1175/1520-0426(1999)016<1143:GBFAPM>2.0.CO;2, 1999.
Ghan, SJ, Leung, LR, and Easter, RC: Prediction облачной капли число в модели общей циркуляции, J. Geophys. Res., 102, 21777–21794, https://doi.org/10.1029/97JD01810, 1997.
Ган, С. Дж., Абдул-Раззак, Х., Ненес, А., Мин, Ю., Лю, X., Овчинников, М., Шипвей Б., Месхидзе Н., Сюй Дж. И Ши X .: Зарождение капель: Физически обоснованные параметризации и сравнительная оценка, J.Adv. Модель. Earth Syst., 3, 1–34, https://doi.org/10.1029/2011ms000074, 2011.
Гонг, С.Л .: Параметризация функции источника аэрозоля морской соли для суб- и сверхмикронные частицы, Global Biogeochem. Cy., 17, 1097, г. https://doi.org/2003gb002079, 2003.
Grosvenor, D. P., Sourdeval, O., Zuidema, P., Ackerman, A., Alexandrov, M. Д., Беннарц, Р., Берс, Р., Кэрнс, Б., Чиу, Дж. К., Кристенсен, М., Денеке, Х., Даймонд, М., Фейнгольд, Г., Фридлинд, А., Хюнербейн, А., Книст, К., Коллиас, П., Маршак, А., Маккой, Д., Мерк, Д., Пейнемал, Д., Рауш, Дж., Розенфельд, Д., Русшенберг, Х., Зайферт, П., Синклер, К., Стир П., ван Диденховен Б., Вендиш М., Вернер Ф., Вуд Р., Чжан, З. и Кваас Дж .: Дистанционное зондирование концентрации числа капель в тепле. Облака: Обзор текущего состояния знаний и перспектив, Rev. Geophys., 56, 409–453, https://doi.org/10.1029/2017RG000593, 2018.
Hegg, D.A .: Влияние газовой фазы HNO 3 и NH 3 на микрофизические процессы в атмосферные облака, Geophys.Res. Lett., 27, 2201–2204, https://doi.org/10.1029/1999GL011252, 2000.
Хегг Д. А. и Хоббс П. В. Химия облачной воды и производство сульфаты в облаках, Атмос. Environ., 15, 1597–1604, https://doi.org/10.1016/0004-6981(81)
-X, 1981.
Хегг, Д. А. и Хоббс, П. В. Химия сульфатов и нитратов в кучевых формах облака, Атмос. Окружающая среда, 20, 901–909, https://doi.org/10.1016/0004-6981(86)
-X, 1986.
Хегг, Д. А., Ферек, Р. Дж., и Хоббс, П. Ф .: Рассеяние света и облака активность ядра конденсации сульфатного аэрозоля, измеренная на северо-востоке Атлантический океан, Дж.Geophys. Res.-Atmos., 98, 14887–14894, https://doi.org/10.1029/93JD01615, 1993.
Hersey, SP, Sorooshian, A., Murphy, SM, Flagan, RC, and Seinfeld, JH: Гигроскопичность аэрозолей в морской атмосфере: исследование закрытия с использованием высокого временного разрешения, множественной RH DASH-SP и C с разрешением по размеру -ToF-Данные AMS, Атмос. Chem. Phys., 9, 2543–2554, https://doi.org/10.5194/acp-9-2543-2009, 2009.
Hudson, PK, Murphy, DM, Cziczo, DJ, Thomson, DS, de Gouw, JA, Варнеке, К., Холлоуэй, Дж., Йост, Х. Дж., И Хюблер, Г.: сжигание биомассы. измерения частиц: характеристика состава и химическая обработка, J. Geophys. Res.-Atmos., 109, D23S27, https://doi.org/10.1029/2003JD004398, 2004.
Межправительственная группа экспертов по изменению климата: изменение климата 2007: физическое Научная основа, вклад Рабочей группы I в четвертую оценку Отчет Межправительственной группы экспертов по изменению климата, Кембридж University Press, Кембридж, Соединенное Королевство и Нью-Йорк, Нью-Йорк, США, 996 стр., 2007.
Межправительственная группа экспертов по изменению климата: изменение климата, 2013 г .: физическое Научная основа, вклад Рабочей группы I в пятую оценку Отчет Межправительственной группы экспертов по изменению климата, Кембридж University Press, Кембридж, Соединенное Королевство и Нью-Йорк, Нью-Йорк, США, 1535 стр., 2013.
Юнг, Э., Альбрехт, Б.А., Йонссон, Х.Х., Чен, Ю.-К., Сайнфельд, Дж. Х., Сорошиан, А., Меткалф, А.Р., Сонг, С., Фанг, М., и Рассел, Л.М.: Осадки эффекты гигантских облачных ядер конденсации, искусственно введенных в слоисто-кучевые облака, Атмос.Chem. Phys., 15, 5645–5658, https://doi.org/10.5194/acp-15-5645-2015, 2015.
Jickells, TD, An, ZS, Andersen, KK, Baker, AR, Bergametti, G ., Брукс, Н., Цао, Дж. Дж., Бойд, П. У., Дуче, Р. А., Хантер, К. А., Кавахата, Х., Кубилай Н. и Лисс П. С. Глобальные связи железа между пустыней Пыль, биогеохимия океана и климат, Наука, 308, 67–71, https://doi.org/10.1126/science.1105959, 2005.
Кахане, Л. Х .: Основы регрессии, 2-е изд., Sage Publications, Thousand Oaks, CA, 234 с., 2008.
Кох, Д. и Дель Генио, А. Д .: Полупрямое воздействие черного углерода на облачный покров: обзор и синтез, Atmos. Chem. Phys., 10, 7685–7696, https://doi.org/10.5194/acp-10-7685-2010, 2010.
Кульмала М., Лааксонен А., Корхонен П., Весала Т. , Ахонен, Т., и Барретт, Дж. К .: Влияние атмосферных паров азотной кислоты на облако. активация ядра конденсации, J. Geophys. Res., 98, 22949–22958, https://doi.org/10.1029/93JD02070, 1993.
Лэнс, С., Ненес, А., Маззолени, К., Дубей, М. К., Гейтс, Х., Варутбангкул, В., Рисман, Т. А., Мерфи, С. М., Сорушян, А., Флаган, Р. К., Сайнфельд, Дж. Х., Фейнгольд, Г., Йонссон, Х. Х .: Активность ядер конденсации облаков. закрытие и кинетика роста капель хьюстонского аэрозоля во время залива Исследование состава атмосферы и климата Мексики (GoMACCS), J. Geophys. Res., 114, D00F15, https://doi.org/10.1029/2008jd011699, 2009.
Leaitch, W. R., Strapp, J. W., Wiebe, H. A., Anlauf, K. G. и Isaac, G.А .: Химические и микрофизические исследования летних облаков без осадков в г. Онтарио, Канада, J. Geophys. Рес.-Атмос., 91, 11821–11831, https://doi.org/10.1029/JD091iD11p11821, 1986.
Лиич, У. Р., Исаак, Г. А., Страпп, Дж. У., Баник, К. М. и Вибе, Х. А .: Связь между числовыми концентрациями облачных капель и антропогенное загрязнение: наблюдения и климатические последствия, J. Geophys. Res.-Atmos., 97, 2463–2474, https://doi.org/10.1029/91JD02739, 1992.
Leaitch, W.Р., Банич, К. М., Исаак, Г. А., Кутюр, М. Д., Лю, П. С. К., Гультепе, И., Ли, С. М., Клейнман, Л., Даум, П. Х., и Макферсон, Дж. И .: Физико-химические наблюдения в морских слоях на севере 1993 г. Атлантический региональный эксперимент: факторы, влияющие на количество облачных капель концентрации, J. Geophys. Рес.-Атмос., 101, 29123–29135, https://doi.org/10.1029/96JD01228, 1996.
Ловенталь Д. Х. и Борис Р. Д. Источники микрофизических вариаций в морские слоистые облака в Северной Атлантике, Geophys.Res. Lett., 27, 1491–1494, https://doi.org/10.1029/1999GL010992, 2000.
Ловенталь, Д. Х., Борис, Р. Д., Чоулартон, Т. У., Бауэр, К. Н., Флинн, М. Дж., Галлахер М. У.: Параметризация облачной капли-сульфата. отношения, Атмос. Окружающая среда, 38, 287–292, https://doi.org/10.1016/j.atmosenv.2003.09.046, 2004.
Лу, М. Л., Сорушян, А., Йонссон, Х. Х., Фейнгольд, Г., Флаган, Р. К., и Сейнфельд, Дж. Х .: Взаимосвязь морских слоисто-кучевых аэрозольных облаков в Эксперимент MASE-II: Восприимчивость к осадкам в морской части восточной части Тихого океана слоисто-кучевые, J.Geophys. Рес.-Атмосфера, 114, 1–11, https://doi.org/10.1029/2009JD012774, 2009.
Ма, Л., Дадашазар, Х., Браун, Р. А., Макдональд, А. Б., Агдам, М. А., Модлин, Л.С., и Сорошян, А .: Разрешенные по размеру характеристики водорастворимые твердые частицы в прибрежной зоне: идентификация источника, влияние лесных пожаров и суточная изменчивость, Атмос. Окружающая среда, 206, 72–84, https://doi.org/10.1016/j.atmosenv.2019.02.045, 2019.
Макдональд, А. Б., Дадашазар, Х., Чуанг, П. Ю., Кросби, Э., Ван, Х., Ван З., Йонссон Х. Х., Флаган Р. К., Сайнфелд Дж. Х. и Сорушян А .: Характерные вертикальные профили состава облачной воды в морской среде Слоисто-кучевые облака и их связь с осадками, J. Geophys. Res.-Atmos., 123, 3704–3723, https://doi.org/10.1002/2017JD027900, 2018.
Марди, А. Х., Дадашазар, Х., Макдональд, А. Б., Браун, Р. А., Кросби, Э., Сиань П., Торсен Т. Дж., Коггон М. М., Фенн М. А., Ферраре Р. А., Волосы, Дж. У., Вудс, Р. К., Йонссон, Х. Х., Флаган, Р. К., Сайнфельд, Дж. Х. и Сорошян, А .: Шлейфы сжигания биомассы в окрестностях Калифорнии Побережье: определение физико-химических свойств в воздухе, нагрев Скорости и пространственно-временные особенности, J. Geophys. Рес.-Атмос., 123, 13560–13582, https://doi.org/10.1029/2018JD029134, 2018.
Марди, А. Х., Дадашазар, Х., Макдональд, А. Б., Кросби, Э., Коггон, М. М., Агдам, М. А., Вудс, Р. К., Йонссон, Х. Х., Флаган, Р. К., Сайнфельд, Дж. Х., и Сорошян А.: Влияние горения биомассы на каплю слоисто-кучевых облаков Характеристики, скорость дождя и состав, J. Geophys. Res.-Atmos., 124, 12301–12318, https://doi.org/10.1029/2019JD031159, 2019.
Мацумото, К., Танака, Х., Нагао, И. и Ишизака, Ю.: Вклад сульфат в виде твердых частиц и органический углерод для замутнения ядер конденсации в морская атмосфера, геофизика. Res. Lett., 24, 655–658, https://doi.org/10.1029/97GL00541, 1997.
Модлин, Л. К., Ван, З., Йонссон, Х. Х., Сорушян, А.: Влияние лесные пожары, связанные с аэрозольным составом с разрешенным размером частиц на прибрежной территории Калифорнии, Атмос. Environ., 119, 59–68, https://doi.org/10.1016/j.atmosenv.2015.08.039, 2015.
Маккой, Д. Т., Бендер, Ф. А.-М., Морманн, Дж. К. К., Хартманн, Д. Л., Вуд, Р., Гросвенор Д. П .: Первый косвенный эффект глобального аэрозольного облака оценено с использованием MODIS, MERRA и AeroCom, J. Geophys. Рес.-Атмос., 122, 1779–1796, https://doi.org/10.1002/2016JD026141, 2017.
Маккой Д.Т., Бендер Ф.А.-М., Гросвенор, Д. П., Морманн, Дж. К., Хартманн, Д. Л., Вуд, Р., и Филд, П. Р.: Прогнозирование десятилетних тенденций в изменении количества облачных капель с использованием реанализа и спутниковых данных, Atmos. Chem. Phys., 18, 2035–2047, https://doi.org/10.5194/acp-18-2035-2018, 2018.
Medina, J., Nenes, A., Sotiropoulou, REP, Cottrell, LD, Ziemba , Л. Д., Бекман, П. Дж., И Гриффин, Р. Дж .: Закрытие ядер конденсации облаков. во время Международного консорциума атмосферных исследований в области транспорта и Кампания Transformation 2004: эффекты размерной композиции, J.Geophys. Рес.-Атмос., 112, Д10С31, https://doi.org/10.1029/2006JD007588, 2007.
Менон С. и Саксена В. К .: Роль сульфатов в региональных облаках — климат взаимодействия, Атмос. Res., 47, 299–315, https://doi.org/10.1016/S0169-8095(98)00057-X, 1998.
Менон, С., Генио, А. Д. Дел, Кох, Д., и Целиудис, Г.: Моделирование методом GCM Косвенный эффект аэрозоля: чувствительность к параметризации облаков и Бремя аэрозолей, J. Atmos. Наук, 59, 692–713, https://doi.org/10.1175/1520-0469(2002)059<0692:gsotai>2.0.co; 2, 2002.
Ненес А., Чарлсон Р. Дж., Факкини М. К., Кульмала М., Лааксонен А. и Сайнфельд, Дж. Х .: Может ли химическое воздействие на количество облачных капель соперничать с первый косвенный эффект ?, Geophys. Res. Lett., 29, 1848 г., https://doi.org/10.1029/2002gl015295, 2002.
Ненес, А. и Сайнфельд, Дж. Х .: Параметризация образования облачных капель в моделях глобального климата, J. Geophys. Рес.-Атмосфера, 108, 1–14, https://doi.org/10.1029/2002jd002911, 2003.
Новаков Т. и Пеннер Дж.E .: Большой вклад органических аэрозолей в концентрации ядер конденсации облаков, Природа, 365, 823–826, https://doi.org/10.1038/365823a0, 1993.
Новаков Т., Ривера-Карпио К., Пеннер Дж. Э. и Роджерс К. Ф .: Эффект антропогенных сульфатных аэрозолей на концентрации капель морских облаков, Теллус В, 46, 132–141, https://doi.org/10.3402/tellusb.v46i2.15758, 1994.
Партридж, Д.Г., Вругт, Дж. А., Танвед, П., Экман, AML, Струтерс, Х. и Сорошиан, А.: Обратное моделирование взаимодействий облака и аэрозоля — Часть 2: Тесты на чувствительность жидкофазных облаков с использованием цепного метода Маркова Подход к моделированию на основе Карло, Atmos.Chem. Phys., 12, 2823–2847, https://doi.org/10.5194/acp-12-2823-2012, 2012.
Paulot, F., Jacob, DJ, Johnson, MT, Bell, TG, Baker, AR, Кин, У. К., Лима, И. Д., Дони, С. К., и Сток, К. А.: Глобальные выбросы в океан. аммиака: ограничения по наблюдениям за морской водой и атмосферой, Global Biogeochem. Cy., 28, 1165–1178, https://doi.org/10.1002/2015GB005106, 2015.
Прабхакар, Г., Эрвенс, Б., Ван, З., Модлин, Л. К., Коггон, М. М., Йонссон, Х. Х., Сайнфельд, Дж.Х., Сорушян А. Источники нитратов в слоисто-кучевые облака воды: измерения в воздухе во время E-PEACE 2011 г. и Исследования NiCE 2013, Атмос. Окружающая среда, 97, 166–173, https://doi.org/10.1016/j.atmosenv.2014.08.019, 2014.
Прингл, К. Дж., Карслав, К. С., Спраклен, Д. В., Манн, Г. М., и Чипперфилд, М. П .: Взаимосвязь между аэрозольными и количественными концентрациями капель облаков в глобальной модели аэрозольной микрофизики, Atmos. Chem. Phys., 9, 4131–4144, https://doi.org/10.5194/acp-9-4131-2009, 2009.
Пуэшель, Р. Ф., Валин, К. К., Кастильо, Р. К., Кадлечек, Дж. А., и Ганор, E .: Аэрозоли в загрязненных и незагрязненных воздушных массах: перенос на большие расстояния и воздействие на облака, J. Appl. Meteorol. Клим., 25, 1908–1917, г. https://doi.org/10.1175/1520-0450(1986)025<1908:AIPVNA>2.0.CO;2, 1986.
Пай, ХОТ, Ненес, А., Александр, Б., Олт, А.П., Барт, М.С., Клегг, С.Л., Коллетт-младший, Дж.Л., Фейи, К.М., Хенниган, Си-Джей, Херрманн, Х., Канакиду, М., Келли, JT, Ку, И.-Т., Макнил, В.Ф., Ример, Н., Шефер, Т., Ши, Г., Тилгнер, А., Уокер, Дж. Т., Ван, Т., Вебер, Р., Син, Дж., Завери, Р.А. , и Зуэнд, А .: Кислотность атмосферных частиц и облаков, Атмос. Chem. Phys., 20, 4809–4888, https://doi.org/10.5194/acp-20-4809-2020, 2020.
Quinn, PK, Coffman, DJ, Johnson, JE, Upchurch, LM, and Bates, Т. С .: Небольшая часть ядер конденсации морских облаков состоит из морских брызг. аэрозоль, нат. Geosci., 10, 674–679, https://doi.org/10.1038/ngeo3003, 2017 г.
Рэндлс, К. А., да Силва, А. М., Бушард, В., Коларко, П. Р., Дарменов, А., Говиндараджу, Р., Смирнов, А., Холбен, Б., Ферраре, Р., Волос, Дж., Шинозука, Й., Флинн К. Дж .: Реанализ аэрозолей MERRA-2, начиная с 1980 года. Часть I: Описание системы и оценка усвоения данных, J. Climate, 30, 6823–6850, https://doi.org/10.1175/JCLI-D-16-0609.1, 2017.
Рид, Дж. С., Хоббс, П. В., Ферек, Р. Дж., Блейк, Д. Р., Мартинс, Дж. В., Данлэп М. Р. и Лисс Ч. Физические, химические и оптические свойства региональные туманы, в которых преобладает дым в Бразилии, J.Geophys. Res.-Atmos., 103, 32059–32080, https://doi.org/10.1029/98JD00458, 1998.
Рассел, Л. М., Сорошян, А., Сайнфельд, Дж. Х., Альбрехт, Б. А., Ненес, А., Алм, Л., Чен, Ю.-К., Коггон, М., Крейвен, Дж. С., Флаган, Р. К., Фроссар, А. А., Йонссон, Х., Юнг, Э., Лин, Дж. Дж., Меткалф, А. Р., Модини, Р., Мюльменштадт, Дж., Робертс, Г. К., Шинглер, Т., Сонг, С., Ван, З., и Wonaschütz, A .: Восточно-тихоокеанский эксперимент с испускаемыми облаками аэрозолей, B. Am. Meteorol. Soc., 94, 709–729, https: // doi.org / 10.1175 / BAMS-D-12-00015.1, 2013.
Саксена В. К. и Менон С. Охлаждение, вызванное сульфатами, на юго-востоке США: Наблюдательная оценка, Geophys. Res. Lett., 26, 2489–2492, https://doi.org/10.1029/1999GL
5, 1999.Шлоссер, Дж. С., Браун, Р. А., Брэдли, Т., Дадашазар, Х., Макдональд, А. Б., Алдаиф, А. А., Азади Агдам, М., Хоссейн Марди, А., Сиань, П., и Сорошян, А .: Анализ данных о составе аэрозолей для Western United. Лесные пожары в штатах в период 2005–2015 гг .: выбросы пыли, истощение хлоридов и Наиболее усовершенствованные компоненты аэрозолей, J.Geophys. Рес.-Атмос., 122, 8951–8966, https://doi.org/10.1002/2017JD026547, 2017.
Шлоссер, Дж. С., Дадашазар, Х., Эдвардс, Э.-Л., Марди, А. Х., Прабхакар, Г., Шталь, К., Йонссон, Х. Х. и Сорушян, А .: Отношения между супермикрометрический аэрозоль морской соли и условия морского пограничного слоя: выводы из повторяющихся идентичных схем полета, J. Geophys. Рес.-Атмос., 125, https://doi.org/10.1029/2019JD032346, 2020.
Сейнфельд, Дж. Х. и Пандис, С. Н .: Атмосферная химия и физика, 3-е изд., New York, NY, 1152 pp., 2016.
Сильва, П. Дж., Лю, Д. Ю., Нобл, К. А., и Пратер, К. А.: Размер и химический состав. характеристика отдельных частиц в результате сжигания биомассы местный южнокалифорнийский вид, Environ. Sci. Технол., 33, 3068–3076, https://doi.org/10.1021/es980544p, 1999.
Сорошян А., Лу, М. Л., Брехтель, Ф. Дж., Йонссон, Х., Фейнгольд, Г., Flagan, R.C., и Seinfeld, J.H .: Об источнике аэрозолей органических кислот. слои над облаками, Environ.Sci. Technol., 41, 4647–4654, https://doi.org/10.1021/es0630442, 2007.
Сорошян А., Падро Л. Т., Ненес А., Фейнгольд Г., МакКомиски А., Херси, С. П., Гейтс, Х., Йонссон, Х. Х., Миллер, С. Д., Стивенс, Г. Л., Флаган Р. К. и Сайнфельд Дж. Х .: О связи между биотой океана. выбросы, аэрозоли и морские облака: воздушные, наземные и спутниковые измерения у побережья Калифорнии, Global Biogeochem. Cy., 23, г. 1–15, https://doi.org/10.1029/2009GB003464, 2009.
Сорушян А., Макдональд, А. Б., Дадашазар, Х., Бейтс, К. Х., Коггон, М. М., Крейвен, Дж. С., Кросби, Э., Эдвардс, Э.-Л., Херси, С. П., Ходас, Н., Лин, Дж. Дж., Хоссейн Марди, А., Марти, А. Н., Модлин, Л. К., Меткалф, А. Р., Мерфи, С. М., Падро, Л. Т., Прабхакар, Г., Риссман, Т. А., Шлоссер, Дж., Шинглер Т., Варутбангкул В., Ван З., Вудс Р. К., Чуанг П. Ю., Ненес, А., Йонссон, Х. Х., Флаган, Р. К., и Сайнфельд, Дж. Х .: многолетний набор данных о взаимодействии аэрозолей, облаков, осадков и метеорологии для морских слоисто-кучевые облака, фигшаре, https: // doi.org / 10.6084 / m9.figshare.5099983.v10, 2017.
Сорушян А., Макдональд А. Б., Дадашазар Х., Бейтс К. Х., Коггон М. М., Крейвен, Дж. С., Кросби, Э., Херси, С. П., Ходас, Н., Лин, Дж. Дж., Негрон Марти, А., Модлин, Л. К., Меткалф, А. Р., Мерфи, С. М., Падро, Л. Т., Прабхакар, Г., Рисман, Т. А., Шинглер, Т., Варутбангкул, В., Ван, З., Вудс, Р. К., Чуанг, П. Ю., Ненес, А., Йонссон, Х. Х., Flagan, R.C., и Seinfeld, J.H .: набор данных за несколько лет по взаимодействие аэрозолей, облаков, осадков и метеорологии для морских слоисто-кучевые облака, Sci.Данные, 5, 1–13, https://doi.org/10.1038/sdata.2018.26, 2018.
Сорушян А., Андерсон Б., Бауэр С. Э., Браун Р. А., Кэрнс Б., Кросби, Э., Дадашазар, Х., Дискин, Г., Ферраре, Р., Флаган, Р. К., Волосы, Дж., Хостетлер, К., Йонссон, Х. Х., Клеб, М. М., Лю, Х., Макдональд, А. Б., Маккомиски, А., Мур, Р., Пейнемал, Д., Рассел, Л. М., Сайнфельд, Дж. Х., Шук, М., Смит, В. Л., Торнхилл, К., Целиудис, Г., Ван, Х., Цзэн, X., Чжан Б., Зиемба Л. и Зуидема П .: Аэрозоль – облако – метеорология. взаимодействие с воздушными полевыми исследованиями: использование уроков, извлеченных из U.S. West Coast в проекте активировать у восточного побережья США, B. Am. Meteorol. Soc., 100, 1511–1528, https://doi.org/10.1175/BAMS-D-18-0100.1, 2019.
Сорошян, А., Коррал, А.Ф., Браун, Р.А., Кэрнс, Б., Кросби, Э., Ферраре, Р., Волос, Дж., Клеб, М.М., Хоссейн Марди, А., Маринг, Х., МакКомиски, А., Мур, Р., Пейнемал, Д., Скарино, А. Дж., Шлоссер, Дж., Шинглер, Т., Шук, М., Ван, Х., Зенг, X., Зиемба, Л. и Зуидема, П. .: Атмосферные исследования в западной части региона Северной Атлантики и восточном побережье Северной Америки: обзор прошлой работы и предстоящих задач, J.Geophys. Res. Atmos., 125, e2019JD031626, DOI: 10.1029 / 2019JD031626, 2020.
Strapp, J. W., Leaitch, W. R., and Liu, P. S. K .: Hydrated and dry измерения распределения размеров аэрозолей с помощью систем измерения частиц Зонд FSSP-300 и зонд PCASP-100X, очищенный от льда, J. Atmos. Океан. Technol., 9, 548–555, https://doi.org/10.1175/1520-0426(1992)009<0548:HADASD>2.0.CO;2, 1992.
Twomey, S .: Ядра естественной части образования облаков II: The пересыщение в естественных облаках и изменение облачной капли концентрация, Geofis.Pura e Appl., 43, 243–249, https://doi.org/10.1007/BF01993560, 1959.
Туми, С .: Влияние загрязнения на коротковолновое альбедо облаков, Дж. Атмос. Наук, 34, 1149–1152, https://doi.org/10.1175/1520-0469(1977)034<1149:TIOPOT>2.0.CO;2, 1977.
Ван Дингенен, Р., Раес, Ф., и Йенсен, Н.Р .: Доказательства в пользу антропогенный влияние на числовую концентрацию и сульфатность облачно-обработанных аэрозольные частицы над Северной Атлантикой, J. Geophys. Res.-Atmos., 100, 21057–21067, https: // doi.org / 10.1029 / 95jd02141, 1995.
Wang, Z., Sorooshian, A., Prabhakar, G., Coggon, M. M., and Jonsson, H.H .: Влияние выбросов судоходства, суши и океана на слоисто-кучевые облака элементный состав воды в облаках во время полевой кампании E-PEACE 2011 г., Атмос. Environ., 89, 570–580, https://doi.org/10.1016/j.atmosenv.2014.01.020, 2014.
Ван, З., Мора Рамирес, М., Дадашазар, Х., Макдональд, А. Б., Кросби, Э., Бейтс, К. Х., Коггон, М. М., Крейвен, Дж. С., Линч, П., Кэмпбелл, Дж.Р., Азади Агдам, М., Вудс, Р. К., Йонссон, Х., Флаган, Р. К., Сайнфельд, Дж. Х., и Сорушян, А .: Контрастный состав облаков между связанными и развязанные облака морского пограничного слоя, J. Geophys. Рес.-Атмос., 121, 11679–11691, https://doi.org/10.1002/2016JD025695, 2016.
Уэст, Р.Э., Стир, П., Джонс, А., Джонсон, К.Е., Манн, Г.В., Беллуин, Н., Партридж, Д.Г., и Киплинг, З .: Важность изменчивости вертикальной скорости для оценки косвенных аэрозольных эффектов. , Атмос.Chem. Phys., 14, 6369–6393, https://doi.org/10.5194/acp-14-6369-2014, 2014.
Вонашютц, А., Коггон, М., Сорушян, А., Модини, Р., Фроссард, А.А., Альм, Л., Мюльменштадт, Дж., Робертс, Г.К., Рассел, Л.М., Дей, С., Брехтель, Ф.Дж. , и Сайнфельд, Дж. Х .: Гигроскопические свойства образующихся из дыма органических аэрозольных частиц, выбрасываемых в морскую атмосферу, Atmos. Chem. Phys., 13, 9819–9835, https://doi.org/10.5194/acp-13-9819-2013, 2013.
Сюэ, Х. и Фейнгольд, Г.: Моделирование влияния азотной кислоты. на свойства облака, J.Geophys. Res., 109, D18204, https://doi.org/10.1029/2004JD004750, 2004.
28 Кодекс США § 41 — Количество и состав цепей | Кодекс США | Закон США
Исторические и редакционные заметкиНа основании раздела 28, U.S.C. Издание 1940 г., §211, и раздел 864 раздела 48, USC, изд. 1940 г., Территории и островные владения (12 апреля 1900 г., глава 191, §35, 31 Закон 85; 3 марта 1911 г., глава 231, §116, 36 Закон 1131; 28 января 1915 г., глава 22, §§1, 2, 38 Закон 803; 2 марта 1917 г., глава 145, §42, 39 Закон.966; 13 февраля 1925 г., гл. 229, §§1, 13, 43 Стат. 936, 942; 31 января 1928 г., гл. 14, §1, 45 Стат. 54; 28 февраля 1929 г., гл. 363, § 1, 45 Стат. 1346; 17 мая 1932 г., гл. 190, 47 Стат. 158).
Форма раздела упрощена.
Округ Колумбия был добавлен в качестве отдельной цепи. Это соответствует решению Верховного суда Соединенных Штатов Америки, согласно которому Апелляционный суд округа Колумбия является окружным апелляционным судом в соответствии с Законом о передаче от сентября.14, 1922, гл. 305, 42 Стат. 837, включенный в Судебный кодекс как §238 (a), но отмененный законом от 13 февраля 1925 г., гл. 229, §13, 43 Стат. 942. (См. Swift and Co. против США, 1928, 48 S.Ct. 311, 276 U.S. 311, 72 L.Ed. 587.)
Признавая округ Колумбия отдельным округом, Верховный суд недавно использовал такой язык: «* * * одиннадцать округов, образующих единую федеральную судебную систему * * *». Comm’r. v. Bedford’s Estate, 65 S.Ct. 1157, на странице 1160, 325 U.S. 283, 89 L.Ed. 611.
См. Раздел 17 заголовка 28, USC, 1940 ed., Где говорится: «Для целей разделов 17–23 настоящего заголовка округ Колумбия считается судебным округом * * *», и действуйте от декабря 2006 г. 23, 1944, гл. 724, 58 Stat. 925, которая внесла поправки в раздел 215 заголовка 28, U.S.C., 1940 ed., Включенный в раздел 42 этого заголовка. Такая поправка предусматривала, что для целей указанного раздела 215 «Округ Колумбия считается судебным округом».
Многие другие акты Конгресса признали округ Колумбия отдельным округом.(См. Следующие акты; 24 августа 1937 г., глава 754, 50 Stat. 751; 11 февраля 1938 г., глава 25, 52 Stat. 28; 5 августа 1939 г., глава 433, 53 Stat. 1204). ; 7 августа 1939 г., глава 501, 53 Stat. 1223; 29 декабря 1942 г., глава 835, 56 Stat. 1094; 11 мая 1944 г., глава 192, 58 Stat. 218; 23 декабря 1944 г. , ch.724, 58 Stat.925.)
См. Также следующие акты, признающие Апелляционный суд округа Колумбия окружным апелляционным судом: 15 августа 1921 г., гл. 64, 42 Стат. 162; 5 июля 1935 г., гл. 372, 49 Стат. 454; Авг.24, 1937, гл. 754, 50 Стат. 751; 6 апреля 1942 г., гл. 210, 56 Стат. 198; 9 мая 1942 г., гл. 295, 56 Стат. 271. См. Также Правило 81 (d) Федеральных правил гражданского судопроизводства.
В следующих делах Верховный суд Соединенных Штатов признал статус Апелляционного суда округа Колумбия в качестве постоянного учреждения в рамках федеральной судебной системы: О’Донохью против Соединенных Штатов, 1933, 53 S.Ct . 740, 289 U.S. 516, 77 L.Ed. 1356; Федеральная торговая комиссия против Клеснера, 1927, 47 S.Кт. 557, 274 U.S. 145, 71 L.Ed. 972; Клэйборн-Аннаполис Ферри против Соединенных Штатов, 1932, 52 S.Ct. 440, 285 U.S. 382, 76 L.Ed. 808; США против California Canneries, 1929, 49 S.Ct. 423, 279 U.S. 553, 73 L.Ed. 838.
Аляска, Зона каналов и Виргинские острова были добавлены к 9-му, 5-му и 3-му округам соответственно, чтобы соответствовать разделу 1294 этого названия.
Некоторые положения раздела 864 раздела 48, U.S.C., 1940 ed., Были сохранены в указанном заголовке. Для тех, которые были включены в другие разделы этого пересмотренного названия, см. Таблицу распределения.
От редакции
Поправки1982 — Pub. L. 97–164 увеличил количество судебных округов с двенадцати до тринадцати за счет добавления Федерального округа, состоящего из всех федеральных судебных округов.
1980 — Pub. L. 96–452 заменил «двенадцать» на «одиннадцать» в тексте предыдущей таблицы, заменил «Район зоны канала» на «Алабама, зона канала, Флорида, Джорджия» в элементе, относящемся к пятому контуру, и добавил новый элемент, относящийся к одиннадцатый контур.
1951 — Закон, октябрь.31 января 1951 года в часть, касающуюся состава Девятого судебного округа, добавлена ссылка на Гуам.
Обязательства и связанные с ними дочерние компании
Дата вступления в силу поправки 1980 г.Паб. L. 96–452, §12, 14 октября 1980 г., 94 Stat. 1996 г., при условии, что:
«Настоящий Закон и поправки, внесенные этим Законом [вносящие поправки в этот раздел и разделы 44 и 48 настоящего заголовка, и вводящие в действие положения, изложенные в примечаниях к этому разделу], вступают в силу 1 октября 1981 г.» Прекращение действия Окружного суда США по району зоны каналаДля прекращения действия Окружного суда Соединенных Штатов по району зоны канала в конце «переходного периода», то есть 30-месячного периода, начинающегося окт.1 января 1979 года и до полуночи 31 марта 1982 года, см. Параграф 5 статьи XI Договора о Панамском канале 1977 года и разделы 2101 и 2201–2203 Pub. L. 96–70, раздел II, 27 сентября 1979 г., 93 Stat. 493, ранее относившиеся к разделам 3831 и 3841–3843, соответственно, раздела 22 «Международные отношения и половые отношения».
Комиссия по альтернативным структурам федеральных апелляционных судовПаб. L. 105–119, раздел III, §305, 26 ноября 1997 г., 111 Stat. 2491, учредившая Комиссию по структурным альтернативам для федеральных апелляционных судов, поручила Комиссии изучить разделение Соединенных Штатов на судебные округа, изучить структуру и согласованность системы Федерального апелляционного суда и сообщить президенту и Конгрессу свои рекомендации об изменениях, необходимых для оперативного и эффективно избавляться от большого количества дел в Федеральных апелляционных судах в соответствии с фундаментальными концепциями справедливости и надлежащей правовой процедуры, предусматривающими членство в Комиссии и вознаграждение членов и сотрудников, утвержденные ассигнования и предусматривающие прекращение деятельности Комиссии через 90 дней после представления ее отчета.
Назначение судей и порядок рассмотрения нерассмотренных дел в связи с реорганизацией Апелляционного суда пятого округаПаб. L. 96–452, §§5–10, 14 октября 1980 г., 94 Stat. 1994, 1995, при условии, что:
«Разд. 5. Каждый окружной судья, находящийся на регулярной действующей службе бывшего пятого округа, официальная должность которого на день до даты вступления в силу настоящего Закона [окт. 1, 1981] — «(1)находится в Луизиане, Миссисипи или Техасе и назначен окружным судьей нового пятого округа; и
«(2)находится в Алабаме, Флориде или Джорджии и назначен окружным судьей одиннадцатого округа.
«Разд. 6.Каждый судья, который является старшим судьей бывшего пятого округа на день до даты вступления в силу настоящего Закона [окт. 1, 1981] может выбрать назначение в новый пятый округ или в одиннадцатый округ и должен уведомить директора административного офиса судов Соединенных Штатов о таком избрании.
«Разд. 7. Старшинство каждого судьи: «(1), назначенный в соответствии со статьей 5 настоящего Закона; или
«(2), который выбирает назначение в соответствии со статьей 6 настоящего Закона;
исчисляется со дня назначения такого судьи судьей предыдущего пятого округа.
«Разд. 8.Одиннадцатый округ уполномочен проводить заседания или заседания суда в Новом Орлеане, штат Луизиана, до тех пор, пока в Атланте, штат Джорджия, не будут созданы соответствующие помещения для такого суда.
«Разд. 9. Положения следующих параграфов данного раздела применяются к любому случаю, в котором за день до даты вступления в силу настоящего Закона [Oct. 1, 1981], апелляция или иное производство было подано в бывший пятый округ: «(1) Если вопрос был передан на решение, дальнейшее разбирательство по нему должно проводиться таким же образом и с такой же силой, как если бы этот Закон [вносящий поправки в разделы 41, 44 и 48 настоящего раздела и вводящие в действие положения, изложенные как примечания к этому разделу] не был принят.«(2)Если дело не было передано на рассмотрение, апелляция или судебное разбирательство вместе с оригиналами документов, распечатанными записями и должным образом заверенными записями должны быть переданы соответствующими приказами в суд, в который они бы обратились, если бы это было Закон был в полной силе и действовал на момент подачи такой апелляции или начала другого производства, и дальнейшее производство по делу должно проводиться таким же образом и с той же силой, как если бы апелляция или другое производство было подано в такой суд.
«(3)Ходатайство о повторном слушании или ходатайство о повторном слушании в банке по делу, решенному до даты вступления в силу настоящего Закона [Октябрь. 1, 1981], или представленные до даты вступления в силу настоящего Закона и решение о дате вступления в силу или после нее, как предусмотрено в пункте (1) настоящего раздела, должны рассматриваться таким же образом и с той же силой, как если бы этот Закон имел не был принят. Если ходатайство о повторном слушании в полном объеме удовлетворено, дело должно быть рассмотрено судом, как если бы настоящий Закон не был принят.
«Разд. 10. В статьях 5, 6, 7, 8 и 9 настоящего Закона термин — «(1)«бывший пятый округ» означает пятый судебный округ Соединенных Штатов, существовавший за день до даты вступления в силу настоящего Закона [октябрь. 1, 1981];
«(2)термин «новый пятый округ» означает пятый судебный округ Соединенных Штатов, учрежденный поправкой, внесенной в соответствии с разделом 2 (2) настоящего Закона [поправка, касающаяся пятого округа в этом разделе]; и
«(3)термин «одиннадцатый округ» означает одиннадцатый судебный округ Соединенных Штатов, учрежденный поправкой, внесенной в соответствии с разделом 2 (3) настоящего Закона [добавление пункта, относящегося к одиннадцатому округу в этом разделе].”
Административные действия Апелляционного суда Пятого округа; Прекращение судопроизводстваПаб. L. 96–452, §11, 14 октября 1980 г., 94 Stat. 1996 г., при условии, что:
«Апелляционный суд пятого округа, учрежденный за день до даты вступления в силу настоящего Закона [окт. 1, 1981] может принимать такие административные меры, которые могут потребоваться для выполнения настоящего Закона [внесение поправок в разделы 41, 44 и 48 настоящего раздела и принятие положений, изложенных в примечаниях к этому разделу]. Такой суд прекращает свое существование в административных целях с 1 июля 1984 г.” Административные подразделения апелляционного судаПаб. L. 95–486, §6, 20 октября 1978 г., 92 Stat. 1633, при условии, что:
«Любой апелляционный суд, имеющий более 15 действующих судей, может образовывать административные единицы, укомплектованные такими помещениями и персоналом, которые могут быть предписаны Административным управлением судов Соединенных Штатов, и может выполнять свои функции в полном объеме. функционирует таким количеством членов своих судов en banc, которое может быть предписано постановлением апелляционного суда ».
Комиссия по ревизии Федеральной апелляционной системыПаб.L. 92–489, 13 октября 1972 г., 86 Stat. 807, с изменениями, внесенными Pub. L. 93–420, 19 сентября 1974 г., стр. 88 Stat. 1153, предусмотренных на создание, членство, командировочные расходы, персонал, экспертов и консультантов, административные и исследовательские услуги, сотрудничество с другими правительственными учреждениями, а также ассигнования в размере не более 606 000 долларов США Комиссии по пересмотру апелляционной системы Федерального суда, которая была изучить географическое разделение судебных округов, а также структуру и внутренние процедуры апелляционной судебной системы и сообщить президенту, Конгрессу и главному судье свои рекомендации по изменению географических границ округов для ускорения решения судебных дел и внесение изменений в структуру апелляционных судов для ускорения рассмотрения дел в апелляционных судах в соответствии с фундаментальными концепциями справедливости и надлежащей правовой процедуры.Комиссия должна была прекратить свое существование через девяносто дней после представления своего окончательного отчета, который был представлен 20 июня 1975 года.
Продолжение организации судаЗакон от 25 июня 1948 г., гл. 646, §2 (b), 62 Стат. 985, частично при условии, что положения этого раздела, изложенные в разделе 1 закона от 25 июня 1948 г., в отношении организации каждого из нескольких судов, предусмотренных в нем, должны быть истолкованы как продолжение существующего закона, а срок полномочий судей, офицеров и служащих, а также поверенных и маршалов Соединенных Штатов, их заместителей и помощников, занимающих свои должности с сентября1, 1948, не будут затронуты его вступлением в силу, но каждый из них должен продолжать исполнять свои обязанности в том же качестве согласно соответствующим положениям настоящего раздела в соответствии с его предыдущим назначением.
Эффективная передача сигналов IL-2R по-разному влияет на стабильность, функцию и состав пула регуляторных Т-клеток
Характеристика Treg-клеток с гипоморфной мутацией в гене
Cd25Во время рутинного анализа проточной цитометрии мы наблюдали снижение CD25 экспрессия в клетках Treg и определенных субпопуляциях тимоцитов, а также in vitro активированных CD4 + Т-клеток из одного штамма с дефицитом Ccr2 ( Ccr2 tm1Ifc ), но не из другого штамма с дефицитом Ccr2- Ccr2 tm1Mae ) (рис.1а и дополнительный рис. 1). Мы выполнили полногеномное секвенирование ДНК, чтобы определить, может ли спонтанная мутация у мышей Ccr2 tm1Ifc быть причиной низкой экспрессии CD25. Из всех найденных вариантов последовательности мы идентифицировали один, влияющий на ген Cd25 (дополнительная таблица 1). Идентифицированный вариант нес точечную мутацию (T – C) в положении Chr2: 11680290 в экзоне 4, что привело к замене тирозина (Y) на гистидин (H) в кодоне 129 (Y129H) (рис.1б). Исходя из известной трехмерной структуры человеческого CD25, 27 Y129 расположен во внеклеточной части белка и, вероятно, образует водородную связь с P8, которая может помочь стабилизировать область белка CD25, которая находится в прямом контакте с Ил-2 (дополнительный рис. 2а). Наличие мутации Y129H было независимо подтверждено секвенированием по Сэнгеру фрагмента геномной ДНК у мышей Ccr2 tm1fFc (данные не показаны). Более того, чтобы избежать смешанных эффектов, мутировавшие аллели Ccr2 были заменены путем обратного скрещивания с мышами B6 в течение по меньшей мере трех поколений, что привело к поколению мышей Cd25 wt / Y129H .Скрещивание этих гетерозиготных мутантов привело к получению гомозиготных мутантных мышей ( Cd25 Y129H ), которых использовали во всех экспериментах вместе с их однопометниками дикого типа в качестве контроля. Гомозиготные мыши , Cd25, , , Y129H, были фертильны и выживали до зрелого возраста без явных дефектов развития (данные не показаны). Чтобы лучше понять молекулярный механизм, с помощью которого мутация Cd25 -Y129H приводит к снижению экспрессии CD25, мы проанализировали уровни белка и мРНК CD25 в клетках Foxp3 + Treg от мышей WT и Cd25 Y129H .Средняя интенсивность флуоресценции (MFI) как клеточной поверхности, так и внутриклеточного CD25 была сильно снижена у мышей Cd25 Y129H (фиг. 1c). Кроме того, мы обнаружили аналогичные результаты в активированных in vitro Т-клетках CD4 + мышей WT и Cd25 Y129H и в клетках 293T, временно трансфицированных плазмидами, кодирующими ген CD25-WT или CD25-Y129H (дополнительный рисунок 3a). , б). Этот результат был подтвержден конфокальной иммунофлуоресцентной микроскопией, которая показала, что подавление CD25 было особенно заметным на уровне клеточной поверхности, тогда как уровни внутриклеточной экспрессии CD25 были более устойчивыми (рис.1г). Более того, количественная оценка уровней транскрипта Cd25 с помощью ПЦР в реальном времени показала, что клетки WT и Cd25 Y129H Treg экспрессировали сравнимые количества мРНК Cd25 , что дает возможность того, что мутация Cd25 -Y129H влияет на транскрипцию мРНК или стабильность маловероятна (рис. 1д). Однако после стимуляции in vitro Т-клеток CD4 + с помощью mAb против CD3 и против CD28 мы обнаружили снижение уровней транскрипта Cd25 в клетках Cd25 Y129H по сравнению с клетками WT, что позволяет предположить, что транскрипционная активность может быть затронуто в мутантных Cd25 Y129H Т-клетках после активации через TCR (дополнительный рис.3в). Альтернативно, снижение экспрессии CD25 может ограничивать петлю положительной обратной связи IL-2 / CD25, что приводит к снижению синтеза мРНК. Более того, мы также показали, что белок CD25 не накапливается внутриклеточно, за исключением гипотетического дефекта транспорта белка к клеточной мембране в клетках Cd25 Y129H . Независимо от основного течения, наши данные показали, что мутация Cd25 Y129H привела к снижению уровня CD25.
Фиг.1Идентификация и функциональная характеристика гипоморфной мутации Cd25 Y129H . a FACS-графики CD3 + CD4 + лимфатических узлов дикого типа (слева), Ccr2 tm1Ifc (в центре) и Ccr2 9 tm после окрашивания антителами к CD25 и Foxp3. Цифры внутри точечных графиков указывают процент ячеек в пределах стробируемой области.Показаны репрезентативные точечные графики из двух независимых экспериментов с 3 мышами на каждый генотип. b Нуклеотидные и кодируемые аминокислотные последовательности локуса Cd25 у мышей WT и Cd25 Y129H . c Экспрессия поверхностного и внутриклеточного CD25 с помощью Foxp3 + Treg-клеток от мышей WT и Cd25 Y129H проанализирована с помощью проточной цитометрии. Числа на графиках указывают среднюю интенсивность флуоресценции (MFI) CD25 и суммированы на гистограмме (справа).Данные были получены из двух независимых экспериментов с 2 мышами на каждый генотип. d Изображения конфокальной микроскопии WT и Cd25 Y129H Treg-клеток; поверхность CD25 — зеленый; внутриклеточный CD25 — красный; DAPI — синий. Изображения представляют два независимых эксперимента; масштабная линейка 5 мкм. e Анализ qRT-PCR экспрессии мРНК Cd25 в клетках Foxp3 + Treg от мышей WT и Cd25 Y129H . Данные получены из четырех независимых экспериментов с 4 мышами на каждый анализируемый генотип. f Внутриклеточный фосфорилированный тирозином STAT5 (pSTAT5) у WT (белые кружки) и Cd25 Y129H (темные кружки) Foxp3 + Treg-клетки, стимулированные in vitro в течение 20 минут с различными количествами IL-2. Данные были получены из трех независимых экспериментов. г Цитометрический анализ IL-2 в среде через 3 часа после культивирования без (серая полоса) или с Foxp3 + Tregs от мышей WT (белая полоса) или Cd25 Y129H (черная полоса).Данные были получены из трех независимых экспериментов. ч Сравнение способности WT (светлые кружки) и Cd25 Y129H (темные кружки) Foxp3 + Treg-клетки подавлять CD4 + пролиферацию Т-клеток in vitro. Серийные номера Treg-клеток культивировали с меченными ef670 общими CD4 + Т-клетками в течение 3 дней с шариками, покрытыми анти-CD3 и анти-CD28 антителами. Пролиферацию Т-клеток CD4 + определяли с помощью проточной цитометрии, и результаты рассчитывали как процент подавления.Данные были получены из двух независимых экспериментов и представлены как среднее ± стандартное отклонение; нс не имеет значения; * p <0,05, ** p <0,01, *** p <0,001; двусторонний непарный тест Стьюдента t
Чтобы продемонстрировать функциональные последствия снижения экспрессии CD25 в Treg-клетках Cd25 Y129H , мы оценили индуцированное IL-2 фосфорилирование сигнального преобразователя и активатора транскрипции 5 (pSTAT5 ), что отражает основной результат передачи сигналов IL-2R.WT и Cd25 Y129H Treg-клетки проявляли разные дозовые реакции на рекомбинантный hIL-2 (фиг. 1f). При низкой концентрации ИЛ-2 (0,32 Ед / мл) величина ответа (измеренная как% клеток pSTAT5 + ) была значительно выше (в четыре раза) в клетках Treg дикого типа, чем в клетках Treg Cd25 Y129H . , тогда как при высокой концентрации IL-2 (8 Ед / мл) фракция клеток pSTAT5 + была сопоставима между клетками WT и Cd25 Y129H Treg.Эти результаты показывают, что реактивность Treg-клеток Cd25 Y129H к физиологическим количествам IL-2, которые обычно не превышают 0,05 Ед / мл, снижена. 28 Затем мы спросили, могут ли Treg-клетки Cd25 Y129H поглощать и истощать экзогенный IL-2, что, как предполагалось, является важным механизмом опосредованного Treg-клетками подавления. 11,29,30 Используя анализ in vitro, мы обнаружили, что потребление IL-2 Treg-клетками Cd25 Y129H было значительно снижено по сравнению с потреблением Treg-клетками WT (рис.1г). Это открытие предполагает, что in vivo низкая экспрессия CD25 Cd25 Y129H Treg-клетками может снизить их способность конкурировать за IL-2, продуцируемый активированными Т-клетками, что, в свою очередь, потенциально может привести к снижению способности подавлять пролиферацию обычные Т-клетки. Более того, сравнивая in vitro супрессивную активность Treg-клеток WT и Cd25 Y129H , оценивая их способность ингибировать пролиферацию CD4 + Т-клеток при различных соотношениях клеток, мы обнаружили, что подавляющая способность Cd25 Y129H Treg клеток сильно уменьшился (рис.1ч). В целом, эти результаты демонстрируют, что снижение экспрессии CD25 в Treg-клетках Cd25 Y129H значительно снижает их чувствительность к IL-2 и их иммуносупрессивную способность.
Treg клеток в
Cd25 Мыши Y129H демонстрируют измененный гомеостаз и фенотипИсследование тимоцитов мышей WT и Cd25 Y129H выявило аналогичные процентные и абсолютные числа клеток CD4 — CD8 — двойные отрицательные 9041 + CD4 + двойных положительных и CD4 + и CD8 + одноположительных тимоцитов, что позволяет предположить, что мутация Cd25 Y129H не влияет на развитие тимоцитов (дополнительный рис.4а). Аналогичным образом, процентное и абсолютное количество CD4 + Т-клеток, CD8 + Т-клеток и CD44 hi CD4 + и CD44 hi CD8 + эффекторных Т-клеток / Т-клеток памяти также были в значительной степени сопоставимы между WT и Cd25 Y129H мышей в периферических лимфоидных органах (лимфатические узлы и селезенка), что указывает на то, что мутация Cd25 Y129H существенно не влияет на активацию периферических Т-клеток (дополнительный рис.4б – д). Затем мы исследовали компартмент Treg-клеток в лимфоидных органах и нелимфоидных тканях (NLT), оценивая экспрессию Foxp3 (рис. 2a). В целом процентное и абсолютное количество клеток Treg было значительно снижено у мышей Cd25 Y129H по сравнению с их однопометниками дикого типа (рис. 2b, c). По сравнению с таковым у мышей WT, процент Treg-клеток среди CD4 + Т-клеток у мышей Cd25 Y129H был снижен на ~ 50% в pLN, селезенке и mLN, но был менее выражен в NLT.В соответствии с меньшими процентными значениями, абсолютное количество Treg-клеток также было значительно снижено у мышей Cd25 Y129H . Интересно, что эта тенденция была менее выражена в тимусе, где процентное содержание и абсолютное количество клеток Treg среди тимоцитов CD4SP было лишь немного ниже у мышей Cd25 Y129H , чем у мышей WT (рис. 2b, c). Как и ожидалось, экспрессия CD25 в Treg-клетках была сильно снижена у мышей Cd25 Y129H (рис.2г). Примечательно, что Cd25 Y129H Treg клетки обнаруживают умеренное, но значительное снижение экспрессии Foxp3, на что указывает уменьшение MFI Foxp3-GFP (фиг. 2e).
Рис. 2Гипоморфная мутация Cd25 Y129H приводит к снижению экспрессии CD25 и потере Foxp3 + Treg-клеток. a Типичные графики FACS, показывающие экспрессию Foxp3 и CD25 Т-клетками CD4 + в периферических лимфатических узлах (pLN), селезенке, тимусе, брыжеечных лимфатических узлах (mLN), печени и легких в течение 8–12 недель. -старых мышей WT и Cd25 Y129H . b Процентные доли и c абсолютное количество клеток Foxp3 + Treg-клеток в различных органах мышей WT (закрашенные кружки) и Cd25 Y129H (белые кружки). Средние интенсивности флуоресценции (MFI) CD25 ( d ) и Foxp3 ( e ) на Treg-клетках мышей WT и Cd25 Y129H . Данные были получены как минимум из трех экспериментов с 5–8 мышами на каждый генотип и представлены в виде среднего значения ± стандартное отклонение. нс не имеет значения; * p <0.05, ** p <0,01, *** p <0,001, **** p <0,0001; двусторонний непарный тест Стьюдента t
Затем мы сравнили экспрессию молекул адгезии и хоминговых рецепторов между WT и Cd25 Y129H Treg-клетками из разных лимфоидных тканей и NLT. По сравнению с Treg-клетками WT, Cd25 Y129H Treg-клетки показали повышенную экспрессию хемокинового рецептора CXCR3 и нескольких интегринов и подавленную экспрессию хемокинового рецептора CCR7 лимфоидного хоминга (дополнительный рис.5а). Напротив, в клетках, не являющихся Treg, эти молекулы экспрессировались на одинаковых уровнях в обеих линиях мышей (дополнительный рис. 5b). Этот результат предполагает, что в Treg-клетках передача сигналов IL-2R может придавать фенотип, позволяющий им жить в лимфоидных тканях, а не в NLT. Несмотря на наличие «эффекторного» фенотипа, Cd25 Y129H Treg-клетки были распределены аналогично их WT-аналогам в различных лимфоидных органах, как показывает гистология (дополнительный рис. 5c).Таким образом, наши результаты предполагают, что эффективная передача сигналов IL-2R необходима для полного гомеостаза Treg-клеток и поддержания фенотипического разнообразия.
Усиленный активированный / эффекторный фенотип
Cd25 Y129H Treg-клеткиСообщалось, что в отсутствие передачи сигналов IL-2R Treg-клетки проявляют активированный фенотип. 4 Поэтому мы измерили уровни ключевых молекул сигнатур Treg-клеток у мышей WT и Cd25 Y129H с помощью проточной цитометрии. Cd25 Y129H Клетки Treg экспрессировали более высокие уровни маркеров, связанных с активацией клеток, включая CD103 (рис. 3a), GITR (рис. 3c) и ICOS (рис. 3d), в то время как экспрессия маркеров, связанных с Treg-клетками супрессивная функция, включая CTLA-4 (фиг. 3b) и PD-1 (фиг. 3e), существенно не отличалась от таковой у Treg-клеток дикого типа. Кроме того, снижение экспрессии CD25 в Cd25 Y129H Treg-клетках сопровождалось повышенной экспрессией CD127 (IL-7Rα) (рис.3е). Сходные результаты были обнаружены как в клетках CD62L + , CD44 — Treg, так и в клетках CD62L — CD44 + Treg (данные не показаны). Однако у мышей Cd25, , , Y129H было снижено процентное и абсолютное количество клеток CD62L + CD44 — Treg-клеток в лимфоидных тканях и NLT (рис. 3g). Напротив, абсолютное количество клеток CD62L — CD44 + Treg-клеток было сопоставимо у мышей WT и Cd25 Y129H , хотя повышенное процентное содержание CD62L — CD44 + Treg-клеток наблюдалось у Cd25 . Y129H мышей (рис.3ч). В целом, эти результаты показали, что сниженная экспрессия CD25 специфически нарушает поддержание CD62L + CD44 — Treg-клеток, независимо от того, находятся ли они в лимфоидных тканях или NLT, что согласуется с предыдущими сообщениями, показывающими, что IL-2R передача сигналов особенно необходима для гомеостаза клеток cTreg. 31,32
Рис. 3Активированный / эффекторный фенотип Foxp3 + Treg-клеток у мышей Cd25 Y129H . a — f Репрезентативные гистограммы (верхние панели) и количественный анализ (нижние панели) MFI указанных поверхностных маркеров Treg на CD4 селезенки + Foxp3 + Treg-клетки от WT (закрашенные кружки) и Cd25 Y129H (светлые кружки) мыши. Данные были получены как минимум из трех независимых экспериментов с 5 мышами на каждый генотип. Процентное соотношение (слева) и абсолютное количество клеток (справа) CD62L + ( г ) и CD44 + ( h ) популяций Treg-клеток в разных органах мышей WT и Cd25 Y129H .Данные были получены как минимум из трех независимых экспериментов с 6-8 мышами на каждый генотип и представлены как среднее значение ± стандартное отклонение; нс не имеет значения; * p <0,05, ** p <0,01, *** p <0,001; двусторонний непарный тест Стьюдента t
Отсутствие спонтанного аутоиммунитета у
Cd25 Y129H мышиЗатем мы исследовали, проявляют ли мыши Cd25 Y129H признаки аутоиммунитета, как ранее сообщалось для мышей, у которых отсутствует экспрессия различных компонентов передачи сигналов IL-2R. 4,11,12,13,14 По сравнению с мышами WT, мыши Cd25 Y129H показали аналогичный прирост массы тела с возрастом (рис. 4a), не имели существенного увеличения CD4 + или CD8 + Т-клеток и показали сопоставимые процентные и абсолютные количества клеток, демонстрирующих фенотип памяти CD44 hi (рис. 4b). Кроме того, гистопатологический анализ старых мышей (35-40 недель) обеих линий выявил сопоставимую инфильтрацию лимфоцитов во многих органах, таких как слюнная железа, слезные железы и поджелудочная железа (рис.4в, г). Таким образом, отсутствие аутоиммунитета у мышей Cd25, , Y129H , вероятно, указывает на то, что Treg-клетки, развивающиеся в этом состоянии нарушенной передачи сигналов IL-2R, все еще функциональны и сохраняют свою супрессивную способность во время старения. С другой стороны, отсутствие аутоиммунитета у мышей Cd25, , Y129H может также отражать потерю функции IL-2R в аутореактивных Т-клетках CD4 + и CD8 + . Чтобы изучить эту возможность, обычные Т-клетки CD4 + от этих мышей стимулировали антителами против CD3 / CD28 и подвергали дифференцировке Т-хелперных клеток in vitro.По сравнению с клетками WT, Т-клетки от мышей Cd25, , Y129H показали значительно более низкую внутриклеточную экспрессию цитокинов Th2 (IFN-γ), аналогичную экспрессию цитокинов Th3 (IL-4) и умеренно повышенную экспрессию цитокинов Th27 (IL-17) ( Дополнительный рис.6). Эти данные предполагают, что передача сигналов IL-2R необходима для максимальной генерации IFN-γ-продуцирующих клеток во время иммунного ответа типа 1. Более того, аномальная реакция потенциально аутореактивных Т-клеток может частично способствовать отсутствию спонтанного аутоиммунитета у мышей Cd25 Y129H .
Рис. 4Отсутствие аутоиммунных симптомов у старых мышей Cd25 Y129H . a Еженедельный мониторинг массы тела самок (слева) и самцов (справа) мышей WT (черная линия) и Cd25 Y129H (красная линия). Данные были получены от 6 до 12 мышей по генотипу и полу. b Процентное и абсолютное количество клеток CD44 hi среди CD4 + и CD8 + Т-клеток из pLN (слева) и селезенки (справа) WT (закрашенные кружки) и Cd25 Y129H (светлые кружки) мыши.Данные были получены как минимум из трех независимых экспериментов с 9-13 мышами на каждый генотип. c Репрезентативная иммунофлуоресцентная микроскопия слюнных желез, слезных желез и поджелудочной железы мышей WT (вверху) и Cd25 Y129H (внизу), окрашенных указанными антителами. Масштабная линейка 500 мкм. d Индивидуальный размер (слева) и количество (справа) лимфоидных инфильтратов на разрез в слюнных железах, слезных железах и поджелудочной железе мышей WT (закрашенные кружки) и Cd25 Y129H (светлые кружки).Данные были получены как минимум из трех независимых экспериментов с 9-13 мышами на каждый генотип. a — d Органы выделяли от мышей в возрасте от 35 до 40 недель. Данные представлены в виде среднего значения ± стандартное отклонение. нс не имеет значения; * p <0,05 (двусторонний непарный тест Стьюдента t )
Передача сигналов IL-2R уравновешивает ландшафт подмножеств Treg-клеток
Чтобы понять, как передача сигналов IL-2R формирует гетерогенность различных подмножеств Treg-клеток на молекулярном уровне. На уровне скРНК CD4 + Foxp3 + Treg-клеток селезенки мы выполнили секвенирование мышей WT и Cd25 Y129H (рис.5а). Неконтролируемая графическая кластеризация организовала клетки Treg в четыре кластера и идентифицировала транскрипты, которые лучше всего характеризовали различные группы (рис. 5b; дополнительная таблица 3). Чтобы связать каждый кластер с известными подмножествами Treg-клеток, мы скринировали 60 наиболее значимо активированных генов каждого кластера и перекрыли их с ранее сообщенными маркерами Treg-клеток и каноническими сигнальными путями (рис. 5c; дополнительная таблица 2). Гены кластера 1 с высокой степенью экспрессии, такие как Ifit1, Ifit3, Isg15 и Stat1 , связанные с ответами интерферона I типа (IFN), но также гены, которые в первую очередь экспрессируются наивными клетками (например.g., Sell и Ly6c1) , которые, скорее всего, отражают промежуточное состояние активации клеток или прямой ответ на IFN. Мы назвали эту популяцию Stat-1 + Treg-клетками, как было предложено ранее для Treg-клеток, выделенных из плечевых LN. 33 Кластер 2 включал гены, экспрессируемые в клетках cTreg, 31,32 , такие как , как Sell, Ccr7, Bcl2, Ly6c1 и S1pr1 . Treg-клетки кластера 1 и 2 также показали повышенную экспрессию Ms4a4b и Ms4a6b, что может способствовать состоянию покоя этих субпопуляций Treg-клеток, ингибируя прогрессирование клеточного цикла. 34 Кластер 3 был обогащен генами, которые были высоко экспрессированы в эффекторных клетках Treg (eTreg). 35,36 Этот кластер включал ключевые гены сигнатуры Treg-клеток, такие как Ctla4, Tnfrsf9 (4-1BB) и Izumo1r , а также рецепторы клеточной поверхности, такие как Tnfrsf4 (Ox40), Tnfrsf18 (GITR), Pdcd1 (PD-1), Icos и Nrp1 . Кроме того, экспрессия многих факторов транскрипции, которые контролируют дифференцировку и функцию клеток eTreg, 37 , таких как Batf, Ikzf2 (Helios), Maf и Gata3 , была усилена в кластере 3, тогда как экспрессия из Klf2, Satb1 , Bach3 и Lef1 , которые противодействуют дифференцировке клеток eTreg, 38,39,40,41 было увеличено в кластере 2 (рис.5г). Кроме того, кластер 3 также сверхэкспрессирует гены, связанные с ответом на сигналы TCR, такие как Rel, Orai1 и Zap70 , что отражает недавнее вовлечение TCR. Некоторые из маркеров клеточной поверхности, которые характерны для активированных Treg-клеток, такие как Maf, Nrp1 и Icos , также были высоко экспрессированы в кластере 4, что согласуется с идеей, что эти кластеры идентифицируют субпопуляции Treg-клеток, которые представляют собой континуум дискретных состояний, а не отдельных сущностей. 33 Этот кластер также включает факторы транскрипции, которые способствуют дифференцировке или функции резидентных в ткани Treg-клеток, 37 , таких как , как Rora, Runx3, Fli1 и Id2 , а также гены, функционально связанные с тканью. хоминг и трафик, включая Itgb1 , Itgb7 , Cxcr3 и Ccr2 , что указывает на то, что кластер 4, вероятно, соответствует клеткам, претерпевающим более позднюю стадию созревания и предварительной адаптации к их транспортировке на NLT.Мы назвали эту популяцию NLT-подобными Treg-клетками, как описано ранее. 33
Рис. 5Идентификация субпопуляций Treg-клеток с помощью одноклеточного анализа РНК-seq. a Экспериментальный рабочий процесс для анализа scRNA-seq, включая сортировку Treg-клеток селезенки Foxp3 GFP Treg мышей WT или Cd25 Y129H и использование набора реагентов Chromium Single Cell 3 ‘. b Кластеризация на основе UMAP объединенных профилей scRNA-seq WT и Cd25 Y129H Treg-клеток.Тепловая карта, показывающая экспрессию выбранных маркерных генов ( c ) и выбранных факторов транскрипции ( d ) для каждого из четырех кластеров, показанных в b . Полный список генов и значений экспрессии доступен в дополнительной таблице 3
Затем мы сравнили долю каждого подмножества Treg-клеток у мышей WT и Cd25 Y129H . В то время как подмножество клеток cTreg было значительно сокращено, подмножество клеток eTreg было высоко обогащено у мышей Cd25 Y129H (рис.6а, б). Подмножества Stat-1 + и NLT-подобных Treg-клеток имели одинаковую численность как у мышей WT, так и у мышей Cd25 Y129H . Эти данные согласуются с данными проточной цитометрии, показывающими, что Cd25 Y129H Treg клетки содержат более высокую фракцию CD44 + CD62L — Treg (фиг. 3h). Кроме того, анализ дифференциальной экспрессии показал, что более 40% DEG в клетках WT и Cd25 Y129H Treg в значительной степени перекрываются среди кластеров 2, 3 и 4, что указывает на то, что передача сигналов IL-2R важна для поддержания программ транскрипции эти разные подмножества Treg-клеток (рис.6c; Дополнительная таблица 4). Это было в отличие от клеток кластера 1, которые демонстрировали очень мало DEG между WT и Cd25 Y129H Treg-клетками и имели уменьшенное перекрытие с другими кластерами, предполагая, что гомеостаз этого кластера в значительной степени не зависит от IL-2R (дополнительный рис. . 7а). Однако, чтобы прояснить этот момент, необходимо определить более качественные поверхностные маркеры. Общее сравнение профилей экспрессии генов селезенки WT и Cd25 Y129H Treg клеток показало, что 64 гена были активированы, а 29 генов подавлены в Treg клетках WT по сравнению с Cd25 Y129H Treg-клетками (рис.6d; Дополнительная таблица 5). Клетки WT Treg экспрессировали более высокие уровни генов, необходимых для поддержания Т-клеток, таких как антиапоптотические молекулы Mcl-1 и Bclb11 (дополнительный рис. 7b), что согласуется с ролью передачи сигналов IL-2R в выживании Т-клеток. . 42,43 Напротив, Cd25 Y129H Treg-клетки показали более высокую экспрессию CD127, что указывает на то, что они могут использовать передачу сигналов IL-7 для выживания, чтобы компенсировать сниженную передачу сигналов IL-2R.Кроме того, наиболее значимо активированные гены в Treg-клетках WT включают многие ДНК-связывающие регуляторы транскрипции, такие как Klf2, Cited2, Pura, Crebzf, Jund , и некоторые репрессоры транскрипции, включая Aes (также известный как Tle5), Ncor1. и Suds3 , подтверждая, что передача сигналов IL-2R регулирует транскрипцию генов Treg-клеток. 44 Затем анализ обогащения на основе GO показал, что биологические процессы, связанные с метаболизмом, биосинтезом и экспрессией генов, были значительно больше представлены в Treg-клетках дикого типа (дополнительный рис.7в). Напротив, гены, активируемые в клетках Cd25 Y129H Treg, были связаны с развитием и функцией клеток eTreg, включая Ikzf2, Maf, Itgb1, Gata3, S100a6, S100a10 и S100a11 , как предполагалось ранее (Рис. 6d). Интересно, что этот список генов сильно перекрывается с генами, связанными с кластером 4 (дополнительная таблица 3), подтверждая идею о том, что передача сигналов IL-2R уравновешивает гетерогенность клеток Treg, и что снижение передачи сигналов IL-2R смещает баланс в сторону эффекторного фенотипа.Для дальнейшего изучения этой гипотезы мы оценили кластеры Treg-клеток в отношении их развития с помощью программного пакета Slingshot. 26 Мы обнаружили, что предполагаемый спектр дифференцировки частично воспроизводит кластеризацию клеток, упорядочивая cTregs и eTregs, за которыми следуют NLT-подобные Tregs на линейной траектории развития (Fig. 6e). Используя тот же подход, мы также наблюдали, что Cd25 Y129H cTreg и eTreg клетки имели псевдовремя развития, наклоненное в сторону более эффекторного фенотипа, что указывает на более быстрое приобретение эффекторных свойств при нарушенной передаче сигналов IL-2R (рис.6е). В соответствии с недавними исследованиями, наш анализ scRNA-seq выявил присутствие различных субпопуляций Treg-клеток в лимфоидных тканях в стабильном состоянии. Кроме того, сравнивая одноклеточные транскриптомы Treg-клеток, получающих оптимальную или низкую передачу сигналов IL-2R, мы идентифицировали генные программы, связанные с Treg-клетками, которые зависят от полной передачи сигналов IL-2. В целом эти данные демонстрируют, что нарушение передачи сигналов IL-2R не позволяет поддерживать гетерогенность Treg-клеток.
Фиг.6Сравнение WT и Cd25 Y129H Субпопуляции Treg-клеток по одноклеточной последовательности РНК. Кластеризация на основе UMAP ( a ) и количественный анализ ( b ) Treg-клеток селезенки мышей WT и Cd25 Y129H . Данные были получены от 3 мышей на группу и представлены как log 2 -кратных изменений (см. «Материалы и методы»). c Диаграмма Венна, показывающая перекрытие между кластерами 2, 3 и 4 для генов, дифференциально экспрессируемых между WT и Cd25 Y129H Treg-клетками. d Тепловая карта, показывающая 20 лучших дифференциально экспрессируемых генов в клетках WT и Cd25 Y129H Treg. Полный список генов и значений экспрессии показан в дополнительной таблице 5. e , f Анализ вывода траектории, выполненный с помощью программного обеспечения Slingshot
Уменьшение оральной толерантности у
Cd25 Y129H мышиБыло показано, что локальная экспансия Treg-клеток в собственной пластинке кишечника имеет важное значение для установления толерантности к пищевым антигенам. 46 Таким образом, мы использовали Cd25 Y129H мышей для изучения роли передачи сигналов IL-2R в индукции оральной толерантности в модели DTH. С этой целью мыши WT и Cd25 Y129H получали либо две дозы 25 мг OVA, либо PBS через желудочный зонд (фиг. 7a). Через неделю мышей сенсибилизировали подкожной иммунизацией OVA / CFA. Две недели спустя индукцию толерантности оценивали путем измерения реакций DTH на внутрикожную инъекцию OVA. Мыши WT, получавшие OVA, показали сниженные ответы DTH по сравнению с мышами, получавшими PBS, что указывает на индукцию оральной толерантности (рис.7б). Напротив, в тех же условиях мыши Cd25, , Y129H не смогли развить оральную толерантность, о чем свидетельствует нарушение способности подавлять ответы DTH, что свидетельствует о дефектной функции Treg-клеток у мышей Cd25, , Y129H .
Рис. 7Cd25 Y129H Мыши не смогли развить оральную толерантность. a Блок-схема протокола индукции допуска. Мышей b WT (кружки) и Cd25 Y129H (квадраты) дважды кормили 25 мг овальбумина в PBS (OVA; серые столбцы) или только PBS (белые столбцы).На 7 день всех мышей иммунизировали подкожной инъекцией OVA, эмульгированной в CFA, и через 2 недели заражали подкожной инъекцией OVA в PBS в ухо. Индукцию толерантности анализировали путем измерения отека уха через 48 ч после заражения (см. «Материалы и методы»). Данные были получены как минимум из трех независимых экспериментов с 6-8 мышами в группе и представлены как среднее значение ± стандартное отклонение. нс не имеет значения; * p <0,05, *** p <0,001 (двусторонний непарный тест Стьюдента t )
Cd25 Y129H Treg-клетки обладают пониженной супрессорной способностью in vivoЗатем мы исследовали способность Cd25 Y129H Treg-клеток подавлять воспалительные реакции кишечника in vivo.В модели адоптивного колита с переносом Т-клеток доставка наивных Т-клеток CD4 + реципиентам с иммунодефицитом приводит к тяжелому воспалению кишечника, которое можно предотвратить с помощью котрансфера Treg-клеток. 47,48 Адоптивный перенос эффекторного CD25 — CD62L + CD4 + WT Т-клетки (наивные CD4 + ) реципиентам Rag2 — / — привел к развитию колита с обширной инфильтрацией воспалительного клеток и значительной индукции воспалительных цитокинов в ткани толстой кишки (рис.8а – г). Примечательно, что котрансфер наивных WT CD4 + T-клеток и Cd25 Y129H Treg-клеток реципиентам Rag2 — / — не предотвращал развитие колита так же эффективно, как котрансфер Treg-клеток WT. Гистологический анализ образцов толстой кишки от мышей-реципиентов, коинфицированных наивными WT CD4 + Т-клетками и Cd25 Y129H Treg-клетками, показал значительно увеличенные признаки патологического воспаления (рис. 8a, b) и повышенную продукцию воспалительных цитокинов по сравнению с реципиентом. мышам, коинъектированным совместно с наивными T-клетками WT CD4 + и Treg-клетками WT (рис.8г).
Фиг. 8Treg-клетки от доноров Cd25 Y129H демонстрируют пониженную супрессивную функцию in vivo и стабильность клонов в модели колита с переносом клеток. a Гистопатологическая оценка ткани толстой кишки мышей Rag2 — / — через 8 недель после внутрибрюшинного переноса наивных Т-клеток (CD25 — CD62L + CD4 + ) отдельно (белые кружки) или с WT (темные кружки ) или Cd25 Y129H (серые кружки) GFP + Foxp3 + Treg-клетки в соотношении 1: 1. b Репрезентативные срезы толстой кишки, окрашенные гематоксилином и эозином, из трех групп реципиентов. c Количество донорских эффекторных Т-клеток в мл количественно определяли с помощью FACS. d Экспрессию воспалительных цитокинов анализировали в общей ткани толстой кишки с помощью qRT-PCR. e Процент клеток Foxp3 + среди повторно выделенных Thy1.1 + CD4 + WT и Cd25 Y129H Treg-клеток через 8 недель после переноса в Rag2 — / — мышей-реципиентов. f Foxp3 + и Foxp3 — Спленоциты CD4 + были отсортированы от мышей WT и Cd25 Y129H , и геномная ДНК была проанализирована для определения статуса метилирования TSDR. Показан один репрезентативный анализ из двух независимых экспериментов. г Статус метилирования TSDR в повторно изолированном Thy1.1 + CD4 + WT и Cd25 Y129H Treg-клетки и наивные Thy1.2 + CD4 + T-клетки из мезентериальных клеток ) и селезенку через 8 недель после переноса в мышей-реципиентов Rag2 — / — .Процент метилирования обозначен цветом в соответствии с показанной шкалой. Данные были получены из четырех ( a — d ) и трех ( e , g ) независимых экспериментов и представлены как среднее значение ± стандартное отклонение. нс не имеет значения; * p <0,05, ** p <0,01, *** p <0,001. Односторонний дисперсионный анализ с поправкой Тьюки для множественных сравнений ( a , c ) или двусторонний непарный тест Стьюдента t ( d , e )
Примечательно, 30% от Cd25 Y129H Treg-клетки в mLN продемонстрировали сниженную экспрессию Foxp3 через 8 недель после индукции колита, тогда как Treg-клетки WT в значительной степени сохранили экспрессию Foxp3 (рис.8д). Это наблюдение предполагает, что нарушение передачи сигналов IL-2R клетками Treg значительно влияет на стабильность клонов и супрессивную способность во время воспаления. Деметилирование TSDR необходимо для стабильной экспрессии Foxp3 и, следовательно, для стабильного поддержания их супрессивной способности. По сравнению с Treg-клетками, выделенными ex vivo, Treg-клетки, индуцированные in vitro, демонстрируют фенотип неполного деметилирования, несмотря на высокую экспрессию Foxp3, что указывает на то, что характер деметилирования ДНК TSDR служит маркером долгосрочной стабильности и функции Treg-клеток. 49 Чтобы определить, является ли нестабильность адоптивно перенесенных Cd25 Y129H Treg-клеток из-за снижения деметилирования TSDR, мы сравнили статус метилирования TSDR в WT селезенки и Cd25 Y129H Treg-клетках. Однако пиросеквенирование обработанной бисульфитом геномной ДНК от сортированных Foxp3 + и Foxp3 — CD4 + Т-клеток выявило аналогичный деметилированный TSDR в Treg-клетках мышей WT и Cd25 Y129H (рис.8f). Учитывая роль TSDR в поддержании экспрессии Foxp3, эти результаты не были неожиданными, поскольку уровни белка Foxp3 практически не изменились в Treg-клетках Cd25 Y129H в стационарных условиях (рис. 2e). Более того, это наблюдение подтвердило, что в клетках Treg длительное снижение передачи сигналов IL-2R уменьшает размер пула клеток Treg, не влияя на стабильность клеток. Затем мы исследовали деметилирование TSDR в WT и Cd25 Y129H Treg клетках после переноса в Rag2 — / — мышей.В соответствии со сниженной супрессорной функцией Cd25 Y129H Treg-клеток, эти клетки показали тенденцию к снижению деметилирования TSDR, хотя это изменение не было статистически значимым (рис. 8g). Более того, это снижение коррелирует со снижением уровней Foxp3, что указывает на роль передачи сигналов IL-2R в стабильности Treg-клеток.
Казалось бы, неожиданное наблюдение заключалось в том, что у мышей Cd25, , Y129H не развивался спонтанный аутоиммунитет, тогда как в модели адоптивного переносного колита Treg-клетки этих мышей не могли предотвратить аутоиммунитет.Эти данные открывают возможность того, что Cd25 Y129H Treg-клетки способны в достаточной степени лишать IL-2 только Cd25 Y129H эффекторные клетки, а не эффекторные клетки WT, поскольку последние имеют более высокие уровни IL-2R. Чтобы проверить эту гипотезу, мы провели дополнительные эксперименты по колиту, в которых реципиентам Rag2 — / — были перенесены наивные Cd25 Y129H CD4 + Т-клетки отдельно или в комбинации с WT или Cd25 Y129H Treg-клетками. .В отличие от изложенной выше гипотезы, мы обнаружили, что перенос наивных Cd25 Y129H CD4 + Т-клеток приводил к заболеванию с серьезностью, аналогичной той, которая наблюдалась у мышей, перенесенных с наивными Т-клетками WT CD4 + (рис. 8 и дополнительный рисунок 8). В обеих ситуациях Cd25 Y129H Treg-клетки были неспособны контролировать воспаление у мышей-реципиентов. Это открытие указывает на то, что потеря супрессорной функции Treg-клеток Cd25 Y129H в модели колита может быть независимой от лишения IL-2, и указывает на внутреннюю роль передачи сигналов IL-2R в подавлении аутоиммунитета помимо лишения эффекторных клеток Ил-2.
JAC: Журнал теории композиции, том 20, номер 3, лето 2000 г.
Кто
Люди и организации, связанные либо с созданием этого журнала, либо с его содержанием.
Какие
Описательная информация, помогающая идентифицировать это периодическое издание. Перейдите по ссылкам ниже, чтобы найти похожие предметы в Электронной библиотеке.
Когда
Даты и периоды времени, связанные с этим периодическим изданием.
Статистика использования
Когда в последний раз использовалась эта проблема?
Взаимодействовать с этим периодическим изданием
Вот несколько советов, что делать дальше.
Ссылки, права, повторное использование
Международная структура взаимодействия изображений
Распечатать / Поделиться
Печать
Электронная почта
Твиттер
Facebook
Tumblr
Reddit
Ссылки для роботов
Полезные ссылки в машиночитаемом формате.
Ключ архивных ресурсов (ARK)
Международная структура взаимодействия изображений (IIIF)
Форматы метаданных
Изображений
URL
Статистика
Ассоциация преподавателей современного сочинения (У.С.). JAC: журнал теории композиции, том 20, номер 3, лето 2000 г., периодическое издание 2000; (https://digital.library.unt.edu/ark:/67531/metadc28630/: по состоянию на 3 июня 2021 г.), Библиотеки Университета Северного Техаса, Цифровая библиотека UNT, https://digital.library.unt.edu; .
Ингибирование передачи сигналов Hedgehog изменяет состав фибробластов при раке поджелудочной железы
Abstract
Цель: Протоковая аденокарцинома поджелудочной железы (PDAC) — смертельное заболевание, характеризующееся обширной фибровоспалительной стромой, которая включает многочисленные популяции фибробластов, ассоциированных с раком (CAF).CAF PDAC неоднородны, но природа этой неоднородности до конца не изучена. Путь Hedgehog функционирует в PDAC паракринным образом, при этом лиганды, секретируемые раковыми клетками, передают сигнал стромальным клеткам в микроокружении. Предыдущие сообщения об исследовании роли передачи сигналов Hedgehog в PDAC были противоречивыми, с альтернативным предложением передачи сигналов Hedgehog для стимулирования или ограничения роста опухоли. В свете недавно обнаруженной гетерогенности CAF мы исследовали, как ингибирование пути Hedgehog репрограммирует микросреду PDAC.
Дизайн эксперимента: Мы использовали комбинацию фармакологического ингибирования, генетических экспериментов с усилением и потерей функции, цитометрии по времени пролета и секвенирования одноклеточной РНК для изучения роли передачи сигналов Hedgehog в PDAC.
Результаты: Мы обнаружили, что передача сигналов Hedgehog однозначно активируется в фибробластах и дифференциально повышается в миофибробластных CAF (myCAF) по сравнению с воспалительными CAF (iCAF). Сверхэкспрессия Sonic Hedgehog способствует росту опухоли, в то время как ингибирование пути Hedgehog сглаженным антагонистом LDE225 замедляет рост опухоли.Кроме того, ингибирование пути Hedgehog снижает количество myCAF и увеличивает количество iCAF, что коррелирует с уменьшением цитотоксических Т-клеток и увеличением количества регуляторных Т-клеток, что согласуется с усилением иммуносупрессии.
Выводы: Ингибирование пути Hedgehog изменяет состав фибробластов и иммунную инфильтрацию в микросреде рака поджелудочной железы.
Трансляционная релевантность
Лучшее понимание роли сигнальных путей, дифференциально активируемых в разных популяциях фибробластов, прольет свет на сложность гетерогенности фибробластов при раке поджелудочной железы.Здесь мы показываем, что ингибирование пути Hedgehog изменяет состав фибробластов в сторону более воспалительной микросреды, открывая возможность для будущих комбинированных терапий.
Введение
Аденокарцинома протока поджелудочной железы (PDAC) — это летальная злокачественная опухоль с 5-летней выживаемостью примерно 10% (1). Микроокружение PDAC характеризуется обширной стромой, состоящей из неопухолевых клеток и компонентов внеклеточного матрикса (ECM), включая ассоциированные с раком фибробласты (CAF).Внутри стромы CAF секретируют факторы роста, иммуномодулирующие лиганды и белки ECM (2–4). Путь передачи сигналов Hedgehog активируется в PDAC CAFs и связан с отложением ECM и онкогенезом поджелудочной железы (5-10). В PDAC передача сигналов Hedgehog действует через паракринный механизм, посредством которого лиганды Hedgehog, полученные из опухоли, активируют передачу сигналов ниже по течению в CAF посредством взаимодействия с каноническим патченным рецептором Hedgehog (PTCh2) и корецепторами, GAS1, CDON и BOC (11-15). . Эти взаимодействия инициируют нижестоящий сигнальный каскад, опосредованный сглаживанием (SMO), что приводит к клеточным ответам, управляемым белками GLI, транскрипционными эффекторами пути Hedgehog (16).Однако сообщения об исследовании роли передачи сигналов Hedgehog в прогрессировании рака поджелудочной железы противоречивы. Первоначальные исследования показали, что ингибирование пути Hedgehog в моделях трансплантации ухудшает рост PDAC (5, 6, 12, 17, 18). Более того, острое ингибирование пути Hedgehog в генно-инженерной модели PDAC на мышах (GEMM) истощило ECM, увеличило перфузию сосудов и сделало опухоли сенсибилизированными к химиотерапии, обеспечивая умеренное преимущество в выживаемости (19). Однако эти результаты не нашли применения в клинических испытаниях, которые в значительной степени были безуспешными или даже пагубными для здоровья пациента, что привело к досрочному прекращению лечения (20, 21).Впоследствии генетическая инактивация лиганда Sonic Hedgehog (SHH) в контексте онкогенной модели рака поджелудочной железы на мышах, управляемой Kras , привела к кахексии и к низкодифференцированным и сильно васкуляризованным опухолям. фармакологическое ингибирование пути Hedgehog (22, 23). Эти несопоставимые результаты, при которых передача сигналов Hedgehog, по-видимому, одновременно способствует и ограничивают прогрессирование рака поджелудочной железы, указывают на потенциально плейотропные роли этого пути, которые еще предстоит полностью выяснить.
Здоровая поджелудочная железа является домом для различных популяций фибробластов (24, 25), которые обладают разным потенциалом роста во время канцерогенеза (26). Мы и другие недавно продемонстрировали, что PDAC CAFs также являются гетерогенной популяцией, состоящей из миофибробластных CAFs (myCAF), воспалительных CAFs (iCAF) и антигенпредставляющих CAFs (apCAF; ссылки 24, 25, 27-30). Учитывая различные функциональные роли, предлагаемые для этих подтипов CAF, изменения в соотношении этих популяций могут привести к различным результатам в прогрессии PDAC.Поэтому мы исследовали, влияет ли модуляция Hedgehog по-разному на подтипы CAF PDAC. Здесь мы показываем, что активация пути Hedgehog выше в myCAF по сравнению с iCAF как в мышиных, так и в человеческих PDAC. Наши данные предполагают, что управление более высокими уровнями передачи сигналов Hedgehog способствует росту опухоли поджелудочной железы, тогда как резкое ингибирование передачи сигналов Hedgehog снижает объем опухоли. Это исследование также определяет новую дозозависимую роль лиганда Indian Hedgehog (IHH) при раке поджелудочной железы. Наши данные в совокупности подтверждают представления о том, что передача сигналов Hedgehog является ключевым путем в поддержании подтипа myCAF при раке поджелудочной железы, и что ингибирование пути Hedgehog изменяет соотношение популяций myCAF / iCAF в PDAC и смещает воспалительный ответ в сторону более иммуносупрессивного микроокружения.
Материалы и методы
Одобрение исследований
Все процедуры и исследования на животных проводились в соответствии с руководящими принципами Институционального комитета по уходу за животными и их использованию при Мичиганском университете (Анн-Арбор, штат Мичиган, номер протокола, PRO00007983) и в холоде. Лаборатория Спринг-Харбор (Колд-Спринг-Харбор, Нью-Йорк).
Все процедуры, выполняемые в исследованиях с участием людей, соответствовали этическим стандартам институционального исследовательского комитета, в котором проводились исследования, а также Хельсинкской декларации 1964 года и ее более поздним поправкам или сопоставимым этическим стандартам.Все эксперименты с органоидами на людях были одобрены Институциональным наблюдательным советом (IRB) лаборатории Колд-Спринг-Харбор (Колд-Спринг-Харбор, штат Нью-Йорк) и проводились в соответствии с признанными этическими принципами (Хельсинкская декларация). Хирургические образцы опухолевой ткани или прилежащей / нормальной поджелудочной железы были получены от пациентов, направленных на процедуру Уиппла или дистальную резекцию поджелудочной железы в соответствии с IRB HUM00025339. Информированное письменное согласие было получено от каждого испытуемого.
Эксперименты на животных
Модели ортотопической трансплантации
Мыши C57BL / 6J (BL / 6J) (инвентарный номер 000664) были приобретены в лаборатории Джексона.Инъекции органоидных культур для создания ортотопически привитых органоидных опухолей (OGO) проводили, как описано ранее (29). Обычно 1–2,5 × 10 5 отдельных клеток, полученных из органоидных культур, ресуспендировали в виде 45 мкл суспензии 50% матригеля в PBS и вводили в поджелудочную железу. Опухоли получали с помощью ультразвукового сканера Vevo 3100 в двух или трех разных ориентациях по отношению к датчику. Объем опухоли измеряли под двумя углами, если это возможно, с помощью программы Vevo LAB Software (версия 2.2.0). Для создания моделей ортотопического рака поджелудочной железы с клеточной линией 7940b PDAC (31) 5 × 10 4 7940b клеток (штамм BL / 6J) были введены в поджелудочную железу BL / 6J или Gli1 lacZ / + гетерозиготных мышей, полученных от скрещивания Gli1 lacZ / lacZ животных, содержащихся на смешанном генетическом фоне (129S6 / SvEvTac; C57BL / 6J; Swiss Webster) с животными BL / 6J (32). Для ингибирования пути Hedgehog мышей ежедневно вводили через желудочный зонд в течение 12 дней, начиная с 6 дней после хирургической имплантации опухолевых клеток, либо 20 мг / кг LDE225 (Sonidegib, ChemieTek, # NVP-LDE225), ресуспендированных в PEG400 / 5. % раствора декстрозы (обработанного LDE) или такого же объема только раствора PEG400 / 5% декстрозы (обработанного носителем).
GEMM
Фон BL / 6J (> 20 обратных скрещиваний) Мыши KPC ( Kras LSL-G12D / + ; Trp53 LSL-R172H / + ; Pdx1 -Cre), использованные для исследования LD были описаны ранее (33). За мышами KPC наблюдали пальпацией живота и ультразвуком до тех пор, пока опухоли не достигли 6-8 мм в диаметре. Мышей KPC ежедневно вводили через желудочный зонд, как описано выше, либо носителем, либо LDE225 в течение 2 недель, а объем опухоли контролировали с помощью ультразвука.Опухоли получали с помощью ультразвукового сканера Vevo 3100 в двух или трех разных ориентациях по отношению к датчику. Объем опухоли измеряли под двумя углами, если возможно, с помощью программы Vevo LAB Software (версия 2.2.0). Фон BL / 6J iKC- Shh мышей были получены путем скрещивания iKC ( Ptf1a -Cre ERT ; Kras LSL-G12D / + ; подарок Криса Райта, Университет Вандербильта) и Shh ( CLEShh). ) мышей. Для; Мыши iKC- Shh , активация мутанта Kras и / или избыточная экспрессия Shh вызывалась введением трех отдельных обработок тамоксифена (Sigma, T-5648), ресуспендированного в кукурузном масле, по 5 мг на 30 г веса тела, когда возраст мышей составлял 8 недель.В этой модели эти мыши экспрессировали SHH мыши (мРНК; GenBank, # NM009170). Мыши выдерживали 34 недели после лечения тамоксифеном, а затем собирали поджелудочную железу для иммуноокрашивания. Для локализации SHH-отвечающих клеток in vivo мы использовали репортерных мышей Gli1 lacz / + (32). Соответственно, вводили тамоксифен и поджелудочную железу фиксировали в течение 1 часа 4% параформальдегидом (PFA) в течение 1 часа на льду, замораживали в соединении с оптимальной температурой резки (OCT) и делали криосрезы для окрашивания β-галактозидазой (β-gal) с использованием X-GAL субстрат.Срезы контрастировали с Nuclear Fast Red.
Анализ секвенирования одноклеточной РНК
Наборы данных были получены от Elyada и коллег (30), Peng и коллег (34) и Steele и коллег (35). Снижение размерности было выполнено в Scanpy (PMC5802054) с помощью анализа главных компонентов на верхних 3000 сильно вариабельных генах с последующей визуализацией однородного многообразия аппроксимации и проекции (UMAP) с использованием 30 основных компонентов. Кластеризация проводилась с помощью алгоритма Лувена (https: // doi.org / 10.1088 / 1742-5468 / 2008/10 / P10008) путем повторения параметра разрешения до тех пор, пока грубые типы клеток не могут быть идентифицированы с помощью дифференциально экспрессируемых канонических маркерных генов, которые затем подвергались дополнительным циклам уточнения кластера для аннотирования подтипов.
Клеточные линии и клеточная культура
Ihh клетки дикого типа (WT) и Ihh -нокаут (KO) 7940b KPC клетки и Gli1 lacZ / lacZ мышиных фибробластов (до пассажа номер 15) выращивали в среде DMEM с добавлением 10% FCS и 5% пенициллина / стрептомицина (Thermo Fisher Scientific, # 15140163), как описано ранее (15, 36).Органоиды PDAC T69A мыши (37) культивировали, как описано ранее, и трансплантировали при пассаже <15 (38). Все клетки культивировали при 37 ° C с 5% CO 2 . Аутентификация клеточной линии не проводилась. Mycoplasma Тестирование с помощью набора MycoAlert Mycoplasma Detection Kit (LT07-318, Lonza) проводилось ежемесячно в лаборатории Колд-Спринг-Харбор (Колд-Спринг-Харбор, штат Нью-Йорк) и в Университете Мичигана (Анн-Арбор, штат Мичиган), а также для каждой линии органоидов / клеток. был протестирован по крайней мере один раз после размораживания или изоляции и повторно протестирован перед экспериментами по ортотопической трансплантации.
Ihh CRISPR / Cas-9 KO и эктопическая экспрессия SHH
To KO Ihh , однонаправляющие РНК, нацеленные на экзон 1 гена Ihh , были отобраны из библиотеки GeCKO (39) и клонированы в pSpCas9 (BB ) -2A-Puro (PX459) V2.0 Векторная магистраль (Addgene, # 62988). Введение направляющих было подтверждено секвенированием по Сэнгеру. Вектор со вставленными направляющими трансфицировали липофектамином 2000 (Thermo Fisher Scientific, # 11668) в клетки KPC PDAC 7940b, и отдельные клоны отбирали путем добавления пуромицина.Клоны подвергали скринингу с помощью qRT-PCR с праймерами, нацеленными на экзон 3 Ihh . Отдельные клоны дополнительно проверяли секвенированием по Сэнгеру (данные не показаны). Для эктопической экспрессии полноразмерного SHH Shh клонировали из pBS mShh # CT258 (Addgene, # 13999) в вектор pMXs (Addgene) и экспрессировали в органоидах T69A PDAC, полученных из опухоли KPC (37). Затем с помощью бластицидина отбирали органоиды, и повышенную экспрессию SHH подтверждали с помощью ELISA.
ELISA
Для обнаружения SHH мы провели ELISA сред после выращивания культур органоидов T69A в течение 3-4 дней.Среды собирали, центрифугировали и анализировали, используя протокол производителя (MSHH00, R&D Systems).
Анализ qRT-PCR
РНКэкстрагировали с использованием набора PureLink Mini Kit (Thermo Fisher Scientific, # 12183025), как описано в протоколе производителя. Набор для обратной транскрипции кДНК высокой емкости (Thermo Fisher Scientific, # 4368814) использовали для обратной транскрипции РНК в кДНК. ПЦР в реальном времени использовали для Shh (Mm00436528_m1), Ihh (Mm_00439613_m1) и Ppia (Mm02342430_g1).ПЦР-амплификации проводили с использованием предварительно проверенных зондов для анализа экспрессии 20X TaqMan, 2X TaqMan Universal Mastermix (Thermo Fisher Scientific, # 4364340) и матрицы кДНК. Амплификации выполняли в дублирующих лунках в системе для ПЦР в реальном времени StepOne, и dCT были нормализованы до Ppia значений домашнего хозяйства.
Гистопатологический анализ и окрашивание ИГХ
Ткани фиксировали в течение ночи в 10% нейтральном забуференном формалине, заливали парафином и делали срезы. Стандартные процедуры использовались для ИГХ и иммунофлуоресценции (IF).Вкратце, ткани депарафинизировали с помощью ксилола или Histoclear и подвергали поиску антигена лимонной кислоты (Thermo Fisher Scientific, NC0359148) перед блокированием 1% BSA в PBS в течение 1 часа при комнатной температуре. Первичные антитела добавляли в течение ночи при 4 ° C, а затем вторичные антитела в течение 1 часа при комнатной температуре. Используемые антитела: альфа-актин гладких мышц (αSMA; Sigma, A2547, 1: 1000 или ab5694, Abcam), Ki67 (Cell Signaling Technology, 12202, 1: 100), подопланин (BioLegend, 127403, 1: 200), бета- галактозидаза (Abcam ab9361, 1: 2,500), E-Cadherin (технология передачи сигналов клеток, 14472S, 1:50 или 610181, BD Biosciences), фосфо-ERK (технология передачи сигналов клеток, 4370L, 1: 100), CD31 (технология передачи сигналов клеток , 77699, 1: 100), расщепленная каспаза 3 (Cell Signaling Technology, 9661, 1: 200), CD8A (Cell Signaling Technology, 98941, 1: 100) и FOXP3 (Cell signaling Technology, 12653, 1: 100).Для ИГХ гематоксилин использовали в качестве ядерного контрастного вещества. Для IF использовали вторичные антитела Alexa Fluor (1: 500). Ядра клеток контрастировали с помощью реагента Prolong Gold Antifade Reagent (Invitrogen, P10144) или средства ProLong Diamond Antifade Mountant с DAPI (Invitrogen, # P36935). Изображения были получены с использованием микроскопа Olympus BX53F, цифровой камеры Olympus DP80 и программного обеспечения CellSens Standard. Изображения конфокальной микроскопии были получены с использованием конфокальных микроскопов LeicaSP5X или SP8 и программного обеспечения Leica LAS. Окрашивание X-GAL и IF на β-gal проводили на замороженных срезах.Для замороженных срезов ткани фиксировали в 4% PFA в течение 1 часа при 4 ° C. Ткани трижды промывали PBS и инкубировали в течение 48 часов при качании при 4 ° C в 30% сахарозе перед погружением в соединение OCT. Замороженные срезы вырезали толщиной 10 мкм. Окрашивание X-GAL проводили с использованием 5 нмоль / л K3Fe (CN) 6 , 5 нмоль / л K4Fe (CN) 6 , 2 ммоль / л MgCl 2 , 0,01% Na-дезоксихолата, 0,02% NP- 40 и 1 мг / мл X-GAL (суспендированный в диметилформамиде), приготовленный в PBS при 37 ° C в течение 48 часов.После этого фиксированные ткани контрастировали с помощью Nuclear Fast Red в течение 10 минут и затем закрепляли с помощью Permount (VWR, # 100496-550). Окрашивание гематоксилином и эозином (H&E) и трихромом по Массону проводили в соответствии со стандартными протоколами. Гистопатологический анализ опухолей KPC проводился патологом поджелудочной железы слепым методом на деидентифицированных слайдах H&E. Опухоли были классифицированы как умеренно или слабо дифференцированная аденокарцинома. Оценка некроза указывала количество областей некроза с малым увеличением (объектив 10 ×) на 10 полей с малым увеличением на каждом слайде.
ISH и двойной ISH с co-IF
ISH были выполнены с помощью набора для мультиплексной флуоресцентной диагностики RNA Scope (# 323100, Advanced Cell Diagnostics) или колориметрических наборов (322371-коричневый или 322360-красный, Advanced Cell Diagnostics). согласно протоколу производителя. Использовали мышиные зонды для Gli1 (311001, Advanced Cell Diagnostics) и Shh (314361, Advanced Cell Diagnostics). Вкратце, свежесрезанные залитые парафином срезы перед окрашиванием запекали в течение 1 часа при 60 ° C.Затем предметные стекла депарафинизировали и обрабатывали перекисью водорода в течение 10 минут при комнатной температуре. Извлечение мишени выполняли в пароварке с кипячением в течение 15 минут, а затем слайды обрабатывали реагентом ProteasePlus (Advanced Cell Diagnostics, # 322381) в течение 30 минут. После этого зонд для РНК гибридизовали в течение 2 часов при 40 ° C. Стадия амплификации AMP5 выполнялась в течение 1 часа. Для двойных красителей ISH и co-IF сигнал был усилен с использованием материалов AMP, предоставленных в наборе ACD Multiplex Kit (320850, Advanced Cell Diagnostics).Сигнал был разработан с использованием 1–3 каналов пероксидазы хрена в зависимости от количества зондов и желаемого канала. По завершении образцы промывали PBS, а затем блокировали в течение 1 часа 20% ослиной сывороткой при комнатной температуре. Первичные антитела инкубировали в течение ночи при 4 ° C (αSMA, Sigma, A2547, 1: 1000; Podoplanin, BioLegend, 127403, 1: 200; и E-Cadherin, Cell Signaling Technology, 14472 или 3195, 1: 100). Вторичные антитела Alexa Flour (1: 500 в блокирующем буфере) инкубировали в течение 1 часа при комнатной температуре, а образцы трижды промывали в PBS.Слайды окрашивали DAPI и закрепляли с помощью ProLong Gold Antifade Mountant (Thermo Fisher Scientific, P36930).
Количественный анализ изображений
Количественный анализ проводился как минимум в трех случайных неперекрывающихся полях (увеличение, 20 ×) в каждом образце с использованием программного обеспечения ImageJ, Fiji версии 2.0.0-rc-69 / 1.52p или Halo (Indica Labs) для измерения процента положительной области. По крайней мере, три репрезентативных образца на группу были проанализированы слепым методом. Для окрашивания Gli1 ISH на рис.2M, изображения сканера Pannoramic SCAN (PerkinElmer) были количественно определены с использованием алгоритмов программного обеспечения Halo для определения структуры ткани и анализа области окрашивания красной точки по всей площади ткани. Для всех количественных измерений были исключены кровеносные сосуды, некротические области и соединительные ткани. Для количественного определения DAPI цветовые каналы каждого захваченного изображения были разделены и преобразованы в двоичные. Клетки подсчитывались автоматически (анализировать → анализировать частицы), и результаты нормализовались по общему количеству клеток на изображении.Для количественного определения частиц (таких как Gli1 puncta) в конкретном пятне захваченные изображения сначала преобразовывались в двоичные. Частицы отбирались с помощью бинарной маски канала, содержащего желаемое пятно (редактировать → выделение → создать маску), и автоматически подсчитывались (анализировать → анализировать частицы). Затем результаты были нормализованы к общему количеству клеток, положительных по данному маркеру. Флуоресцентная визуализация тканей выполнялась с помощью конфокального лазерного сканирования Leica TCS SP8 (Leica Microsystems) под управлением LAS AF 3.3.10134 программное обеспечение. Изображения IF были количественно определены с использованием модуля анализа населения в Volocity (Improvision). Светлопольные изображения тканей получали с помощью Axio Imager.A2 (Zeiss). Окрашенные срезы сканировали с помощью Aperio ScanScope CS и анализировали с использованием алгоритма ImageScope Positive Pixel Count. Для количественного определения трихромного пятна по Массону значения оттенка для синего и розового были измерены с использованием среднего значения оттенка 0,6 и ширины оттенка 0,854. Затем определяли процентную площадь коллагена путем вычисления процента синих пикселей по отношению ко всей окрашенной области.Для количественной оценки окрашивания αSMA и PDPN IHC с помощью программного обеспечения ImageScope был рассчитан процент ярких положительных пикселей по отношению ко всему срезу.
Проточная цитометрия и FACS
Опухоли обрабатывали, как описано ранее (29). Для проточного цитометрического анализа популяций myCAF, iCAF и apCAF клетки окрашивали в течение 30 минут антимышиным CD31-PE / Cy7 (102418, BioLegend), CD45-PerCP / Cy5.5 (103132, BioLegend), CD326 (Ep -CAM) -Alexa Fluor 488 (118210, BioLegend), PDPN-APC / Cy7 (127418, BioLegend), CD140a (PDGFRα) -PE (135905, BioLegend), Ly6C-APC (128015, BioLegend) и IA / IE ( MHC-II) -BV785 (107645, BioLegend) и в течение 15 минут с DAPI.Графики потока были созданы с помощью программного обеспечения FlowJo. Для сортировки фибробластов из опухолей KPC клетки окрашивали в течение 30 минут CD45-Alexa Fluor 488 (103122, BioLegend), CD326 (EpCAM) -Alexa Fluor 647 (118212, BioLegend), CD324 (E-cadherin) -Alexa Fluor 647. (147307, BioLegend) и PDPN-APC / Cy7 (127418, BioLegend) и в течение 15 минут с DAPI. DAPI — CD45 — EpCAM — ECAD — Клетки PDPN + были отсортированы на FACSAria Cell Sorter (BD) и обработаны для массового секвенирования РНК (RNA-seq).
RNA-seq и анализ данных
Образцы тотальной РНК выделяли с использованием TRizol в соответствии с инструкциями производителя. Концентрация и качество РНК были определены, и библиотеки мРНК, обедненные рибосомами, были подготовлены Мичиганским университетом (Анн-Арбор, Мичиган) Advanced Genomics Core. Библиотеки секвенировали с использованием 150 парных концов NovaSeq 6000 (Illumina). Данные доступны в базе данных NCBI Gene Expression Omnibus под регистрационным номером GSE156867. Библиотеки RNA-seq были сопоставлены с первичной сборкой GENCODE GRCm38 с использованием STAR (v2.7.3), а оценки экспрессии генов были выполнены с использованием RSEM (v1.2.29). DESeq2 (v1.22.2) использовался для идентификации дифференциально экспрессируемых генов с параметрами по умолчанию. Гены со скорректированными значениями P ниже 0,05 были выбраны в качестве значимых генов. Анализ обогащения путей проводился с использованием программы анализа обогащения набора генов (GSEA; v4.0.3) и коллекций канонических путей (cp_v7.0_all), доступных в MSigDB. Для создания наборов сигнатурных генов из исследования Олунда и его коллег (27) были отобраны 200 лучших генов дифференциальной экспрессии с повышенной и понижающей регуляцией в iCAF по сравнению с myCAF и определены как сигнатуры генов myCAF и iCAF соответственно.Точный тест Фишера, доступный в R, использовался для выполнения теста ассоциации между генами дифференциальной экспрессии и сигнатурами myCAF / iCAF. Все графики были получены с использованием данных R. RNA-seq для нормальной поджелудочной железы человека и опухолевых органоидов от Tiriac et al. (40).
Статистический анализ
Программное обеспечение GraphPad Prism использовалось для графического представления данных. Статистический анализ проводился с использованием двустороннего критерия Стьюдента t , и P <0,05 считалось статистически значимым.Односторонний дисперсионный анализ был выполнен, чтобы определить, было ли какое-либо значение в наборе данных. За этим последовало апостериорных тестов с использованием метода разницы в средних по Тьюки с P <0,05, считающимся статистически значимым.
Цитометрия по времени пролета
Опухоли переваривали, как описано ранее (29). До 1 × 10 6 клеток окрашивали цисплатином Cell-ID (1,67 мкмоль / л) в течение 5 минут при комнатной температуре. Вкратце, после гашения реакции с цисплатином 5-кратным объемом буфера для окрашивания клеток MaxPar клетки центрифугировали при 300 x 92 · 106 g в течение 5 минут.Затем клетки окрашивали коктейлем антител на клеточной поверхности (каталожные номера и разведения см. В дополнительной таблице S1) в объеме 100 мкл буфера для окрашивания клеток MaxPar в течение 30 минут при комнатной температуре. После двукратной промывки в 1 мл буфера для окрашивания клеток MaxPar клетки фиксировали в 1,6% свежеприготовленном растворе формальдегида в течение 10 минут при комнатной температуре. Клетки дважды промывали 2 мл буфера для окрашивания клеток MaxPar, затем ресуспендировали в 1 мл раствора для интеркаляции клеток (125 нмоль / л Cell-ID Intercalator-Ir в Maxpar Fix и перманентном буфере) и отправляли в ядро проточной цитометрии в Университете Нью-Йорка. Медицинский центр Рочестера (Рочестер, штат Нью-Йорк), где была завершена подготовка образца и проведен анализ масс-цитометра CyTOF2.
Цитометрия с помощью анализа данных времени пролета
Необработанные файлы FCS были проанализированы с использованием программного обеспечения Premium CytoBank (cytobank.org). Данные были проверены на качество окрашивания и нормализованы с использованием внутренних стандартов для шариков. Живые синглетные клетки идентифицировали с использованием комбинации интеркалятора ДНК Ir191 и интенсивности окрашивания цисплатином Pt195. Было выполнено ручное стробирование.
Результаты
Активация пути Hedgehog выше в myCAF по сравнению с iCAF в PDAC
Пренеопластическая интраэпителиальная неоплазия поджелудочной железы (PanIN) и инвазивный PDAC связаны с повышенной экспрессией Hedgehog-лигандов (у 5 или нормальных лигандов поджелудочной железы низкий уровень экспрессии). , 6).Чтобы оценить экспрессию компонентов пути Hedgehog в различных клеточных компартментах поджелудочной железы, мы проанализировали данные одноклеточной последовательности РНК (scRNA-seq) из соседней нормальной ткани человека или нормальной поджелудочной железы ( n, = 3) и PDAC ( n = 16) образцов (рис. 1А; ссылка 35). В соответствии с предыдущими исследованиями (5, 6, 13) экспрессия лигандов SHH и IHH была ограничена эпителиальными клетками и увеличивалась в PDAC по сравнению с соседними нормальными / нормальными тканями, тогда как лиганд пустыни Hedgehog ( DHH ) не обнаружен в эпителиальных клетках, но экспрессируется на низких уровнях в эндотелиальных клетках (рис.1Б). Аналогичным образом, анализ основной последовательности РНК соседней нормальной / нормальной поджелудочной железы человека ( n = 4) и тканей, полученных от пациента с PDAC ( n = 178) из набора данных Атласа генома рака, показал значительное увеличение SHH и IHH в образцах PDAC по сравнению с контролями, тогда как уровни DHH существенно не изменились (дополнительный рисунок S1A). Количество транскриптов SHH и IHH также значительно увеличилось в органоидах PDAC, полученных от пациентов ( n = 63) по сравнению с органоидами, выделенными из нормальной поджелудочной железы человека ( n = 14; исх.40; Дополнительный рис. S1B и S1C). Напротив, экспрессия транскрипционных мишеней Hedgehog, включая GLI1, PTCh2, PTCh3 и HHIP , была обогащена фибробластами PDAC вместе с компонентами пути, такими как SMO (41), GAS1, CDON , и BOC (рис. 1B; ссылки 11–13, 15, 42, 43). Наконец, анализ двух ранее опубликованных наборов данных scRNA-seq для PDAC человека и нормальной поджелудочной железы (30, 34) подтвердил, что SHH и IHH ограничены эпителиальным компартментом и выше в PDAC по сравнению с нормальными тканями, тогда как ген-мишень Hedgehog Экспрессия была в значительной степени ограничена фибробластами и, как правило, выше в PDAC по сравнению с доброкачественными образцами (дополнительный рис.S1D – S1G).
Рисунок 1.Активация пути Hedgehog выше в myCAF по сравнению с iCAF в PDAC. A, UMAP визуализация популяций клеток из scRNA-seq соседних нормальных / нормальных тканей поджелудочной железы человека ( n = 3) и PDAC ( n = 16) тканей. Нормальные образцы (92 · 106 n 92 · 107 = 3) были изолированы от пациентов, перенесших операцию по поводу аденомы двенадцатиперстной кишки, ампулярной карциномы или PDAC, где в резекцию была включена не вовлеченная часть поджелудочной железы. Кластеры различных типов ячеек имеют цветовую маркировку.Данные получены от Стила и его коллег (35). B, Визуализация точечной диаграммы уровня экспрессии гена пути Hedgehog (интенсивность цвета) и частоты (размер точки) в различных популяциях клеток соседней нормальной / нормальной поджелудочной железы человека (синий, 92 · 106 n = 3) и PDAC (красный, 92 · 106 n = 16) образцов из A . Рамками выделены лиганды Hedgehog ( SHH, IHH и DHH ) и мишени Hedgehog ( GLI1, PTCh2, PTCh3 и HHIP ). C, UMAP визуализация scRNA-seq кластеров фибробластов в объединенных образцах соседней нормальной поджелудочной железы человека ( n = 2) и PDAC ( n = 6) образцов.Различные кластеры подтипа CAF имеют цветовую кодировку. Данные получены от Elyada с коллегами (30). D, UMAP визуализация экспрессии маркеров myCAF ( ACTA2 , кодирующей αSMA и TAGLN ) и iCAF ( IL6 и CLEC3B ) в кластерах фибробластов в образцах PDAC человека из C . E, UMAP визуализация экспрессии мишени Hedgehog ( GLI1 и PTCh2 ) в кластерах фибробластов в образцах PDAC человека из C . F, Тепловые карты нормализованной экспрессии мишеней Hedgehog ( GLI1, PTCh2, PTCh3 и HHIP ), рецептора Hedgehog ( SMO ) и корецепторов ( GAS1, CDON и в каждом) кластер фибробластов в образцах PDAC человека из C . Цвета показывают количество генов в логарифмической шкале. G, Тепловая карта масштабируемой экспрессии лигандов Hedgehog ( Shh, Ihh и Dhh ), мишеней Hedgehog ( Gli1, Ptch2, Ptch3 и Hhip ), рецептора Hedgehog и Smoor корецепторов ( Gas1, Cdon и Boc ) в различных популяциях клеток опухолей поджелудочной железы модели PDAC мыши KPC ( n = 4).Данные масштабируются таким образом, что кластер с самым низким средним выражением = 0 и самым высоким = 1 для каждого гена. Данные получены от Elyada с коллегами (30). H, Репрезентативный ISH РНК Gli1 (белый) и co-IF PDPN (зеленый) в опухоли KPC. Контрастное пятно, DAPI (синий). Масштабная линейка 20 мкм. I, Количественное определение красителя Gli1 в клетках PDPN + (CAF) и PDPN — (не CAF) в опухолях KPC. Результаты показывают среднее значение ± стандартная ошибка среднего для семи биологических повторов.***, P <0,001, непарный тест Стьюдента t . J, UMAP-визуализация scRNA-seq кластеров фибробластов в опухолях KPC ( n = 4) от Elyada et al. (30). Различные кластеры подтипа CAF имеют цветовую кодировку. K, Тепловые карты нормализованной экспрессии мишеней Hedgehog ( Gli1, Ptch2, Ptch3 и Hhip ), рецептора Hedgehog ( Smo ) и корецепторов ( Gas1, Cdon и Boc) в каждом кластер фибробластов из J .Цвета показывают количество генов в логарифмической шкале. L, Репрезентативный ISH РНК Gli1 (белый) и co-IF PDPN (зеленый) и αSMA (красный) в опухоли KPC. Контрастное пятно, DAPI (синий). Масштабная линейка 20 мкм. Стрелка указывает на клетку PDPN + αSMA + с более низкой экспрессией Gli1 ; сплошная стрелка указывает на клетку PDPN + αSMA + с более высокой экспрессией Gli1 . M, Количественное определение красителя Gli1 в клетках αSMA + PDPN + (myCAFs) и αSMA — PDPN + (не myCAFs) в опухолях KPC.Результаты показывают среднее значение ± стандартная ошибка среднего для семи биологических повторов. **, P <0,01, непарный тест Стьюдента t .
Затем мы исследовали, была ли передача сигналов Hedgehog по-разному активна в разных подтипах CAF PDAC человека. Анализ фибробластов из нашего ранее опубликованного набора данных scRNA-seq человеческого PDAC (30) выявил обогащение генов-мишеней Hedgehog ( GLI1, PTCh2, PTCh3 и HHIP ) в myCAF по сравнению с iCAF (рис. 1C – F). , тогда как основные компоненты пути Hedgehog ( SMO, GAS1, CDON и BOC ) были экспрессированы в обоих компартментах (рис.1F). Эти наблюдения были подтверждены во втором наборе данных scRNA-seq для PDAC человека и нормальной поджелудочной железы (ссылка 34; дополнительный рисунок S1H-S1K). Чтобы определить, были ли эти наблюдения повторены на мышиных моделях PDAC, мы проанализировали наш ранее опубликованный набор данных scRNA-seq автохтонных опухолей поджелудочной железы из модели PDAC у мышей KPC (30, 33). Подобно человеческому PDAC, мышиные опухоли KPC экспрессируют лиганды Hedgehog преимущественно в эпителиальных клетках, в то время как фибробласты являются основным типом клеток, чувствительных к Hedgehog, в зависимости от экспрессии гена-мишени (рис.1G). РНК ISH Gli1 , гена-мишени и медиатора транскрипции пути Hedgehog (32), в сочетании с ко-IF маркера панфибробластов подопланина (PDPN) подтвердили, что экспрессия Gli1 ограничена клетками PDPN + в опухолях KPC (рис. 1H и I). Наконец, аналогично тому, что мы наблюдали в образцах PDAC человека, анализ scRNA-seq CAF из опухолей KPC продемонстрировал обогащение экспрессии гена-мишени Hedgehog в myCAF по сравнению с другими подтипами CAF (рис.1J и K; Дополнительный рис. S1L). Соответственно, ISH РНК Gli1 с ко-IF PDPN и маркером myCAF, альфа-актином гладких мышц (αSMA), показал более высокую экспрессию Gli1 в myCAF (αSMA + PDPN + ) по сравнению с не-myCAF. (αSMA — PDPN + ) в опухолях KPC (рис. 1L и M). Примечательно, что в популяции myCAF мы обнаружили вариабельные уровни Gli1 (рис. 1L), что согласуется с гетерогенностью, наблюдаемой в нашем анализе scRNA-seq фибробластов PDAC человека (рис.1E).
В совокупности эти данные предполагают, что, хотя все подтипы CAF способны отвечать на лиганды SHH и IHH, полученные из опухоли, на основе экспрессии SMO, GAS1, CDON и BOC , передача сигналов Hedgehog в myCAF активируется у мышей и людей. PDAC.
Ингибирование пути Hedgehog нарушает рост PDAC
Чтобы понять функциональную роль передачи сигналов Hedgehog, мы исследовали последствия гиперактивации пути Hedgehog на прогрессирование PDAC.SHH эктопически экспрессировался в KPC опухолевых органоидах PDAC (37), которые мы затем использовали для создания модели PDAC на мышах OGO (ссылка 38; дополнительные рисунки S2A и S2B). В соответствии с предыдущими данными (7), эктопическая экспрессия SHH в эпителиальном компартменте приводила к значительному увеличению объема опухоли по сравнению с контролем (дополнительный рисунок S2C). Повышенная эпителиальная экспрессия Shh была подтверждена с помощью РНК ISH Shh с ко-IF эпителиального маркера E-кадгерина (ECAD) и маркера фибробластов PDPN (дополнительный рис.S2D и S2E). Более того, повышенная активация пути Hedgehog в фибробластах была подтверждена с помощью РНК ISH Gli1 с ко-IF αSMA и PDPN (дополнительные фиг. S2F и S2G). Хотя экспрессия SHH ранее была связана с образованием фиброзной стромы (7–9, 12), в этой модели трансплантации мы не наблюдали значительного изменения десмоплазии, измеренного с помощью окрашивания трихрома Массона и αSMA (дополнительный рисунок S2H – S2J). ). Вместо этого сверхэкспрессия SHH привела к значительному увеличению фосфо-ERK (p-ERK) и снижению уровней расщепленной каспазы 3 в эпителиальных клетках (дополнительный рис.S2H, S2K и S2L), что соответствует изменению роста опухоли.
Для дальнейшей оценки роли передачи сигналов Hedgehog на ранних стадиях развития PDAC мы активировали эпителиальную экспрессию Shh в iKC ( Ptf1a -Cre ERT ; Kras LSL-G12D / + ) индуцируемый тамоксифеном GEMM PDAC (ссылка 44; дополнительный рисунок S2M). В iKC; Shh мышей, активация мутанта Kras и сверхэкспрессия SHH одновременно индуцируются в ацинарных клетках рекомбинацией Cre при введении тамоксифена.Чтобы подтвердить увеличение активации Hedgehog, Ptf1a Cre ERT ; мышей Shh скрещивали с Gli1 lacZ / + животных, которые считывали активность пути Hedgehog путем измерения активности β-гал (32). Полученный Ptf1a Cre ERT ; Gli1 lacZ / + ; Затем мышей Shh лечили тамоксифеном в возрасте 8 недель и в возрасте 34 недель (дополнительная фигура S2N). По сравнению с контролем Ptf1a Cre ERT ; Gli1 lacz / + pancreata, Ptf1a Cre ERT ; Gli1 lacZ / + ; Shh поджелудочной железы имеет повышенное количество клеток X-GAL + (дополнительный рис.S2O), подтверждая, что избыточная экспрессия SHH в ацинарных клетках приводит к повышенной активации Hedgehog. Примечательно, что iKC; У мышей Shh наблюдалось увеличение предопухолевых поражений PanIN по сравнению с мышами iKC, получавшими тамоксифен, мышами WT и Ptf1a Cre ERT ; Shh контролирует (Supplementary Fig. S2P), что согласуется с мнением, что активация пути Hedgehog способствует образованию PDAC (7-9, 12). Кроме того, в соответствии с более развитыми поражениями, поджелудочная железа от iKC; Shh мышей демонстрировали больше p-ERK по сравнению с мышами iKC, получавшими тамоксифен, мышами WT и Ptf1a Cre ERT ; Шх органы управления (дополнительный рис.S2P). Сверхэкспрессия SHH в iKC; Shh мышей также приводили к повышенной десмоплазии, по оценке с помощью красителей трихрома Массона и αSMA (дополнительный рис. S2P), что указывает на то, что активация пути Hedgehog до образования опухоли в этом GEMM оказывает большее влияние на ремоделирование стромы по сравнению с моделями трансплантации мышей. Клетки PDAC (дополнительный рис. S2H – S2J).
Затем мы исследовали влияние истощения лиганда Hedgehog на прогрессирование PDAC. В то время как эффект делеции SHH в PDAC был исследован ранее (22, 23), роль IHH не рассматривалась, несмотря на наблюдение, что оба лиганда экспрессируются как при раке поджелудочной железы человека, так и при раке поджелудочной железы мыши.Поэтому мы удалили Ihh с использованием CRISPR / Cas9 в клеточной линии KPC PDAC, которая экспрессировала низкие уровни Shh (ссылка 31; дополнительные рисунки S3A и S3B). Ihh клетки PDAC WT индуцировали активацию пути Hedgehog в совместно культивируемых фибробластах Gli1 lacZ / lacZ (15), о чем свидетельствует окрашивание на β-gal, тогда как клетки Ihh -KO не смогли этого сделать (дополнительный рисунок S3C). ). Ортотопическая трансплантация клеток Ihh -KO PDAC мышам BL / 6J не показала различий в росте опухоли по сравнению с контрольными клетками WT (дополнительные фиг.S2A и S2B и S3D-S3F), хотя ингибирование пути Hedgehog было подтверждено с помощью ISH РНК Gli1 (рис. 2C и D). Мы предположили, что клетки Ihh -KO PDAC, трансплантированные мышам BL / 6J, потенциально сохраняли активность Hedgehog на низком уровне. Поскольку мы ранее определили дозозависимую роль передачи сигналов Hedgehog в прогрессировании рака поджелудочной железы (15), мы оценили влияние делеции Ihh на прогрессирование PDAC у мышей Gli1 lacz / + , у которых отсутствует одна копия Gli1. .Эти мыши обладают системно более низкими уровнями передачи сигналов Hedgehog и обнаруживают дефекты в ответных на Hedgehog процессах гомеостаза и восстановления взрослых (45–47). Примечательно, что трансплантация клеток Ihh -KO PDAC в поджелудочную железу мышей Gli1 lacz / + привела к образованию значительно меньших опухолей по сравнению с контрольными клетками WT (рис. 2A и B; дополнительный рис. S3D – S3F). Ингибирование пути Hedgehog было подтверждено окрашиванием X-GAL и co-IF для β-gal и αSMA (фиг. 2C и E). Затем мы использовали селективный антагонист SMO для фармакологического ингибирования передачи сигналов Hedgehog у мышей BL / 6J и Gli1 lacz / + , ортотопически трансплантированных с клетками PDAC.Как только пальпируемое образование было обнаружено, мышей обрабатывали один раз в день через желудочный зонд с ингибитором SMO, LDE225 (48), в течение 12 дней (рис. 2F). LDE225 — эффективный ингибитор пути Hedgehog, действующий ниже всех лигандов Hedgehog путем нацеливания на SMO, и активно используется в клинической практике (48–50). Подобно тому, что наблюдалось с делецией Ihh , фармакологическое ингибирование пути Hedgehog приводило к меньшим опухолям только у мышей Gli1 lacz / + (рис. 2G; дополнительный рис.S3G – S3I), которые лишены одной копии Gli1 и, следовательно, имеют более низкие уровни передачи сигналов Hedgehog на исходном уровне до трансплантации опухолевых клеток. РНК ISH Gli1 , окрашивание X-GAL и ко-IF β-gal / αSMA подтвердили ингибирование пути Hedgehog в обеих моделях (рис. 2H – J). В целом, эти данные согласуются с предыдущими сообщениями, в которых у Gli1 -нулевых животных наблюдалось снижение прогрессирования предопухолевых поражений поджелудочной железы, а потеря одной копии Gli1 оказала влияние на гомеостаз ткани поджелудочной железы (45, 51).
Рисунок 2.Ингибирование пути Hedgehog ухудшает рост PDAC. A, Схема ортотопической трансплантации 7940b клеток KPC PDAC с Ihh WT или KO мышам BL / 6J или Gli1 lacZ / + . B, Вес опухоли на 18 день после трансплантации эксперимента из A . Результаты показывают среднее значение ± стандартная ошибка среднего для шести биологических повторов. **, P <0,01, однофакторный дисперсионный анализ. C, Репрезентативные изображения ISH РНК Gli1 в опухолях Ihh WT или -KO у мышей BL / 6J (слева), а также окрашивание X-GAL и ко-IF αSMA (пурпурный), β-gal ( зеленый) и DAPI (синий) в опухолях Ihh WT или -KO у мышей Gli1 lacZ / + (в центре и справа).Вкладыши, увеличения. Масштабные линейки: 50 мкм (слева), 100 мкм (в центре) и 25 мкм (справа). D, Количественное определение Gli1 РНК ISH в опухолях Ihh WT или -KO у мышей BL / 6J. Результаты показывают среднее значение ± SEM для шести биологических повторов. ***, P <0,001, непарный тест Стьюдента t . E, Количественное определение окрашивания X-GAL в опухолях Ihh WT или -KO у мышей Gli1 lacZ / + . Результаты показывают среднее значение ± SEM для шести биологических повторов.**, P <0,01, непарный тест Стьюдента t . F, Схема ортотопической трансплантации клеток 7940b KPC PDAC мышам BL / 6J или Gli1 lacZ / + с последующей 12-дневной обработкой 20 мг / кг ингибитора SMO, LDE225 или носителя путем ежедневного перорального приема желудочный зонд. G, Масса опухоли на 18 день после трансплантации эксперимента из F . Результаты показывают среднее значение ± SEM для 4–6 биологических повторов. ***, P <0,001, однофакторный дисперсионный анализ. H, Репрезентативные изображения ISH РНК Gli1 в опухолях, обработанных носителем или LDE225, у мышей BL / 6J (слева), а также окрашивания X-GAL и ко-IF αSMA (красный), β-gal (зеленый ) и DAPI (синий) в опухолях, обработанных носителем или LDE225, у мышей Gli1 lacZ / + (в центре и справа). Вкладыши, увеличения. Масштабные линейки: 50 мкм (слева), 100 мкм (в центре) и 25 мкм (справа). I, Количественное определение Gli1 РНК ISH в опухолях, обработанных LDE225 или носителем, у мышей BL / 6J.Результаты показывают среднее значение ± SEM для четырех биологических повторов. ***, P <0,001, непарный тест Стьюдента t . J, Количественное определение окрашивания X-GAL в опухолях, обработанных LDE225 или носителем, у мышей Gli1 lacZ / + . Результаты показывают среднее значение ± SEM для четырех биологических повторов. ***, P <0,001, непарный тест Стьюдента t . K, Схема 2-недельного лечения мышей KPC с опухолью 20 мг / кг LDE225 или носителя посредством ежедневного перорального введения через желудочный зонд.У / С, УЗИ. л, Объем опухоли, измеренный ультразвуком опухолей KPC, обработанных носителем ( n = 9) и LDE225 ( n = 8), из K . Результаты показывают среднее значение ± SEM. *, P <0,05, непарный тест Стьюдента t . M, Репрезентативная РНК ISH Gli1 в 2-недельных опухолях KPC, обработанных носителем ( n = 11) и LDE225 ( n = 10). Шкала 100 мкм. N, Количественное определение красителя Gli1 в опухолях KPC, обработанных носителем ( n = 11) и LDE225 ( n = 10).Результаты показывают среднее значение ± SEM. ***, P <0,001, непарный тест Стьюдента t .
Поскольку ускоренная кинетика роста опухоли этих моделей трансплантации PDAC могла предотвратить наблюдение более тонких изменений объема опухоли при ингибировании пути Hedgehog, мы исследовали эффект использования LDE225 в модели мышей KPC. Мышей KPC включали в контрольную или лечебную группу исследования, как только опухоли были обнаружены с помощью ультразвука. Интересно, что 2-недельное лечение LDE225 привело к нарушению роста опухоли в модели KPC (рис.2К и Л; Дополнительный рис. S3J). Устранение передачи сигналов Hedgehog было подтверждено с помощью РНК ISH Gli1 (фиг. 2M и N). Хотя никаких изменений в некрозе опухолей, дифференцировке, эпителиальном апоптозе и пролиферации эпителия не наблюдалось (дополнительный рис. S3K – S3P), мы наблюдали умеренное, но значительное снижение уровней p-ERK в эпителиальных клетках при ингибировании пути Hedgehog (дополнительный рис. S3Q и S3R).
Взятые вместе, эти данные показывают, что, в соответствии с предыдущими исследованиями, передача сигналов Hedgehog в целом способствует прогрессированию PDAC, что как SHH, так и IHH вносят вклад в рост рака поджелудочной железы, и что степень вклада передачи сигналов Hedgehog в рост опухоли зависит от уровня активности пути Hedgehog.
Ингибирование пути Hedgehog изменяет компартмент фибробластов в PDAC
В отличие от предыдущей работы, которая ингибировала передачу сигналов Hedgehog (19, 22, 23), 2-недельное лечение LDE225 не изменяло количество отложения коллагена, как измерено с помощью окрашивания трихрома Массона. (Дополнительный рис. S4A и S4B) или сосудистая сеть, как оценивается IHC для CD31 (Дополнительный рис. S4C и S4D), в опухолях KPC. Однако 2-недельное ингибирование пути Hedgehog привело к снижению количества клеток PDPN + (рис.3А и В). Для дальнейшего исследования мы выделили PDPN + DAPI — CD45 — EpCAM — CAF из 2-недельных опухолей KPC, обработанных носителем и LDE225, с помощью FACS и выполнили объемную последовательность РНК (рис. 3C; дополнительная информация). Рис. S4E). GSEA и дифференциальная экспрессия мишеней Hedgehog подтвердили значительное подавление передачи сигналов Hedgehog в CAF, выделенных из опухолей, обработанных LDE225, по сравнению с контролем, обработанным носителем (дополнительный рисунок S4F – S4H). Затем мы оценили, какие секретируемые факторы подавлялись обработкой LDE225 в наборе данных RNA-seq.Мы обнаружили, что факторы роста, включая Wnt5a, Tgfb1 и Sema3a , а также ангиогенные факторы, такие как Angpt4 и Vegfa , подавлялись в фибробластах, выделенных из опухолей KPC, обработанных LDE225, по сравнению с контролем (дополнительный рис. S4I). Кроме того, гены, кодирующие белки ECM, включая Col4a1, Col4a6 и Mmp15 , также подавлялись при обработке LDE225 (дополнительный рисунок S4I). Таким образом, мы обнаружили зависимые от ингибирования Hedgehog изменения экспрессии генов, связанных с ECM, и дополнительные изменения в факторах, секретируемых фибробластами, которые могли бы объяснить снижение роста опухоли.Более того, в соответствии с наблюдаемым уменьшением количества фибробластов сигнатура гена пролиферации клеток и маркеры пролиферации были значительно снижены в CAF, выделенных из опухолей, обработанных LDE225, по сравнению с контрольными группами, обработанными носителем (фиг. 3D и E). Учитывая, что активность пути Hedgehog выше в myCAF по сравнению с другими подтипами CAF в опухолях KPC (рис. 1K – M), мы исследовали последствия лечения LDE225 на популяциях myCAF и не-myCAF. ИГХ αSMA и обнаружение ко-IF αSMA и PDPN для идентификации myCAF (PDPN + αSMA + ) и не-myCAFs (PDPN + αSMA —) не только выявили снижение количества αSMA + myCAF. (Рисунок.3F и G; Дополнительный рис. S4J и S4K), что согласуется с предыдущими исследованиями (19, 22, 23), но также показало увеличение не-myCAF (рис. 3F и H) в опухолях, обработанных LDE225, по сравнению с контрольными группами, обработанными носителем. В целом, эти изменения фибробластов привели к значительному снижению соотношения myCAF / non-myCAF (дополнительный рисунок S4L).
Рисунок 3.Двухнедельное ингибирование пути Hedgehog изменяет компартмент фибробластов в PDAC. A, Типичный ИГХ PDPN в опухолях KPC, обработанных носителем ( n = 10) и LDE225 ( n = 11).Масштабная линейка 50 мкм. B, Количественное определение окраски PDPN в опухолях KPC, обработанных носителем ( n = 10) и LDE225 ( n = 11). Результаты показывают среднее значение ± SEM. **, P <0,01, непарный тест Стьюдента t . C, Схема стратегии FACS для объемной последовательности РНК фибробластов из опухолей KPC, обработанных носителем и LDE225. D, GSEA сигнатуры клеточного цикла в FACS-отсортированных CAF из опухолей KPC, обработанных LDE225 ( n = 2), по сравнению с FACS-отсортированными CAF из обработанных носителем контролей ( n = 3). E, RNA-seq экспрессия маркеров пролиферации ( Mki67, Ccnb2 и Cks2 ) в отсортированных по FACS CAF из носителя- ( n = 3) и обработанных LDE225 ( n = 2) KPC опухоли. Результаты показывают среднее значение ± SEM. *, P <0,05; **, P <0,01; ***, P <0,001, непарный тест Стьюдента t . F, Co-IF αSMA (красный) и PDPN (зеленый) в опухолях KPC, обработанных носителем ( n = 7) и LDE225 ( n = 4).Контрастное пятно, DAPI. Масштабная линейка 20 мкм. G, Количественное определение myCAF (αSMA + PDPN + DAPI + ) в опухолях KPC, обработанных носителем ( n = 7) и обработанных LDE225 ( n = 4). Результаты показывают среднее значение ± SEM. P = 0,05, непарный тест Стьюдента t . H, Количественное определение не-myCAF (αSMA — PDPN + DAPI + ) в опухолях KPC, обработанных носителем ( n = 7) и обработанных LDE225 ( n = 4).Результаты показывают среднее значение ± SEM. *, P <0,05, непарный тест Стьюдента t .
Наши результаты показывают, что пролиферация фибробластов влияет на лечение LDE225, что может частично объяснить уменьшение количества миофибробластов, наблюдаемое в опухолях, обработанных ингибитором Hedgehog. В целом, наши данные подтверждают роль ингибитора пути Hedgehog, LDE225, в нацеливании на компартмент фибробластов и преимущественно в истощении миофибробластов по сравнению с не-myCAFs в PDAC.
Ингибирование пути Hedgehog изменяет соотношение myCAF и iCAF в PDAC
Затем мы использовали ранее установленный подход проточной цитометрии (29, 30) для дальнейшей оценки myCAF (PDPN + MHCII — LY9C42 — LY9C42) (PDPN + MHCII — LY6C + ) и apCAF (PDPN + LY6C — MHCII + ) в 2-недельных опухолях KPC, обработанных носителем и LDE225 (рис.4А). Ингибирование пути Hedgehog привело к снижению количества myCAF и значительному увеличению количества iCAF (рис. 4B – E), что привело к увеличению соотношения iCAF / myCAF (рис. 4F).
Рисунок 4.Двухнедельное ингибирование пути Hedgehog изменяет соотношение myCAF и iCAF в PDAC. A, Схема стратегии проточной цитометрии для myCAF и iCAF из 2-недельных опухолей KPC, обработанных носителем и LDE225. B, Типичные потоковые диаграммы, показывающие стратегию стробирования для анализа DAPI — CD45 — CD31 — EpCAM — PDPN + CAF в 2-недельном носителе- ( n = 7) и Обработанные LDE225 ( n = 6) опухоли KPC. C, Анализ проточной цитометрии myCAF (рассчитанный из синглетов DAPI + ) в опухолях KPC, обработанных носителем ( n = 7) и LDE225 ( n = 6). Результаты показывают среднее значение ± SEM. Непарный Студент t тест . D, Анализ проточной цитометрии iCAF (рассчитанный из синглетов DAPI + ) в опухолях KPC, обработанных носителем ( n = 7) и обработанных LDE225 ( n = 6). Результаты показывают среднее значение ± SEM. *, P <0,05, непарный тест Стьюдента t . E, Пропорции подтипов myCAF, iCAF и apCAF из ворот PDPN + в опухолях KPC, обработанных носителем ( n = 7 = 7, вверху) и LDE225 ( n = 6, внизу), как измерено методом проточной цитометрии. Результаты показывают средний процент биологических повторов. F, Проточно-цитометрический анализ отношения iCAF / myCAF из ворот PDPN + в опухолях KPC, обработанных носителем ( n = 7) и обработанных LDE225 ( n = 6). Результаты показывают среднее значение ± SEM.*, P <0,05, непарный тест Стьюдента t . G, GSEA сигнатуры гена myCAF в отсортированных по FACS CAF из 2-недельных обработанных LDE225 опухолей KPC ( n = 2) по сравнению с отсортированными по FACS CAF от обработанных носителем контролей ( n = 3). Сигнатура гена myCAF была определена в исследовании Öhlund с коллегами (27). H, RNA-seq экспрессия маркеров myCAF ( Acta2, Thy1, Tagln и Tgfb1 ) в отсортированных по FACS CAF из носителя ( n = 3) и обработанных LDE225 ( n = 2). ) Опухоли KPC.Результаты показывают среднее значение ± SEM. Статистической разницы не наблюдалось при расчете с помощью непарного теста Стьюдента t . I, GSEA сигнатуры гена iCAF в отсортированных по FACS CAF из 2-недельных обработанных LDE225 опухолей KPC ( n = 2) по сравнению с отсортированными по FACS CAF от обработанных носителем контролей ( n = 3). Сигнатура гена iCAF была определена в исследовании Öhlund с коллегами (27). J, RNA-seq экспрессия маркеров iCAF ( Dpt, Clec3b, C3 и Cxcl12 ) в отсортированных по FACS CAF из носителя ( n = 3) и обработанных LDE225 ( n = 2). ) Опухоли KPC.Результаты показывают среднее значение ± SEM. *, P <0,05, непарный тест Стьюдента t .
Затем мы исследовали, приводит ли ингибирование пути Hedgehog к изменению сигнатур iCAF и myCAF, используя ранее установленные сигнатуры генов (27), в большом наборе данных RNA-seq для FACS-сортированных CAF из 2-недельного носителя и LDE225- обработанные опухоли KPC (фиг. 3C; дополнительная фиг. S4E). В соответствии с данными проточной цитометрии этот анализ выявил подавление сигнатуры экспрессии гена myCAF, а также подавление специфических маркеров myCAF при лечении LDE225 (рис.4G и H). Кроме того, мы наблюдали повышенную регуляцию сигнатуры iCAF и маркеров iCAF в опухолях KPC, обработанных LDE225, по сравнению с контрольными опухолями, обработанными носителем (фиг. 4I и J).
Эти данные показывают, что передача сигналов Hedgehog является ключевым путем для поддержания фенотипа myCAF, и его ингибирование изменяет соотношение myCAFs и iCAFs в PDAC.
Ингибирование пути Hedgehog изменяет иммунную инфильтрацию в опухолях поджелудочной железы
Поскольку iCAF являются источником воспалительных сигналов (27, 29, 30), мы исследовали последствия увеличения количества iCAF, вызванного ингибированием пути Hedgehog, на иммунный состав.Цитометрия с помощью анализа времени пролета (CyTOF) диссоциированных опухолей KPC позволила оценить множество иммунных популяций (фиг. 5A; дополнительная таблица S1). Мы не наблюдали никаких изменений в общих В-клетках (CD45 + CD19 + ), естественных клетках-киллерах (CD45 + CD161 + ), миелоидных клетках (CD45 + CD11b + ), незрелых гранулоцитарных миелоидных клетках. (Ly6G + Ly6C + ) и макрофаги (CD11b + F4 / 80 + ) после 2-недельного лечения ингибированием пути Hedgehog (рис.5B; Дополнительный рис. S5A). Однако моноцитарные незрелые миелоидные клетки Ly6C + Ly6G — , макрофаги PD-L1 + и макрофаги CD206 + значительно увеличились после обработки LDE225 (фиг. 5B; дополнительный рисунок S5A). Среди различных популяций дендритных клеток только общие CD11b + CD11c + CD206 + CD45 + Lin — F4 / 80 — клетки были значительно активированы (дополнительный рисунок S5B).Общая инфильтрация Т-клеток CD3 + , CD4 + и CD8 + варьировалась в разных образцах, и, хотя мы наблюдали тенденцию к уменьшению количества Т-клеток, существенной разницы не было (дополнительный рисунок S5C). Однако относительное количество субпопуляций Т-клеток, измеренное как процент от общего количества Т-лимфоцитов CD3 + , было изменено, поскольку ингибирование пути Hedgehog значительно уменьшало Т-клетки CD8 + и увеличивало Т-клетки CD4 + и CD25. + CD4 + Т-клетки (потенциальные регуляторные Т-клетки; рис.5С). Соответственно, с помощью ИГХ мы наблюдали меньше CD8 + Т-клеток (фиг. 5D и E) и большее количество регуляторных Т-клеток FOXP3 + (фиг. 5F и G) в 2-недельных опухолях KPC, обработанных LDE225, по сравнению с носителем. -обработанные элементы управления.
Рисунок 5.Двухнедельное ингибирование пути Hedgehog (HH) изменяет иммунную инфильтрацию в опухолях поджелудочной железы. A, Схема 2-недельного лечения мышей KPC с опухолью 20 мг / кг LDE225 или носителя перед анализом CyTOF (вверху). У / С, УЗИ.Таблица панели CyTOF, включающая металлическую метку, мишень антитела и тип клеток, преимущественно экспрессируемых (внизу). См. Дополнительную информацию о антителах в таблице S1. B, Ручное стробирование данных CyTOF для всех миелоидных клеток (CD45 + CD11b + ), макрофагов (CD11b + F4 / 80 + ), PDL1 + макрофагов (F4 / 8042 PD-L1 + ) и макрофаги CD206 + (F4 / 80 + CD206 + ) в 2-недельном носителе- ( n = 7) и обработанном LDE225 ( n = 6) Опухоли KPC.Результаты показывают среднее значение ± SEM. *, P <0,05, непарный тест Стьюдента t . C, Ручное стробирование данных CyTOF для CD8 + , CD4 + и CD4 + CD25 + Т-клеток в процентах от общего количества CD3 + Т-клеток. Результаты показывают среднее значение ± SEM. *, P <0,05; **, P <0,01, непарный тест Стьюдента t . D, Типичный ИГХ CD8A в 2-недельных опухолях KPC, обработанных носителем и LDE225.Вкладыши, увеличения. Масштабная линейка 50 мкм. E, Количественное определение окраски CD8A в 2-недельных опухолях KPC, обработанных носителем ( n = 12) и LDE225 ( n = 11). Результаты показывают среднее значение ± SEM. *, P <0,05, непарный тест Стьюдента t . F, Типичный ИГХ FOXP3 в 2-недельных опухолях KPC, обработанных носителем и LDE225. Вкладыши, увеличения. Масштабная линейка 50 мкм. G, Количественное определение окраски FOXP3 в 2-недельных опухолях KPC, обработанных носителем ( n = 12) и LDE225 ( n = 11).Результаты показывают среднее значение ± SEM. **, P <0,01, непарный тест Стьюдента t . H, Модель, объясняющая роль передачи сигналов Hedgehog и эффекты ингибирования Hedgehog на микросреду PDAC. Секретируемые раком лиганды Hedgehog, такие как SHH и IHH, активируют передачу сигналов Hedgehog в окружающих фибробластах (стрелка), особенно в myCAF (слева). Ингибирование Hedgehog приводит к снижению myCAF и увеличению iCAF, а также к уменьшению Т-лимфоцитов CD8 + и большему количеству регуляторных Т-клеток (справа).
Таким образом, наблюдаемое увеличение iCAFs при ингибировании Hedgehog коррелировало с изменениями иммунной инфильтрации, которые согласуются с более иммуносупрессивным микроокружением рака поджелудочной железы (рис. 5H).
Обсуждение
Отличительной чертой рака поджелудочной железы является обширная стромальная реакция. Среди этих стромальных компонентов гетерогенные подтипы фибробластов потенциально вносят различный вклад в прогрессирование рака поджелудочной железы (24, 25, 27-30, 52, 53). Передача сигналов Hedgehog — ключевой путь, активируемый в фибробластах при раке поджелудочной железы; однако противоречивые генетические и фармакологические исследования на моделях мышей, а также разочаровывающие клинические испытания ингибиторов пути Hedgehog указывают на сложный и потенциально многофакторный вклад пути Hedgehog в прогрессирование рака поджелудочной железы.Здесь мы подтверждаем, что активация пути Hedgehog, о чем свидетельствует экспрессия мишеней Hedgehog, таких как Gli1 , ограничивается компартментом фибробластов. Примечательно, что мы демонстрируем, что передача сигналов Hedgehog обогащена подмножеством myCAF по сравнению с другими подтипами CAF как в мышиных моделях PDAC, так и в человеческом раке поджелудочной железы. Более того, мы показываем, что кратковременное ингибирование пути Hedgehog с помощью LDE225 приводит к истощению myCAF и обогащению iCAF, изменяя состав фибробластов в сторону более фибровоспалительной стромы.
Особенно неприятный вопрос заключается в том, почему различные исследования пришли к разным выводам о роли передачи сигналов Hedgehog при раке поджелудочной железы. Один из возможных ответов заключается в подходах, используемых для нацеливания на путь Hedgehog. Предыдущая работа продемонстрировала ограничивающую опухоль роль передачи сигналов Hedgehog посредством генетического удаления эпителия Shh (22, 23). Однако, как отмечалось ранее (6) и подтверждено здесь, клетки рака поджелудочной железы экспрессируют как Shh , так и Ihh .Более того, генетическая делеция Shh в GEMM PDAC привела к соответствующему и потенциально компенсаторному увеличению экспрессии Ihh (22). Здесь мы демонстрируем, что Ihh KO приводит к уменьшению размера опухоли при имплантации клеток PDAC, которые не производят значительных уровней Shh , и в контексте генетически сенсибилизированного фона (т.е. у Gli1 lacz / + мышей что не хватает одной копии Gli1 ). Вместе эти данные показывают, что нацеливание как на Shh , так и на Ihh необходимо для полного распознавания вклада передачи сигналов Hedgehog в рост рака поджелудочной железы.
Дополнительным объяснением разногласий в этой области может быть использование различных фармакологических ингибиторов пути Hedgehog. В то время как в предыдущих исследованиях использовались Vismodegib (Genentech) и IPI-926 (Infinity Pharmaceuticals; ссылки 19, 22, 23), мы использовали LDE225 (Sonidegib, Novartis; ссылка 48), который все еще проходит клинические испытания на PDAC (NCT02358161). Примечательно, что хотя все три ингибитора нацелены на SMO, эти соединения демонстрируют различную фармакокинетику у мышей (49, 50), что может приводить к различной степени ингибирования пути Hedgehog в строме поджелудочной железы.В подтверждение этого предположения, лечение висмодегибом только частично снизило экспрессию Gli1 в мышиной модели рака поджелудочной железы (23), тогда как лечение LDE225 привело к почти полному прекращению экспрессии Gli1 в обеих моделях ортотопической трансплантации опухолей PDAC и KPC. Важным соображением в предыдущих исследованиях, а также в этой работе является потенциальное влияние различных уровней активности пути Hedgehog на рост опухоли поджелудочной железы. Лиганды Hedgehog наиболее известны как морфогены, где определенные концентрации лиганда Hedgehog вызывают различные клеточные результаты (54).Продолжительность активности пути Hedgehog также важна для определения клеточных ответов (54). Результат передачи сигналов Hedgehog регулируется динамическими и перекрывающимися вкладами различных рецепторов клеточной поверхности (15). Кроме того, белки GLI, эффекторы транскрипции пути Hedgehog, также влияют на рост опухоли поджелудочной железы (45, 55). Наконец, у мышей Gli1 lacz / + передача сигналов Hedgehog системно снижена (32) до образования опухоли, что может объяснить различные результаты, наблюдаемые при дальнейшем ингибировании Hedgehog обработкой LDE225 по сравнению с моделями трансплантации BL / 6J. .В целом, предыдущая работа нашей группы (15) и наши текущие данные подтверждают представление о том, что передача сигналов Hedgehog играет зависимую от дозы и, возможно, временную роль при раке поджелудочной железы с наилучшим результатом с точки зрения ингибирования роста опухоли, когда путь полностью инактивирован, а не частично подавлен.
Здесь мы демонстрируем, что ингибирование пути Hedgehog изменяет соотношение подтипов фибробластов при раке поджелудочной железы путем изменения численности myCAF. Хотя все подтипы CAF, по-видимому, экспрессируют рецепторы, необходимые для ответа на лиганды Hedgehog, мы наблюдали активацию предпочтительного пути Hedgehog в myCAFs.Это наблюдение может быть связано с проксимальным расположением myCAFs в опухоли (27), что позволяет расположить их ближе к источнику лигандов Hedgehog. Это поднимает вопрос о том, может ли генетическое истощение миофибробластов αSMA + (56) привести к несоответствующей активации передачи сигналов Hedgehog в iCAF и apCAF, что может способствовать прогрессированию заболевания.
Наши наблюдения показывают, что вредные эффекты, связанные с долгосрочным ингибированием пути Hedgehog, могут быть не из-за истощения опухолевой ограничивающей популяции myCAF, как предполагалось ранее (22, 42), а вместо этого могут зависеть от обогащения потенциально подтип iCAF, способствующий развитию опухолей.Действительно, предыдущие исследования наших лабораторий показали, что iCAFs способствует развитию опухолей (27, 29, 36, 57). Хотя изменение состава фибробластов, наблюдаемое в течение 2-недельного периода ингибирования Hedgehog, не привело к более агрессивному PDAC, в долгосрочной перспективе оно может вызвать пагубные эффекты, наблюдаемые в предыдущих исследованиях, поскольку iCAF секретируют лиганды, ответственные за такие состояния, как иммуносупрессия и т. кахексия (27, 30, 58–60). В поддержку этого мнения мы обнаружили, что ингибирование Hedgehog приводит к более иммуносупрессивному микроокружению, о чем свидетельствует меньшее количество CD8 T-клеток и большее количество регуляторных T-клеток, что может привести к худшему долгосрочному прогнозу.Примечательно, что мы не можем исключить, что ингибирование пути Hedgehog также может иметь прямое влияние на иммунные клетки, как сообщалось ранее (61). С другой стороны, вредные эффекты, связанные с долгосрочным ингибированием пути Hedgehog, могут быть связаны с потерей хорошо дифференцированного PDAC и увеличением плохо дифференцированного PDAC, которое мы наблюдали после 2-недельного лечения LDE225, что согласуется с предыдущими исследованиями (22, 23). В целом, эффекты ингибирования пути Hedgehog на прогрессирование PDAC могут быть связаны не только с изменениями в конкретных подтипах CAF, но и с тем, как эти изменения влияют на передачу сигналов внутри и между этими клетками и как конкретные сигналы интегрируются с другими компонентами микросреды опухоли.Более глубокое понимание биологии и функций отдельных подтипов CAF, включая исследования, изучающие перекрестные помехи между опухолью / CAF / иммунными клетками, может привести к будущим комбинаторным методам лечения.
Раскрытие информации авторами
Г. Биффи сообщает о грантах от Cancer Research UK и Human Frontier Science Programme во время проведения исследования. Ю. Чжан сообщает о грантах от NIH во время проведения исследования и вне представленной работы. М.А. Андерсон сообщает о других работах от GlaxoSmithKline, Boehringer Ingelheim, Boston Scientific и Olympus, помимо представленных работ.А. Рао сообщает о других грантах от Voxel Analytics, LLC, Agilent Technologies, Genophyll, LLC и Texas A&M IBT, а также о грантах от NIH и ACS помимо представленных работ. Д.А. Тувсон сообщает о грантах от NIH NCI, Lustgarten Foundation и Simons Foundation во время проведения исследования. Доктор Тувсон также сообщает о грантах от ONO, Fibrogen и Merck; другие от Leap Therapeutics, Surface Oncology и Cygnal Therapeutics; а также гранты и другие материалы от Mestag Therapeutics за пределами представленной работы, а также имеет патент в качестве соавтора на «Ингибиторы пути ежа», поданный CRT / CRUK 23 января 2009 г. как US61 / 205 837 (Olive and коллеги, 2009) и PCT US2010 / 021816, подана 22 января 2010 г., выдана CRUK / CRT.Б.Л. Аллен сообщает о грантах от NIH и Мичиганского университета во время проведения исследования. Остальные авторы не сообщили о разглашении информации.
Заявление об ограничении ответственности
Спонсоры не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Вклад авторов
N.G. Стил: Концептуализация, формальный анализ, проверка, исследование, написание — первоначальный черновик, написание — рецензирование и редактирование. г.Biffi: Концептуализация, проверка, исследование, написание — первоначальный черновик, написание — рецензирование и редактирование. S.B. Кемп: Расследование. Ю. Чжан: Расследование. Д. Друйяр: Расследование. Л. Сю: Расследование. Y. Hao: Формальный анализ. T.E. Oni: Исследование, написание – рецензирование и редактирование. Э. Броснан: Расследование. Эляда Э.: Расследование. А. Доши: Расследование. С.Hansma: Формальный анализ, расследование. К. Эспиноза: Расследование. А. Аббас: Расследование. S. The: Расследование. В. Ирисарри-Негрон: Расследование. C.J. Halbrook: Расследование. N.E. Franks: Расследование. M.T. Хоффман: Расследование. К. Браун: Расследование. E.S. Плотник: Расследование. Z.C. Nwosu: Формальный анализ, расследование. К. Джонсон: Формальный анализ. Ф. Лима: Расследование. M.A. Anderson: ресурсов. Ю. Парк: Формальный анализ. H.C. Кроуфорд: ресурсов. C.A. Lyssiotis: ресурсов. T.L. Франкель: Ресурсы, авторский надзор. А. Рао: Формальный анализ. F. Bednar: Ресурсы, курирование данных, формальный анализ, методология, написание – обзор и редактирование. А.А. Длугош: Исследование, написание – рецензирование и редактирование. J.B. Preall: Ресурсы, курирование данных, формальный анализ, методология, написание – обзор и редактирование. Д.А. Tuveson: Концептуализация, ресурсы, надзор, привлечение финансирования, расследование, написание – обзор и редактирование. Б.Л. Аллен: Концептуализация, ресурсы, формальный анализ, надзор, привлечение финансирования, расследование, методология, написание — первоначальный черновик, написание — рецензирование и редактирование. М. Паска ди Мальяно: Концептуализация, ресурсы, формальный анализ, исследование, написание — первоначальный черновик, написание — рецензирование и редактирование.
Благодарности
Результаты, представленные здесь, частично основаны на данных, собранных Сетью исследований атласа генома рака: https: // www.Cance.gov/tcga. Этот проект был поддержан грантами NIH / NCI R01CA151588, R01CA198074 и U01CA224145, а также Американским онкологическим обществом (М. Паска ди Мальяно). Эта работа была поддержана грантом онкологического центра Мичиганского университета (P30CA046592), включая административное приложение (для Х. К. Кроуфорда и М. Паска ди Мальяно). Эта работа была поддержана NIH (R01 DC014428, R01 CA198074 и R01 118751 — B.L. Allen). Исследование, описанное в этой статье, было поддержано грантом поддержки онкологического центра Мичиганского университета (P30 CA046592) за счет использования следующих общих ресурсов онкологического центра: визуализация клеток и тканей.Ф. Беднар финансировался Премией Джоэла Рослина Ассоциации академической хирургии. T.L. Франкель был профинансирован K08CA201581. Исследование, описанное в этой статье, было поддержано наградой NCI под номером P30CA046592 за счет использования следующих общих ресурсов Рогельского онкологического центра: модели трансгенных животных (на имя А.А. Этот проект был поддержан грантами NIH / NCI R01 CA118875 (для L. Syu) и P01 DK062041 и 5P30 CA046592 (для A.A. Dlugosz). С. Кемп был поддержан T32 GM113900. Н.Г. Стил получил поддержку T32 CA009676 и получил награду PF-19-096-01 Американского онкологического общества и стипендию Мичиганского института клинических и здоровых исследований в программе постдокторских переводов.E.S. Карпентер был поддержан премией Американского колледжа гастроэнтерологии за клинические исследования и фондом T32 DK094775. A. Rao и S. The были поддержаны институциональными фондами стартапов Мичиганского университета, подарком от Agilent Technologies, грантом NCI R37 CA214955 и грантом научного сотрудника Американского онкологического общества (RSG-16-005-01). C.J. Halbrook был поддержан наградами K99 CA241357 и P30 DK034933 NIH. Лаборатория Tuveson была поддержана грантом поддержки онкологического центра NIH P30 CA045508 и фондом Lustgarten Foundation, где Д.А. Тувсон — выдающийся ученый и директор лаборатории исследования рака поджелудочной железы, назначенной Фондом Люстгартена. Д.А. Тувсон также поддерживался Фондом Томпсона, лабораторией Колд-Спринг-Харбор и отделением здравоохранения Northwell, Программой донорства тканей Northwell Health, Ассоциацией лабораторий Колд-Спринг-Харбор и Национальным институтом здравоохранения (NIH P30 CA45508, U01 CA210240, R01 CA229699, U01 CA224013, R01 CA188134 и R01 CA1
). Работа поддержана подарком от Фонда Саймонса (552716 Д.А. Тувсон). Дж. Биффи был научным сотрудником программы Human Frontiers Science Program (LT000195 / 2015-L) и EMBO (ALTF 1203-2014) и получил финансовую поддержку от британского фонда исследований рака (A27463), а также программ по раку поджелудочной железы и CMB Кембриджский центр Великобритании по исследованию рака. Э. Эльяда был научным сотрудником программы Human Frontiers Science Program (LT000403 / 2014-L). J.B. Preall был поддержан лабораторией Колд-Спринг-Харбор и организацией Northwell Health Affiliation. Y. Park получил поддержку R50 CA211506. Авторы хотели бы поблагодарить Тришу Тэмсен и Джуди Опп из Мичиганского университета Advanced Genomics Core и Центр закупок тканей Мичиганского университета.Мы также хотели бы поблагодарить Мэтью Кокрана и Терри Вайтмана из Университета в Лаборатории общих ресурсов проточной цитометрии Медицинского центра Рочестера. Мы хотели бы поблагодарить доктора Христа Райта за то, что он поделился моделью мыши iKC (
Ptf1a -Cre ERT ; Kras LSL-G12D / + ). Эта работа была выполнена при поддержке общих ресурсов Мичиганского университета: биостатистика, аналитика и биоинформатика; Проточной цитометрии; Модели трансгенных животных; Тканевая и молекулярная патология; Структура и проверка лекарств; Визуализация клеток и тканей; Экспериментальное облучение; Доклиническая визуализация и компьютерный анализ; Связь в области здравоохранения; Иммунный мониторинг; и фармакокинетика.Эта работа была выполнена при поддержке общих ресурсов лаборатории Колд-Спринг-Харбор, которые поддерживаются грантом поддержки онкологического центра NIH P30 CA045508: общие ресурсы по биоинформатике, проточной цитометрии, визуализации животных и клеток и тканей.Расходы на публикацию этой статьи были частично покрыты за счет оплаты страницы. Таким образом, данная статья должна быть помечена как реклама в соответствии с 18 U.S.C. Раздел 1734 исключительно для указания этого факта.
Сноски
Примечание: Дополнительные данные для этой статьи доступны на сайте Clinical Cancer Research Online (http://clincancerres.aacrjournals.org/).
Clin Cancer Res 2021; 27: 2023–37
- Получено 21 сентября 2020 г.
- Исправление получено 17 ноября 2020 г.