помогите разобрать слова по составу
#2
Гость
ну, и где же все вумановские грамотейки?)))))))))))
#4
#5
обид-корень, еть-суффикс.
#6
е-суффикс
ть-окончание инфинитива
воз-приставка
выс-корень
и-суффикс
ть-окончание инфинитива
но если у вас в учебнике не пишут про инфинитив,то просто окончание))
#8
#9
#10
_Kristen_
обид-корень
е-суффикс
ть-окончание инфинитива
воз-приставка
выс-корень
и-суффикс
ть-окончание инфинитива
но если у вас в учебнике не пишут про инфинитив,то просто окончание))
#11
#12
#13
Издевка_3
_Kristen_обид-корень
е-суффикс
ть-окончание инфинитива
воз-приставка
выс-корень
и-суффикс
ть-окончание инфинитива
но если у вас в учебнике не пишут про инфинитив,то просто окончание))зачем это уточнять?:))
и в школе тоже бывало)предупредила автора
#15
#16
_Kristen_
его всё0таки чаще выделяют просто окончанием))тем более-в школе)
#17
#18
#19
#20
#21
#22
#23
#24
#25
#26
Корнем с дорогой роднится
В сборе приставка таиться
Суффикс-как в слове дневник
Целым же в космос проник
Эксперты Woman. ru
ru
Международный Институт
Психология, психотерапия,…
20 ответов
Владимир Титаренко
Фитнес-нутрициолог
179 ответов
Алёшина Мария
Психолог
101 ответ
Иванова Светлана
Коуч
82 ответа
Максим Сорокин
Практикующий психолог
1 083
Токарь Дарья Анатольевна
Фитнес-тренер
53 ответа
Анна Антончик
Женский психолог
160 ответов
Мария Бурлакова
Психолог
335 ответов
Мария Синяпкина
Психолог-Сексолог
92 ответа
- Кременецкая Мария
Логопед — дефектолог
44 ответа
#27
#28
#29
#30
Непридуманные истории
Меня бесит муж со своими детьми и внуками.
 ..
..1 368 ответов
Мужчина сразу предупредил, что всё имущество записано на детей
1 022 ответа
Такая зарплата — не хочу работать
674 ответа
Ложь длинною в 22 года. Как разрулить?
932 ответа
Ушел муж, 2 месяца депрессия… Как справится, если ты осталась совсем одна?
202 ответа
 ..
..#31
Гость
помогите разобрать слово благодарить по составу
#32
яхОр
Гость
помогите разобрать слово благодарить по составу
благодарить — сложное слово, два корня — «благо» и «дар», окончание — «ить»
#33
#34
#35
#36
цезарь
как разобрать слово отдых по составу? от-приставка?
#37
нина
помогите разобрать по составу слова: артиллерия, галерея, достоинство, интерьер
1) -ия
2)-я
3)-а
4)нулевое.
в 1 и 2 и 4 остальное-это корень. Достоинство: корень-достоин, суффикс-ств. Исторически выделялась приставка до-, корень стои-. Как-то так…
#38
#39
#40
«рыжеватенький»: рыж- — корень; -еват-, -еньк- — суффиксы; -ий — окончание
Новые темы
Проблемы с коллегой
Нет ответов
Друг моего парня
1 ответ
Когда делать тест?
Нет ответов
Ко мне плохо относятся одногруппницы — почему?
Нет ответов
Я в шоке от невестки
10 ответов
#41
Гость
человечек
#43
#44
Гость
Помогите))) озеро
#45
#46
#47
#48
Р╫Р╣Р╥Р╫Р╟Р╧Р╨Р╟
помогите разобрать слова по составу обидеть.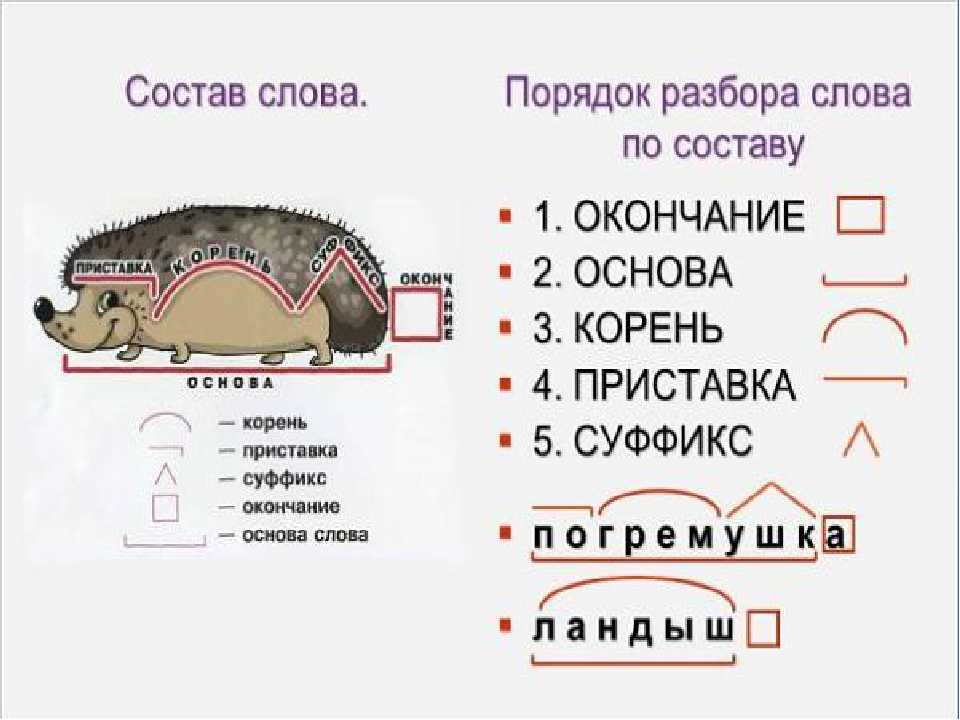 ,возвыситьобид-корень е-суффикс ть- окончание.и слово возвыситьвоз -приставкавыс-кореньи-суффиксть-окончаниетак правильно???
,возвыситьобид-корень е-суффикс ть- окончание.и слово возвыситьвоз -приставкавыс-кореньи-суффиксть-окончаниетак правильно???
#49
#50
Загорится-разобрать слово по составу (морфемный разбор). Слово-ЗАГОРИТСЯ — Знания.site
Последние вопросы
Русский язык
4 минуты назад
Помогите составить диалог СРОООЧНО НАДОРусский язык
29 минут назад
Помогите срочно контрольнаяРусский язык
43 минут назад
Помогите пожалуйста, нужно определить основную мысль текстаРусский язык
48 минут назад
Физика срочно 8 классРусский язык
58 минут назад
Помогите Have / HasРусский язык
1 час назад
Нужна помощь с заданием по русскому языку за 7 класс на тему предлогиРусский язык
1 час назад
Помогите, пожалуйста,Заменить бессоюзные предложения на СППРусский язык
1 час назад
Пожалуйста помогите с русским языком. 7 класс тема предлогиРусский язык
1 час назад
Помогите решить срочно пожалуйстаРусский язык
1 час назад
Помогите сформулировать пжРусский язык
1 час назад
Вольфрам-кобальтовый сплав, вольфрамо-кобальтовый сплав или вольфрамокобальтовый сплавРусский язык
1 час назад
Что значит битый? Битый час, битый деньРусский язык
1 час назад
Пожалуйста решите срочнгРусский язык
1 час назад
Решиье пожалуйста срочншРусский язык
2 часа назад
Как воспринимать фразу «Индустрия плачет ей тебя не хватало» Почему хватало? Ну типо индустрия же женского рода не?
 7 класс тема предлоги
7 класс тема предлогиВсе предметы
Выберите язык и регион
English
United States
Polski
Polska
Português
Brasil
English
India
Türkçe
Türkiye
English
Philippines
Español
España
Bahasa Indonesia
Indonesia
Русский
Россия
How much to ban the user?
1 hour 1 day 100 years
функциональное программирование — Python + дизассемблирование составных функций
Задавать вопрос
спросил
Изменено 2 года, 3 месяца назад
Просмотрено 100 раз
Я представляю своим студентам некоторые аспекты функционального программирования. Мы используем python в качестве основного языка. Для меня создание функций — один из основных аспектов функционального программирования. Чтобы проиллюстрировать это, я предложил следующий пример, не что иное, как классический.
Мы используем python в качестве основного языка. Для меня создание функций — один из основных аспектов функционального программирования. Чтобы проиллюстрировать это, я предложил следующий пример, не что иное, как классический.
импорт дис
защита f(x):
вернуть 2*х+1
определение г (х):
вернуть х**2
деф комп(веселье1, удовольствие2):
вернуть лямбда x:fun1(fun2(x))
дис.дис (ф)
дис.дис (г)
дис.дис (комп (ф, г))
Мне просто интересно, есть ли способ вернуть выражение для comp(f,g) с модулем dis . Я понял, что есть параметры для настройки уровня рекурсивных вызовов каким-то образом, но я не тратил достаточно времени на изучение. Я также наткнулся на модуль ast , который на первый взгляд выглядел немного утомительно.
Итак, я подумал, что это вопрос для Stackoverflow: задолго до любой идеи символического вычисления, есть ли способ получить dis описать comp(f,g) как lambda x:2*x**2+ 1 или ast отображение абстрактного синтаксического дерева для этого выражения?
Спасибо за любой совет.
- python
- функциональное программирование
- абстрактное синтаксическое дерево
- дизассемблирование
- функция-композиция
Насколько мне известно, то, что вы хотите, невозможно просто при обычном использовании пакетов ast или dis . Это связано с тем, что что касается AST или байт-кода comp , реализация вызываемых функций непочтительна, просто нужно знать, чтобы загружать функции, на которые ссылаются, и вызывать их.
Если вы хотите копнуть немного глубже, вы можете смоделировать подстановочные вызовы для их реализации просто для моделирования композиции функций, если вы готовы пожертвовать точностью того, как на самом деле работает выполнение, создавая и манипулируя AST, заменяя звонки в течение comp с реализацией функции, которую они вызывают.
Я собрал небольшой пример того, что вам нужно сделать здесь https://gist.github.com/buckley-w-david/e9c67cca7070282ddb5ab8d37de06f4e (требуется python 3. 9+, и есть много предположений о структура программы, в которой он должен работать), но это немного непрозрачно, если вы не знакомы с работой с AST в Python.
9+, и есть много предположений о структура программы, в которой он должен работать), но это немного непрозрачно, если вы не знакомы с работой с AST в Python.
2
Зарегистрируйтесь или войдите в систему
Зарегистрируйтесь с помощью Google
Зарегистрироваться через Facebook
Зарегистрируйтесь, используя электронную почту и пароль
Опубликовать как гость
Электронная почта
Требуется, но никогда не отображается
Опубликовать как гость
Электронная почта
Требуется, но не отображается
Как разбирают очаговые спайки?
Перейти к содержимомуПоиск:
Как разобрать очаговые спайки?steve2017-12-26T14:16:20+08:30
Разборка имеет решающее значение во время распространения клеток и движения вперед, чтобы позволить ЖК расширяться наружу к периферии клетки [1]. Разборка спаек может происходить в результате ретракции в задней части клетки после созревания спаек или в основании выпячиваний при обороте. Это в первую очередь опосредовано событиями фосфорилирования тирозина и изменениями в натяжении цитоскелета [2], которые контролируются такими регуляторами, как кальпаин [3] и микротрубочки [4].
Разборка спаек может происходить в результате ретракции в задней части клетки после созревания спаек или в основании выпячиваний при обороте. Это в первую очередь опосредовано событиями фосфорилирования тирозина и изменениями в натяжении цитоскелета [2], которые контролируются такими регуляторами, как кальпаин [3] и микротрубочки [4].
Разборка, опосредованная ретракцией, координируется асимметрично, чтобы поддерживать направленную миграцию, и обычно происходит в задней части клетки (rev. [5]). Это связано со «скольжением» спаек по мере продвижения края внутрь [6]. Скользящее движение, по-видимому, зависит от Rho/myosin II, однако механизм до сих пор не ясен (см. обзор в [7]). Исследование, сообщающее о дифференциальной регуляции скорости оборота отдельных белков зубного налета с помощью механической силы [8]. Это указывает на то, что их размер и молекулярный состав изменяются при изменении стресса и, таким образом, могут инициировать разборку ЖК. Известно, что крупные белковые агрегаты покидают адгезивный комплекс и диспергируются по мере удаления от места адгезии [9]. ]. Было замечено, что кластеры, которые расходятся от скользящих спаек, движутся центростремительно вдоль боковых краев клетки, сливаются с другими и могут быть стабильными в течение 30 минут до распада [6].
]. Было замечено, что кластеры, которые расходятся от скользящих спаек, движутся центростремительно вдоль боковых краев клетки, сливаются с другими и могут быть стабильными в течение 30 минут до распада [6].
Известно, что микротрубочки (МТ) отрицательно регулируют сократимость клеток и, следовательно, усиливают разборку (обзор в [10]). Было обнаружено, что они нацелены на сайты адгезии [11] и в конечном итоге вызывают скольжение или дестабилизацию этих структур, релаксацию актиновых пучков на концах, связанных с очагами адгезии [12]. MT могут стимулировать это высоко локализованное событие либо через тирозинкиназу Arg, которая ингибирует Rho [13], либо посредством взаимодействия FAK-опосредованной дестабилизации и динамин-опосредованного/клатрин-опосредованного эндоцитоза интегринов [14][15] (обзор в [14]. 5]). Кинезин также участвует в регулируемой МТ фокальной доставке компонентов, которые замедляют рост спаек и способствуют разборке [16]. Также существуют доказательства других механизмов MT-опосредованной разборки, где деполимеризация MT способствует Rho-зависимой сократимости за счет высвобождения связанного GEF, что может вызывать нестабильность адгезии [17][18].
С др. стороны, на границе ламеллиподий-ламеллюм оборот зарождающихся адгезивных структур происходит во время миграции и зависит от сил, генерируемых деполимеризацией и реорганизацией актина [19][20]. Оборачиваемость этих структур играет важную роль в поддержании определенной границы ламеллиподия-пластинки через циклы выпячивания/сокращения и связывания актина в пластинке [21].
Сигнальный путь FAK-Src, по-видимому, играет критическую роль в опосредовании оборота адгезии на клеточном фронте [22][23]. Он передается посредством фосфорилирования киназы легкой цепи миозина (MLCK) киназой ERK/MAP, что увеличивает сократительную способность актомиозина [24]. При высокой силе высокая плотность интегрина приводит к менее стабильным адгезивным связям интегрин-ECM [25]. Таким образом, в результате разрыва актиновых связей высвобождаются рецепторы [6][26]. С другой стороны, оборот может усиливаться за счет временного подавления Rho-активности, опосредованного FAK [27][28]. Протеолиз ключевых компонентов адгезии, таких как талин [29], β3-интегрин [30] и FAK [31] с помощью кальпаина также участвуют в этой функции.
Было показано, что рециркуляция индивидуальных интегринов посредством активного транспорта к переднему краю ресенсибилизирует интегрины для связывания лиганда [32] (rev. [33]). Таким образом, скоординированная разборка, переработка и направленная транспортировка поддерживают баланс сборки и разборки спаек, необходимый для постоянной миграции.
Просмотреть все
Последние находки
Ссылки
- Партридж М.А. и Маркантонио Э.Э. Инициация прикрепления и образование зрелых фокальных спаек интегринсодержащими филоподиями при распространении клеток. Мол. биол. Сотовый 2006; 17(10):4237-48. [PMID: 16855018]
- Кроули Э. и Хорвиц А.Ф. Фосфорилирование тирозина и напряжение цитоскелета регулируют высвобождение спаек фибробластов. Дж. Клеточная биология. 1995 год; 131(2):525-37. [PMID: 7593176]
- Franco SJ и Huttenlocher A. Регуляция миграции клеток: кальпаины делают разрез. Дж. Селл. науч. 2005 г. ; 118 (часть 17): 3829-38. [PMID: 16129881]
- Остин Дж. Х. и Карсен Г. М. Письмо: Острая ревматическая гематома аорты. Дж. Торак. Кардиовас. Surg. 1976 год; 71(2):321. [PMID: 1246156]
- Broussard JA, Webb DJ, and Kaverina I. Разборка асимметричной фокальной адгезии в подвижных клетках. Курс. мнение Клеточная биол. 2008 г.; 20(1):85-90. [PMID: 18083360]
- Лаукайтис С.М., Уэбб Д.Дж., Донаис К. и Хорвиц А.Ф. Дифференциальная динамика альфа-5-интегрина, паксилина и альфа-актинина при формировании и разборке спаек в мигрирующих клетках. Дж. Клеточная биология. 2001 г.; 153(7):1427-40. [PMID: 11425873]
- Парсонс Дж. Т., Хорвиц А. Р. и Шварц М. А. Клеточная адгезия: интеграция динамики цитоскелета и клеточного напряжения. Нац. Преподобный Мол. Клеточная биол. 2010 г.; 11(9):633-43. [PMID: 20729930]
- Wolfenson H, Bershadsky A, Henis YI и Geiger B. Генерируемое актомиозином напряжение контролирует молекулярную кинетику фокальных спаек. Дж. Селл. науч. 2011 г.; 124 (часть 9): 1425-32. [PMID: 21486952]
- Дигман М.А., Уайзман П.В., Чой С., Хорвиц А.Р. и Граттон Э. Стехиометрия молекулярных комплексов при адгезии в живых клетках. проц. Натл. акад. науч. США 2009 г.; 106(7):2170-5. [PMID: 19168634]
- Малый Дж. В., Гейгер Б., Каверина И., Бершадский А. Как микротрубочки направляют мигрирующие клетки? Нац. Преподобный Мол. Клеточная биол. 2002 г.; 3(12):957-64. [PMID: 12461561]
- Каверина И., Роттнер К. и Смолл СП. Нацеливание, захват и стабилизация микротрубочек при ранних очаговых спайках. Дж. Клеточная биология. 1998 год; 142(1):181-90. [PMID: 9660872]
- Каверина И., Крылышкина О., Малый СП. Нацеливание микротрубочек на контакты субстрата способствует их релаксации и диссоциации. Дж. Клеточная биология. 1999; 146(5):1033-44. [PMID: 10477757]
- Пикок Дж. Г., Миллер А. Л., Брэдли В. Д., Родригес О. С., Уэбб Д. Д. и Колеске А. Дж. Связанный с Abl ген тирозинкиназы действует через p190RhoGAP, ингибируя сократительную способность актомиозина и регулируя динамику фокальной адгезии при адгезии к фибронектину. Мол. биол. Сотовый 2007; 18(10):3860-72. [PMID: 17652459]
- Эзратти Э.Дж., Берто С., Маркантонио Э.Э. и Гундерсен Г.Г. Клатрин опосредует эндоцитоз интегрина для разборки фокальной адгезии в мигрирующих клетках. Дж. Клеточная биология. 2009 г.; 187(5):733-47. [PMID: 19951918]
- Эзратти Э.Дж., Партридж М.А. и Гундерсен Г.Г. Разборка фокальной адгезии, индуцированная микротрубочками, опосредована динамином и киназой фокальной адгезии. Нац. Клеточная биол. 2005 г.; 7(6):581-90. [PMID: 15895076]
- Крылышкина О., Каверина И., Краневиттер В., Штеффен В., Алонсо М.С., Кросс Р.А. и Смолл СП. Для модуляции динамики адгезии субстрата посредством нацеливания на микротрубочки требуется кинезин-1. Дж. Клеточная биология. 2002 г.; 156(2):349-59. [PMID: 11807097]
- van Horck FP, Ahmadian MR, Haeusler LC, Moolenaar WH и Kranenburg O. Характеристика p190RhoGEF, RhoA-специфический фактор обмена гуаниновых нуклеотидов, взаимодействующий с микротрубочками. Дж. Биол. хим. 2000 г.; 276(7):4948-56. [PMID: 11058585]
- Крендель М., Зенке Ф.Т. и Бокоч Г.М. Фактор нуклеотидного обмена GEF-h2 опосредует перекрестные помехи между микротрубочками и актиновым цитоскелетом. Нац. Клеточная биол. 2002 г.; 4(4):294-301. [PMID: 11912491]
- Александрова А.Ю., Арнольд К., Шауб С., Васильев Ю.М., Мейстер Дж., Бершадский А.Д., Верховский А.Б. Сравнительная динамика ретроградного тока актина и фокальных спаек: образование зарождающихся спаек запускает переход от быстрого тока к медленному. ПЛОС ОДИН 2008; 3(9):e3234. [PMID: 18800171]
- Шемеш Т., Верховский А.Б., Свиткина Т.М., Бершадский А.Д., Козлов М.М. Роль фокальных спаек и механических напряжений в формировании и прогрессировании границы между ламеллиподиями и ламеллами [исправлено]. Биофиз. Дж. 2009; 97(5):1254-64. [PMID: 19720013]
- Barthel W, and Markwardt F. Агрегация тромбоцитов адреналином и его поглощение. Биохим. Фармакол. 1975 год; 24(20):1903-4. [PMID: 20]
- Webb DJ, Donais K, Whitmore LA, Thomas SM, Turner CE, Parsons JT и Horwitz AF. Передача сигналов FAK-Src через паксиллин, ERK и MLCK регулирует разборку адгезии. Нац. Клеточная биол. 2004 г.; 6(2):154-61. [PMID: 14743221]
- Оуэн К.А., Пиксли Ф.Дж., Томас К.С., Висенте-Мансанарес М., Рэй Б.Дж., Хорвиц А.Ф., Парсонс Дж.Т., Беггс Х.Е., Стэнли Э.Р. и Бутон А.Х. Регуляция персистенции ламеллиподов, оборота адгезии и подвижности в макрофагах с помощью киназы фокальной адгезии. Дж. Клеточная биология. 2007 г.; 179(6):1275-87. [PMID: 18070912]
- Клемке Р.Л., Кай С., Джаннини А.Л., Галлахер П.Дж., де Ланеролл П. и Череш Д.А. Регуляция подвижности клеток митоген-активируемой протеинкиназой. Дж. Клеточная биология. 1997 год; 137(2):481-92. [PMID: 9128257]
- Ballestrem C, Hinz B, Imhof BA и Wehrle-Haller B. Маршировать впереди и тащиться сзади: дифференциальный оборот alphaVbeta3-интегрина регулирует поведение фокальной адгезии. Дж. Клеточная биология. 2001 г.; 155(7):1319-32. [PMID: 11756480]
- Палечек С.П., Хаттенлохер А., Хорвиц А.Ф. и Лауффенбургер Д. А. Физическая и биохимическая регуляция высвобождения интегрина при заднем отслоении мигрирующих клеток. Дж. Селл. науч. 1998 год; 111 (Пт 7): 929-40. [PMID: 9490637]
- Ren XD, Kiosses WB, Sieg DJ, Otey CA, Schlaepfer DD и Schwartz MA. Киназа фокальной адгезии подавляет активность Rho, способствуя обороту фокальной адгезии. Дж. Селл. науч. 2000 г.; 113 (Пт 20): 3673-8. [PMID: 11017882]
- Schober M, Raghavan S, Nikolova M, Polak L, Pasolli HA, Beggs HE, Reichardt LF и Fuchs E. Киназа фокальной адгезии модулирует передачу сигналов напряжения для контроля динамики актина и фокальной адгезии. Дж. Клеточная биология. 2007 г.; 176(5):667-80. [PMID: 17325207]
- Franco SJ, Rodgers MA, Perrin BJ, Han J, Bennin DA, Critchley DR и Huttenlocher A. Опосредованный кальпаином протеолиз талина регулирует динамику адгезии. Нац. Клеточная биол. 2004 г.; 6(10):977-83. [PMID: 15448700]
- Флеварис П., Стоянович А., Гонг Х., Чишти А., Уэлч Э. и Ду Х. Молекулярный переключатель, контролирующий распространение и сокращение клеток.
 ; 118 (часть 17): 3829-38. [PMID: 16129881]
; 118 (часть 17): 3829-38. [PMID: 16129881] 2011 г.; 124 (часть 9): 1425-32. [PMID: 21486952]
2011 г.; 124 (часть 9): 1425-32. [PMID: 21486952]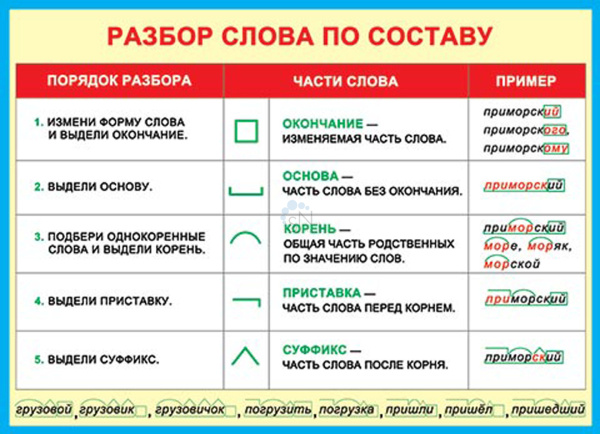 Мол. биол. Сотовый 2007; 18(10):3860-72. [PMID: 17652459]
Мол. биол. Сотовый 2007; 18(10):3860-72. [PMID: 17652459]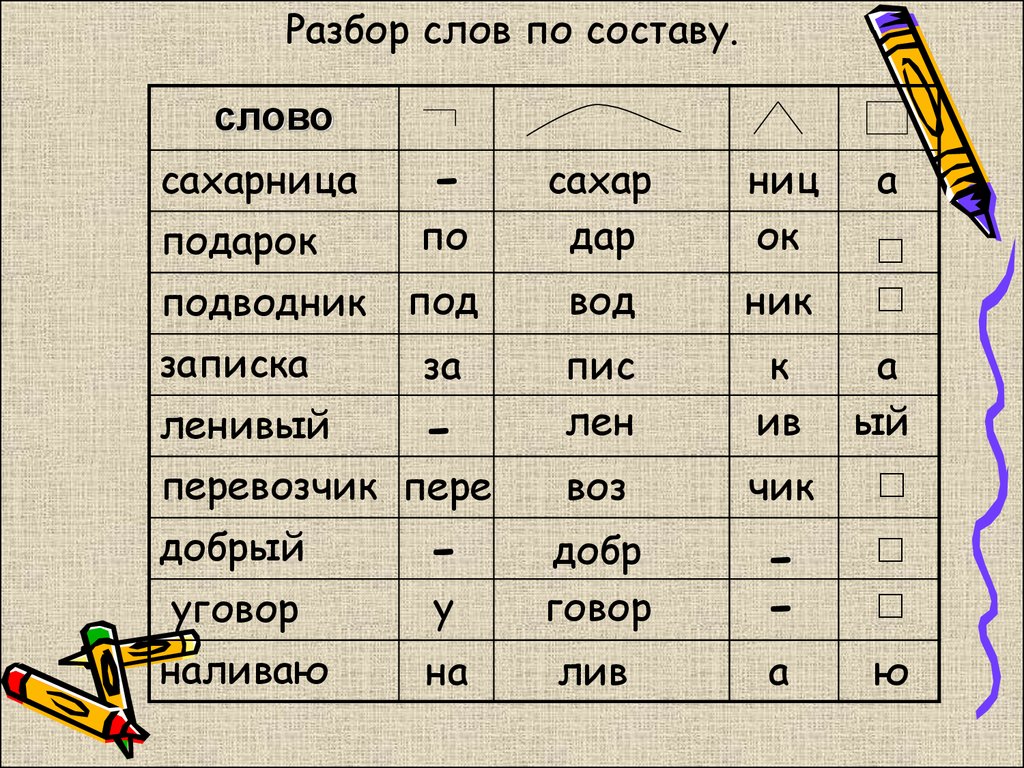 2000 г.; 276(7):4948-56. [PMID: 11058585]
2000 г.; 276(7):4948-56. [PMID: 11058585]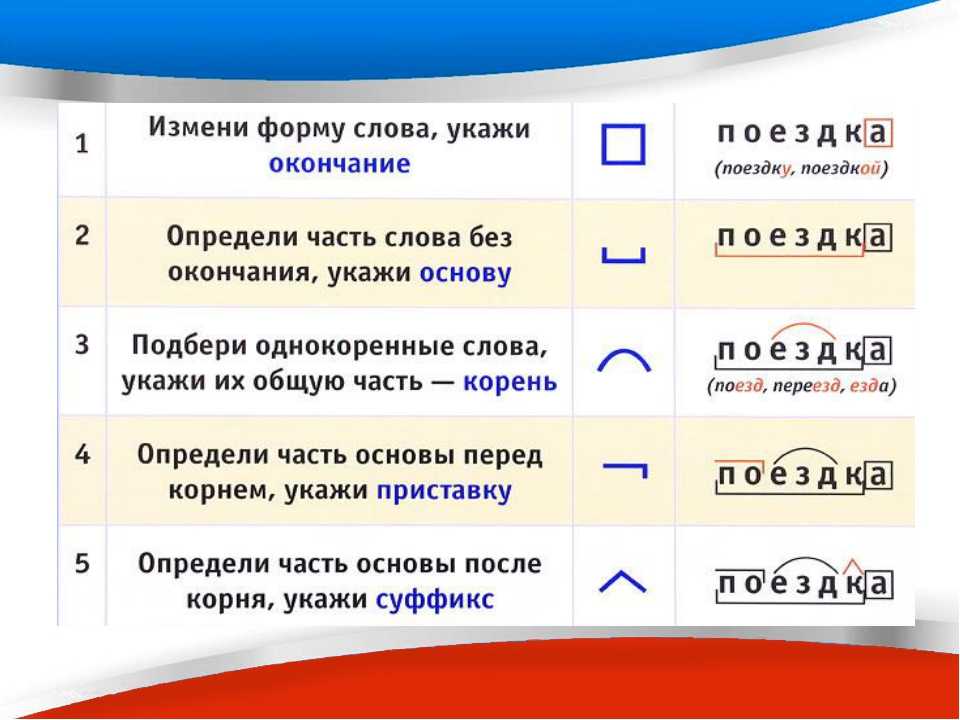 Передача сигналов FAK-Src через паксиллин, ERK и MLCK регулирует разборку адгезии. Нац. Клеточная биол. 2004 г.; 6(2):154-61. [PMID: 14743221]
Передача сигналов FAK-Src через паксиллин, ERK и MLCK регулирует разборку адгезии. Нац. Клеточная биол. 2004 г.; 6(2):154-61. [PMID: 14743221] А. Физическая и биохимическая регуляция высвобождения интегрина при заднем отслоении мигрирующих клеток. Дж. Селл. науч. 1998 год; 111 (Пт 7): 929-40. [PMID: 9490637]
А. Физическая и биохимическая регуляция высвобождения интегрина при заднем отслоении мигрирующих клеток. Дж. Селл. науч. 1998 год; 111 (Пт 7): 929-40. [PMID: 9490637]