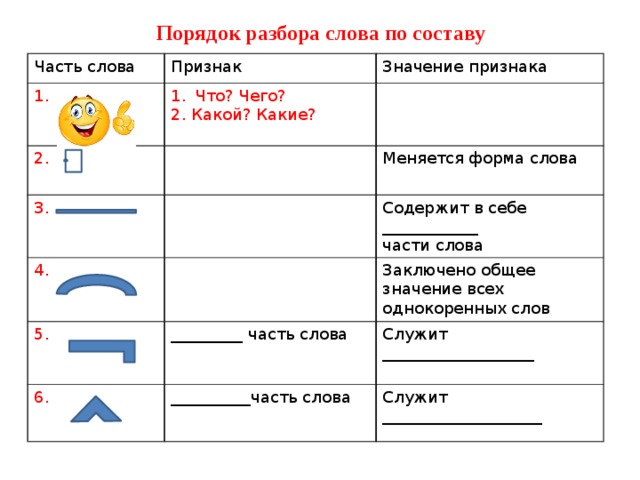
Разбор слова по составу (морфемный анализ) — РОСТОВСКИЙ ЦЕНТР ПОМОЩИ ДЕТЯМ № 7
Содержание
Урок русского языка на тему :Разбор слова по составу (2 класс)
Сквозная тема:
Моя школа.
Школа:
Дата: «__5__»____декабря2018г.
ФИО учителя: Наурзалиева Б Г
Класс: 2 «_б___» класс.
Количество присутствующих: 4
отсутствующих:
Лексическая тема:
История школьных вещей.
Тема урока:
Разбор слова по составу.
Цели обучения, которые достигаются на данном уроке (ссылка на учебную программу):
2.1.1.1 определять с помощью учителя опорные слова, фиксировать их; отвечать на закрытые вопросы.
2.2.3.1 формулировать вопросы с опорой на ключевые слова, отвечать на вопросы по содержанию прочитанного
2.3.8.2 определять значимые части слова, выделять корень в слове и подбирать однокоренные слова
Развитие
навыков:
1.
1 Понимание содержания информации/ сообщения
2.3 Формулирование вопросов и ответов
3.8 Соблюдение грамматических норм.
Критерии успеха (Предполагаемый результат):
Все учащиеся смогут:
Узнать об алгоритме разбора слов по составу.
Узнать о происхождении школьных вещей.
Большинство учащихся смогут:
Объяснить смысл пословицы.
Ответить на вопросы теста.
Проверить правильность выполнения теста.
Развивать логическое мышление.
Выполнять разбор слов по составу по алгоритму.
Находить и разбирать однокоренные слова по составу.
Находить в дополнительных источниках информацию по лексической теме.
Определять в прочитанном тексте основную мысль текста, идейный замысел автора.
Вставлять пропущенные буквы в пословицы.
Некоторые учащиеся смогут:
Придумать загадку о книге.
Языковая
цель
Основные термины и словосочетания:
Состав слова, части слова, приставка, корень, суффикс, окончание, одноко-
ренные слова, родственные слова, пословица, вопрос, текст, рассказ, стихо-
творение, загадка, предложение, упражнение, ответ, пословица.
Используемый язык для диалога/письма на уроке:
Вопрос для обсуждения:
Какие правила обращения с книгой ты знаешь?
Привитие
ценностей
Ценности, основанные на национальной идее «Мәңгілік ел»: казахстанский патриотизм и гражданская ответственность; уважение; сотрудничество; труд и творчество; открытость; образование в течение всей жизни.
Межпредметные
связи
– литература
– самопознание,математика
Навыки
использованияИКТ
На данном уроке учащиеся используют ИКТ
Предварительные
знания
Звуки и буквы, гласные и согласные звуки, печатные и рукописные буквы,алфавитные названия букв, слог, язык – средство человечес.общения,высказывание, текст, языковые и неязыковые средства общения, устная и письменная речь, предложение, прописная буква в начале предложения и в именах собственных, сила голоса и темп речи, алфавитный порядок слов,перенос слов, ударные и безударные гласные, мягкий знак на конце и в середине слов, твердые и мягкие согласные, правописание жи-ши, ча-ща, чу-щу, чк, чн, нщ, рщ, шн, звонкие и глухие согласные, звук [й], разделительный Ь, состав слова, окончание, корень, приставка, суффикс, родственные и однокоренные слова.
Ход урока
Этапы урока
Запланированная деятельность на уроке
Ресурсы
Начало урока
0-4
Создание положительного эмоционального настроя.
— Я рада видеть всех вас на уроке русского языка. А для успешной работы нам сегодня понадобятся: -внимательные глазки, -чуткие ушки, -ловкие ручки, -сообразительные головы. И , конечно же, хорошее настроение! Желаю вам успехов в работе на уроке!
Сегодня мы с вами будем настоящими исследователями. Ребята, а кто такой исследователь? Какими качествами он должен обладать?
( иметь зоркий глаз, быть внимательным, уметь слушать и слышать.). А вот что мы будем исследовать, докажем в ходе нашего урока. Если мы с вами исследователи, то наш кабинет «научная лаборатория». Исследования записываются в научных дневниках. Научный дневник это наши рабочие тетради. Открыли рабочие тетради, записали дату исследования.
5-10 мин
Актуализация знаний. Целеполагание.
Целеполагание.
Цель: ввод ситуации для открытия и постановки целей
урока.
2.-Начнем урок с игры «Четвертый лишний». Перед вами на доске слова: ПРИСТАВКА, СУФФИКС, ГЛАГОЛ, ОКОНЧАНИЕ.
Найдите лишнее слово и объясните свой выбор.
-Так чем же мы будем с вами сегодня заниматься? (находить части слова и разбирать их по составу.)
— Это новая тема или продолжение вчерашней? (Продолжение)
Открывается запись темы урока.
3.. Постановка цели урока
— Как вы думаете, какова цель нашего урока?
(Повторить всё о частях слова и учиться правильно разбирать слова по составу).Что такое корень, окончание, приставка,суффикс?
Найдите лишнее слово и объясните свой выбор.
Критерии успеха
Учащиеся с помощью учителя формулируют тему урока.
Середина урока
11-20 мин
Работа над темой урока.
Цель: усвоение содержания темы.
4.–Я читаю загадку, а вы должны записать отгадку и подобрать однокоренные слова. Выделите корень.
— Кто рыбачит на реке.С тонкой удочкой в руке?
(Рыбак, рыба, рыбалка, рыбачит,…)
— Кто хлопочет с поварёшкой,
Варит вкусный суп с картошкой?
(Повар, сварил, варенье, …)
– Что общего во всех этих словах? (корень) .
Продолжаем исследование.
5.Внимание! Перед вами слова которые рассыпались на части и эти части перепутались. Ваша задача собрать их так, чтобы получились слова. Записать слова и выделить части слова.
-Восстановите нужный порядок следования морфем в каждом слове, и запишите получившиеся слова.
1) к, дар, и, по 2) к, гул, про, а
3) к, а,рас,крас 4) езд, по,а,к
Какие части слова помогли образовать данные слова? (Приставка и суффикс)
— Дайте определение этим частям слова? (Приставка – часть слова — которая стоит перед корнем и служит для образования новых слов. Суффикс – часть слова, которая стоит после корня и служит для образования новых слов)
Оцениваем свою работу смайликом . ( Самооценивание)
( Самооценивание)
6.- Посмотрите внимательно на экран. Перед вами пословица. Прочитайте её.
Без русск… язык… не сколотишь сапог… .
-Получилась у вас пословица? А что нужно сделать, чтобы получился правильный вариант? Какая морфема необходима нам, чтобы связать слова в предложении? (окончание).А как вы понимаете смысл данной пословицы?А теперь скажите, что же такое окончание и для чего оно служит? – Итак, какие морфемы мы с вами повторили?(приставка, корень, суффикс, окончание) .Теперь подведем итог и составим кластер.
7. Кластер «Состав слова»
Загадки
Оцениваем свою работу смайликом .( Самооценивание)
21-22 мин
(Д, К)
8. Минутка отдыха. Гимнастика для глаз.
Видеоролик.
23-26 мин
Работа по теме урока.
Цель: выполнение практических заданий.
10. Работа по карточкам.
— А теперь, ребята, я предлагаю потрудиться самостоятельно каждому из вас. (У каждого ребёнка карточка на парте индивидуальные карточки трёх цветов по степени трудности: зелёные – сильным учащимся; жёлтые – средним учащимся; красные – слабым учащимся. — Ребята, внимательно прочитайте каждый свою карточку и выполните задания.
(У каждого ребёнка карточка на парте индивидуальные карточки трёх цветов по степени трудности: зелёные – сильным учащимся; жёлтые – средним учащимся; красные – слабым учащимся. — Ребята, внимательно прочитайте каждый свою карточку и выполните задания.
Зеленая карточка. Прочитай, озаглавь текст.
Наиди слова с приставкой, наиди однокоренные слова.
Пришла зима.Первые снежинки закружились в воздухе. Полетела снежная пыль.
Жёлтая карточка.Разбери слова по составу.
Зимний, кормушка, закружила.
Красная карточка. Выделите корень слова.
Снеговик, снег, снегурочка.
11. Групповая работа и защита .«Редактор».
Восстанови текст, добавив однокоренные слова. Выдели корень в словах.
У меня встали _________. Ещё ________ назад ходили. А сейчас______ стрелка стоит неподвижно. Нужно, чтобы их починил ________.
Слова для справок: часовщик, час, часовая , часы
Учебник
Дифференцированная работа.
Восстанови текст, добавив однокоренные слова. Выдели корень в словах.
27-28 мин
(Д, К) Динамическая пауза.
По дорожке шли, шли,
Много камешков нашли.
Присели, собрали
И дальше пошли.
Е. Чудиловская
29-35 мин
Работа по развитию речи.
Цель: развитие связной речи.
Работа по учебнику. Самостоятельная работа в тетради.Упр.22
Учебник
Критерии успеха
Ученики составляют загадку о книге, используя рифмы, данные в учебнике (индивидуальная карточка).
Можно предложить некоторым учащимся самим подобрать загадки о книге, используя разные источники информации, или сочинить загадку самостоятельно.
Выборочная проверка.
Конец урока
36-37 мин
(Д, К) Пальчиковая физминутка.
Дружно пальчики сгибаем,
Крепко кулачки сжимаем.
Раз, два, три, четыре, пять –
Начинаем разгибать.
(Вытянуть руки вперед, сжать пальцы в кулачки как можно сильнее, а затем расслабить и разжать.)
О. Гуреева
38-40
Работа в тетради «Что я знаю и умею»
Цель: оценка уровня усвоения навыка по теме.
Итог урока. Рефлексия.Чтобы разобрать слово по составу, нужно … .
12. Обратная связь. « Лестница.»
Тетрадь «Что я знаю и умею»
Критерии успеха
— Что нового вы узнали на уроке?
— Задайте друг другу вопросы по теме урока .
Что мы открыли на уроке? Какое открытие сделали? (Алгоритм разбора состава слова) С помощью стихотворения вспомним порядок разбора слова по составу.
Слово по составу верно разбирай:
Первым, окончание всегда выделяй,
На основу внимательно смотри,
Корень, поскорее, ты у нее найди,
Приставку и суффикс в конце определи.
Дифференциация
Оценивание
Здоровье и соблюдение техники безопасности
Дети с высоким уровнем мотивации создают визитки и вместе с учителем помогают ученикам с низкой мотивацией
Формативное оценивание.
Самооценивание в тетради «Что я знаю и умею».
Взаимооценивание при работе в паре, группе, классом.
Результаты наблюдения учителем качества ответов учащихся на уроке.
Определение уровня усвоения навыка по теме (тетрадь «Что я знаю и умею»).
Динамическая пауза.
по дорожке шли, шли,
Много камешков нашли
.Присели, собралиИ дальше пошли.
Е. Чудиловская
Обл спец школа –интернат № 3
Открытый урок по русскому языку
Тема : Разбор слова по составу
2 « Б» класс
Провела : Наурзалиева Б Г
Атырау 2019г
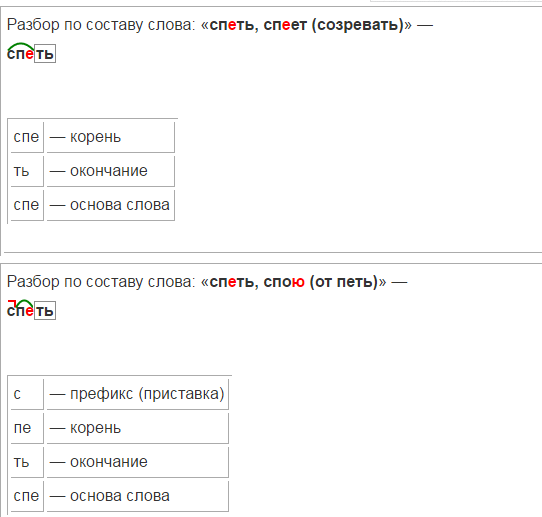
Очень простая инструкция для разбора слова по составу — Морфемный разбор в помощь второклашке | Блог КУМОНомамы
Разбор слова по составу — это очень просто, элементарщина. Частей слова ведь всего 5 (даже почти 4): приставка, корень, суффикс, окончание. А всё вместе, кроме окончания — это основа.
Чего тут сложного, казалось бы.
Проблема в том, что в школе все эти части слова учат и отрабатывают по-отдельности. То суффиксы отдельно ищем, то окончания… И у ребенка не складывается общая картинка — что из себя представляет состав слова. Ему начинает казаться, что это — эдакая игра в угадайку: угадай суффикс, угадай приставку, угадай часть речи.
Интересно, что даже если ребенок хорошо разбирается в частях речи, задания типа «найди суффикс» могут поставить его в тупик, потому что, в отличие от корня, суффикс — это «бессмысленный набор буковок». Пойди угадай, какие именно буковки относятся к этому «треугольничку».
В этой статье предложу схему разбора по составу, применяя которую ребенок всегда — независимо от типа задания — сможет легко разбирать по составу все части речи.
Что нужно знать о частях слова, прежде чем делать разбор
Самое главное — нужно запомнить, что части слова не существуют в изоляции: они все связаны друг с другом. Ну конечно — слово-то одно. Поэтому…
Ну конечно — слово-то одно. Поэтому…
Когда-нибудь ваш второклашка наберется солидного опыта в выявлении любых суффиксов с первого взгляда.
А пока — даже если в задании написано «найди суффикс», для корректного определения этого суффикса нужно сделать разбор полностью. Сначала найди все части слова, а потом среди них вычислить — и уже отметить в тетради только суффикс.
В ближайшие дни я выпущу отдельную статью именно про суффиксы.
Здесь же замечу, что, конечно, не нужно полностью разбирать слово по составу, если ваш второклашка безошибочно узнал суффикс «-еньк-«, например, и ни с чем его никогда не перепутает.
Просто знания о конкретных суффиксах у второклашек редко возникают сами по себе — в связи с этим и возникла рекомендация про полный разбор.
Итак, договорились: для того, чтобы корректно выделить какую-то часть речи, нужно разобрать по составу всё слово.
Теперь двигаемся по схеме.
1. Находим корень
Находим корень
Сначала найдем самое главное — и самое простое.
Чтобы найди корень, нужно придумать похожие слова.
Почитайте эту мою статью про однокоренные слова и формы слова — и обратите внимание: для поиска корня нам нужны НЕ ФОРМЫ одного слова, а именно ОДНОКОРЕННЫЕ слова.
Напр., разбирая слово «подарок» — для поиска корня нам нужно НЕ «подарку, подарком», а «дар, дарить, задарить, подарить«.
«Ударить». Лол 🙂 Поменяли приставку — и вуа-ля 🙂
Реальная история про то, как НЕ НАДО искать однокоренные — расскажу в отдельной статье про приставки.
Итак, подбираем много похожих (однокоренных) слов и внимательно слушаем, что у этих слов изменяется, а что — остается неизменным.
Важно, что это «неизменное» должно иметь смысл — то есть быть не набором абстрактных буковок, а что-то значить.
Напр., в нашем примере с «подарком» у всех подобранных нами слов остается неизменной часть «дар». А мы, слыша эту часть, как-то понимаем, что это не абракадабра, а что-то, связанное с бесплатным получением чего-то.
А мы, слыша эту часть, как-то понимаем, что это не абракадабра, а что-то, связанное с бесплатным получением чего-то.
То, что не поменялось у нескольких однокоренных — это корень. Отмечаем.
Да, конечно, всегда будет 100500 исключений, чередований в корне и т.д. Но русскоговорящий человек (даже ребенок), как правило, эти чередования «считывает» на слух и понимает, что это чередования.
2. Находим окончание
Помним, что окончание так называется — потому что стоит в конце слова. Поэтому его очень просто искать: нужно обвести в квадратик сколько-то буковок в конце слова.
Осталось только выяснить, сколько буковок с конца — это окончание, а не что-то другое.
Напомню, мы уже определили корень.
Возможно, после определения корня ни одной лишней буковки в конце слова не оказалось.
У окончания есть такое явление, как нулевое окончание: это когда окончания нет — но оно как бы есть. Тогда мы ставим в конце слова пустой квадратик — обвели воздух в виде окончания.
Тогда мы ставим в конце слова пустой квадратик — обвели воздух в виде окончания.
Всё слово — один сплошной корень. Окончание нулевое
А еще бывает, что окончания нет в принципе — и обводить воздух в виде нулевого окончания не нужно.
До 4 класса ребенок почти наверняка с этим явлением не столкнется. Но если вдруг…
Как определить — есть ли у слова нулевое окончание или окончания нет вовсе?
Окончаний нет у слов, которые не умеют изменяться по падежам, лицам, числам, временам… чему угодно:
- Союзов, предлогов, междометий — разных коротких служебных словечек, которые сами по себе никакого смысла не несут, а лишь «помогают» связываться словам друг с другом (ах/ох, перед/над…).
- Существительных и прилагательных, которые нельзя изменить (пальто (цвета) индиго — и существительное и прилагательное здесь будут без окончания. .. ). Здесь же — аббревиатуры и сокращения вроде «минфин».
- У всех слов, которые могут ответить на вопрос КАК.


По последнему пункту интересно. «Официально» окончание не существует у деепричастий (что делая? — играя), а также прилагательных (какой? — красивее) и наречий (как? — ярче) в простой сравнительной форме.
А теперь попробуйте к тем же словам, которые я написала, задать тот же вопрос КАК:
- Играя, он вошел в комнату. — КАК он вошел в комнату?
- Он пишет красивее, чем Вася. — КАК он пишет?
- Сегодня солнце светит ярче. — КАК светит солнце?
Видите, несмотря на то, что речь идет про разные части речи, вопрос КАК все равно поможет определить, есть ли у слова окончание.
Хорошо, с нулевым или отсутствующим окончанием разобрались.
А, допустим, нам не повезло, и следом за корнем остались какие-то буковки. Наша задача — найти среди этих буковок окончание.
Для удобства запомним себе подсказки:
** Если разбираем существительное, прилагательное или причастие — будем говорить наше слово в компании со следующими тремя:
- НЕТ (чего?) нашего слова. Напр. «подарка«
- ДОВОЛЕН (чем?) нашим словом. Напр., «подарком«
- ДУМАЮ О (чем?) нашем слове. Напр., «подарке«
 Напр. «подарка«
Напр. «подарка«** Если разбираем глагол — скажем его в настоящем и прошедшем времени:
- что Я ДЕЛАЮ — ОНА ДЕЛАЕТ — МЫ ДЕЛАЕМ сейчас ? — Напр., «думаю — думает — думаем«
- ЧТО Я ДЕЛАЛ — ОНА ДЕЛАЛА — МЫ ДЕЛАЛИ вчера? — Напр. «думал — думала — думали«
То есть существительные и прилагательные мы изменяем по падежам, а глаголы — по лицам и числам в настоящем и прошедшем времени.
Их гораздо больше, чем я показала здесь.
Но нам для разбора по составу больше не нужно — возьмите только самые «показательные» в смысле изменений.
Проговариваем слова и наблюдаем, что изменилось в конце слова.
То, что изменилось после наших изменений — и будет окончанием. Отмечаем.
Здесь, к счастью, исключений будет минимум. Главное — быть очень внимательным. Иногда имеет смысл записать все получившиеся формы на черновичок в столбик — и глазами посмотреть, что изменяется.
3. Находим приставку
Приставка — это то, что стоит перед корнем.
Возможно, вам повезло, и перед корнем у вас ничего не стоит. УРА! Значит, приставки нет — двигаемся дальше.
Если же перед корнем что-то написано — это 100% приставка. А может — несколько приставок.
Четыре приставки в слове
Как определить: одна приставка, или две, или вообще несколько?
В начальной школе ребенок с 99% вероятностью ни разу не столкнется с двумя и более приставками — так что смело отмечайте всё, что оказалось перед корнем, одной квадратной загогулинкой.
Но если вдруг… тогда читайте дальше:
Приставки похожи на предлоги. Если то, что написано перед корнем, выглядит очень-очень длинно — попробуйте выделить оттуда кусочки, похожие на предлоги.
В нашем примере ребенок легко найдет слово «под» целых два раза. Скажем вместе с существительным: «под столом» — работает! Попробуем выделить его в отдельную приставку.
ПОД- в начале слова — это скорее всего приставка
Осталось всего два кусочка: «пере-» и «-вы-«. Проверим, могут ли они быть отдельными приставками.
Осталось проверить, могут ли считаться приставками другие кусочки
Для проверки приставок удобно использовать базовое слово «ХОД» — оно подходит к очень многим приставкам, к большинству из них. Добавляйте к этому слову все кусочки, которые вы считаете отдельными приставками. Если новое слово, которое получилось из слова «ход» и вашей предполагаемой приставки звучит адекватно — значит, вы правильно выделили отдельную приставку.
Добавим к слову «ход» эти приставки. Получилось «переход» и «выход» — вполне себе адекватные слова. Значит, обе приставки тоже имеют право на существование.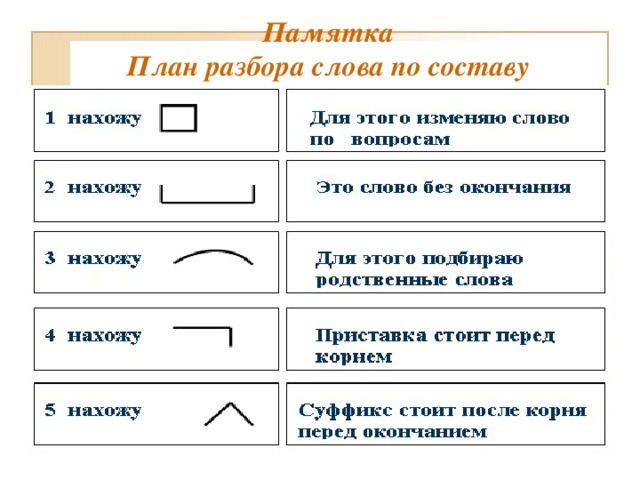
Конечно, слово «ход» подойдет далеко не ко всем приставкам.
Мы это обсудим в отдельной статье на тему приставок.
Там же поговорим о том, как отличить приставку, скажем, «по-» или «под-» от одноименного кусочка корняю
Итак, запоминаем:
- Всё, что перед корнем — это приставка.
- Если перед корнем ничего не написано — значит, приставки нет.
- Если оно слишком длинное — может, приставок несколько.
- Если нам нужно проверить, одна или несколько приставок — пробуем каждую «предположительно отдельную» приставку в качестве предлога или в качестве одной отдельной приставки к другому слову.
4. Находим суффикс
Суффикс — это то, что находится между корнем и окончанием.
Возможно, вам повезло, и между корнем и окончанием никаких букв не написано — УРА! Значит суффикса нет.
Если между корнем и окончанием вы видите только одну букву — смело отмечайте ее как суффикс.
Если же между корнем и окончанием оказалось больше одной буквы — тут начинаются нюансы: суффиксов может быть несколько.
Пример слов с двумя суффиксами
…Вообще, суффиксы — это самая мерзостная тема русского языка, потому что суффиксов немерено, и логика в их выделении присутствует далеко не всегда.
Поэтому в суффиксах нужно искать логику и заучивать их наизусть.
У меня выйдет цикл статей о суффиксах: как выстроить логическую систему суффиксов русского языка, запомнить их и не ошибаться при морфемном разборе.
Запомните: если в суффиксе несколько букв — а в начале или в конце «потенциального суффикса» стоит Л, К или Н — всегда подозревайте, что эта буква — это ОТДЕЛЬНЫЙ СУФФИКС (а всего в слове — два или больше суффиксов).
Дело в том, что буквы Л, К и Н чаще других букв выступают в роли отдельных суффиксов.
Здесь буква Н — в начале «потенциально суффикса», а значит, её можно подозревать в том, что она — отдельный суффикс
Что интересно, букв в суффиксе может быть совсем мало, и все равно буква Л, Н или К может оказаться отдельным суффиксом.
Буква К в конце «потенциального суффикса» с очень высокой вероятностью будет отдельным суффиксом — несмотря на то, что тут между корнем и окончанием всего 3 буквы
Как, например, в глаголах прошедшего времени — Л всегда будет отдельным суффиксом:
В общем, с буквами Н, К и Л в начале и конце потенциального суффикса будьте внимательны — с высокой степенью вероятности они будут выделяться в отдельный суффикс, а всего в слове будет больше одного слова, как в наших примерах:
Примеры с двумя суффиксами, с буквами Н, К и Л
Итак, запоминаем:
- То, что стоит между корнем и окончанием — это один или несколько суффиксов.
- Если между корнем и окончанием нет ни одной буквы — значит, суффикса нет.
- Если в начале или конце потенциального суффикса есть буква Н, Л или К — высока вероятность того, что эта буква — отдельный суффикс, а в слове больше, чем один суффикс.
- Чтобы безошибочно определять суффиксы — придется учить их наизусть, и вообще этот навык нарабатывается, увы, с опытом.

5. Определяем основу
И снова — самое простое. Всё, что не окончание — это основа.
Выделим квадратной закорючкой всё, кроме квадратика окончания — и разбор закончен!
***
Не забывайте, что морфемный анализ — это навык, который нарабатывается большим количеством повторений.
Поэтому, чтобы хорошо научиться разбирать слово по составу, нужно повторять это много — много — много — очень много раз, причем с разными частями речи.
Успехов у учебе вашим детям!
______________________
Мой блог про японскую методику обучения детей KUMON ищите по адресу https://kumon-deti. com
Познакомиться со мной в Инстаграм https://instagram.com/kumon.deti
Определение, фонетический (звуко-буквенный) разбор и разбор слова по составу
На данной странице представлено лексическое значение слова «хмуриться», а также сделан звуко-буквенный разбор и разбор слова по составу с транскрипцией и ударениями.
Оглавление:
- Значение слова
- Звуко-буквенный разбор
- Разбор по составу
Значение слова
ХМУРИТЬСЯ, рюсь, ришься; несов.
1. Хмурить лицо, брови. Недовольно, сердито х.
2. (1 и 2 л. не употр.), перен. Становиться пасмурным, сумрачным. Погода хмурится. Небо хмурится (собираются облака, тучи).
| сов. нахмуриться, рюсь, ришься.
Фонетический (звуко-буквенный) разбор
хму́риться
хмуриться — слово из 3 слогов: хму-ри-ться. Ударение падает на 1-й слог.
Транскрипция слова: [хмур’иц:а]
х — [х] — согласный, глухой непарный, твёрдый (парный)
м — [м] — согласный, звонкий непарный, сонорный (всегда звонкий), твёрдый (парный)
у — [у] — гласный, ударный
р — [р’] — согласный, звонкий непарный, сонорный (всегда звонкий), мягкий (парный)
и — [и] — гласный, безударный
т — [ц:] — согласный, глухой непарный, твёрдый (непарный, всегда произноится твёрдо)
ь — не обозначает звука
с — не образует звука в данном слове
я — [а] — гласный, безударный
В слове 9 букв и 7 звуков.
При разборе слова используются правила:
- Сочетание букв -тьс- обозначает длинный звук [ц:]
Цветовая схема: хмуриться
Ударение в слове проверено администраторами сайта и не может быть изменено.
Разбор слова «хмуриться» по составу
хмуриться (программа института)
хмуриться (школьная программа)
Части слова «хмуриться»: хмур/и/ть/ся
Часть речи: глагол
Состав слова:
хмур — корень,
и, ть, ся — суффиксы,
нет окончания,
хмури + ся — основа слова.
Примечание: ть является формообразующим суффиксом и не входит в основу слова, но во многих школьных программах ть отмечается как окончание. ся является словообразующим суффиксом (постфиксом) и входит в основу слова.
Гдз по русскому языку 8 класс Ладыженская, Тростенцова Решебник
Русский язык усердно и интенсивно изучают во всех школах Российской Федерации. Для большинства детей он является родным, но некоторые учащиеся (например, временно проживающие на нашей земле иностранцы) начинают изучать предмет чуть ли не с нуля. Данная дисциплина отнимает существенное время современного школьника и требует известных инвестиций внимания, мышления и прилежания.
Данная дисциплина отнимает существенное время современного школьника и требует известных инвестиций внимания, мышления и прилежания.
Т.А. Ладыженская, Л.А. Тростенцова, А.Д. Дейкина, О.М. Александрова создали новый учебно-методический комплекс по курсу родной речи для общеобразовательной школы. Он включает собственно учебник, рабочую тетрадь, а также решебник, по которому можно проверять правильность выполнения. Настоящие ГДЗ активно переиздаются издательством «Просвещение» в 2014-2019 годах. На нашем сайте вы всегда найдете самую актуальную версию.
ГДЗ по русскому Ладыженской, Тростенцовой любимый помощник восьмиклассников
Каждое упражнение подробно объяснено, присутствуют информативные схемы и интересные иллюстрации для облегчения понимания той или иной филологической концепции. Информация изложена кратко и точно, в соответствии с действующими нормативами для средней школы. Популярность пособия связана с качественными разъяснениями и вспомогательными комментариями. На страницах сборника можно найти практически всё, что нужно: разбор слова по составу, определение синтаксических элементов, культуру повествования, правила орфографии. Существуют следующие особенности, которые обязательно требуется отметить:
На страницах сборника можно найти практически всё, что нужно: разбор слова по составу, определение синтаксических элементов, культуру повествования, правила орфографии. Существуют следующие особенности, которые обязательно требуется отметить:
- легко искать нужные именно вам номера;
- приведены только новейшие версии пособий, утвержденные ведущими педагогами страны;
- развиваются реальные речевые навыки у школьников;
- сайт доступен во всех популярных устройств: телефонов, смартфонов и ноутбуков.
Решебник, созданный Ладыженским, прежде всего, показывает примеры правильного выполнения упражнений. Поэтому учебный результат достигается быстрее, не требует приложения школьником избыточных когнитивных усилий. Смело готовьтесь к контрольным, самостоятельным, проверочным работам, тестам. Вы скоро заметите более высокие оценки и откроете в себе мотивацию к более глубокому изучению предмета.
Что входит в онлайн-решебник (авторы: Ладыженская, Александрова) для 8 класса по русской речи?
Большая часть разделов посвящена синтаксическому разору и изучению новых правил пунктуации. Несколько параграфов помогают школьнику повторить пройденный материал. Такой расклад необходим, ведь ученики к концу года частично забывают то, что учили в самом его начале. Книга учит новым подходам в восприятии, позволяет глубже увидеть сущность предмета, развить интуитивную грамотность. Основные темы в текущем году:
Несколько параграфов помогают школьнику повторить пройденный материал. Такой расклад необходим, ведь ученики к концу года частично забывают то, что учили в самом его начале. Книга учит новым подходам в восприятии, позволяет глубже увидеть сущность предмета, развить интуитивную грамотность. Основные темы в текущем году:
- вводные и вставные конструкции;
- обращение и пунктуация при нем;
- обособленные члены предложения;
- полноценный синтаксический разбор.
Сборник, который вы можете видеть на данной странице предназначен для учеников. Он соответствует важнейшим требованиям ФГОС. Упражнения могут стать частью авторских программ передовых репетиторов.
Корейская комбинаторная категориальная грамматика и статистический анализ
Адес, А. и М. Стидман. «По порядку слов». Лингвистика и философия , 4 (1982), стр. 517–558.
Google Scholar
Блэк, Э., С. Абни, Д. Фликенгер, К. Гданец, Р. Гришман, П. Харрисон, Д. Хиндл, Р. Ингрия, Ф. Елинек, Дж. Клаванс, М. Либерман, М. Маркус, С. Рукос, Б. Санторини и Т. Стшалковски. Proc. Четвертого семинара DARPA по речи и естественному языку , Процедура количественного сравнения синтаксического покрытия грамматик английского языка, 1991.
Харрисон, Д. Хиндл, Р. Ингрия, Ф. Елинек, Дж. Клаванс, М. Либерман, М. Маркус, С. Рукос, Б. Санторини и Т. Стшалковски. Proc. Четвертого семинара DARPA по речи и естественному языку , Процедура количественного сравнения синтаксического покрытия грамматик английского языка, 1991.
Бозсахин, К. «Получение структуры предиката-аргумента для языка со свободным порядком слов». Proceedings of COLING-ACL ’98, 1998.
Cha, J., G. Lee and J.-H. Ли. «Обобщенное угадывание неизвестных морфем для гибридной POS-маркировки корейского языка». Проведение шестого семинара по очень большим корпусам в Coling-ACL 98 , Монреаль, Канада, 1998.
Charniak, E. Работа с контекстными грамматиками и статистикой слов , {технический отчет} CS-95- 28, Университет Брауна, 1995.
Чо, Х. и Дж. К. Парк. Труды 11-й корейской конференции по языку и обработке информации , Комбинированная категориальная грамматика и синтаксический анализ, 1999.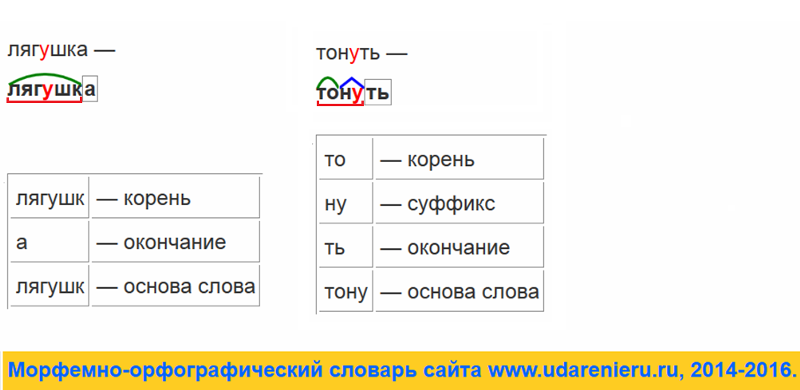
Collins, M. Proceedings of the 34th Annual Meeting of the ACL, Santa Cruz , A New Statistical Parser На основе лексических зависимостей биграмм, 1996.
Collins, M. Proceedings of the 35th Annual Meeting of the ACL , Three Generative, Lexicalized Models for Statistical Parsing, 1997.
Han, S.-K. и К.-Г. Парк. Труды 2-й конференции корейской и корейской обработки информации , Комбинированная категориальная грамматика корейского языка, 1990.
Хиндл, Д. и Х. Рут. «Структурная неоднозначность и лексические отношения». Компьютерная лингвистика , 19 (1) (1993), стр. 103–120.
Google Scholar
Hoffman, B. Proceedings of the 30th Annual Meeting of ACL, Student Session , A CCG Approach to Free Word Order Languages, 1992.
Hoffman, B. Протоколы 31-го ежегодного собрания ACL , Формальные последствия использования переменных в категориях CCG, 1993.
Hoffman, B. Proceedings of the European Chapter of the Association for Computational Лингвистика , Интеграция синтаксиса «свободного» порядка слов и информационной структуры, Дублин, 1995a.
Proceedings of the European Chapter of the Association for Computational Лингвистика , Интеграция синтаксиса «свободного» порядка слов и информационной структуры, Дублин, 1995a.
Hoffman, B. Вычислительный анализ синтаксиса и интерпретация «свободного» порядка слов в турецком языке , Ph.Докторская диссертация, Университет Пенсильвании, Отчет IRCS 95-17, 1995b.
Kim, C., J.H. Ким, Дж. Со и Г.К. Ким. «Анализ диаграммы справа налево с указанием путей для корейской грамматики зависимостей». Компьютерная обработка китайских и восточных языков , 8, приложение, 1994.
Lee, K.J. «Вероятностный анализ с использованием структурного предпочтения и совпадения головы и головы». Кандидат наук. диссертация, КАИСТ, корейский, 1997.
Google Scholar
Ли, К.Дж. И К.С. Ким. Joint Conference on Intelligence Technology , Tree Transformation Rules for Korean Lexicalized Multi-Component TAG Parser, Korean, 1995.
Lee, N. -S. Ящик и маркер ящика , Weol-In, 1998.
-S. Ящик и маркер ящика , Weol-In, 1998.
Lee, W., G. Lee and J. Lee. «Табличный нейросинтаксический анализ корейского языка». Proceedings of the Coling-94 , Киото, Япония, 1994, стр. 911–915.
Ли, У., Дж. Ли и Дж. Ли. «Управляемый диаграммами коннекционистский категориальный разбор разговорного корейского языка». Компьютерная обработка восточных языков , 10 (2) (1996), стр. 147–159.
Google Scholar
Ли, У., Дж. Ли и Дж. Ли. «Морфо-синтаксическое моделирование корейского языка с категориальной грамматикой». Труды симпозиума Тихоокеанского региона по обработке естественного языка , Пхукет, Таиланд, 1997 г., стр. 545–548.
Ли, Чарльз / Сандра Томпсон. Предмет и тема: новая типология языка , Academic Press, NY, 1996.
Google Scholar
Магерман, Д.М. и М. Маркус. «Анализ домена Вояджера с использованием Жемчужины». Proc. Семинара DARPA по речи и естественному языку , 1991, стр. 231–236.
231–236.
Магерман, Д.М. и К. Вейр. «Эффективность, надежность и точность при тщательном анализе диаграмм». В процессе. На 30-м ежегодном собрании доц. Для компьютерной лингвистики (ACL – 92) , 1992, стр. 40–47.
Сео, Дж. Korean Grammar , Hanyang University Press, Korean, 1996.
Google Scholar
Son, S. Исследование корейского вспомогательного глагола , Korean Culture Press, Сеул, корейский язык, 1996.
Google Scholar
Стидман М. «Зависимость и координация в грамматике голландского и английского языков». Язык , 61 (1985), стр. 523–568.
Google Scholar
Стидман, М.«Комбинаторные грамматики и паразитические пробелы». Естественный язык и лингвистическая теория , 5 (1987), стр. 403–439.
Google Scholar
Рабочий лист правил производных
Различайте каждый термин. Разделите функцию на ее члены и найдите производную каждого члена. Затем сложите производные. Таким же образом применяется и к вычитанию. Постоянные кратные — это частный случай правила сумм. ddx [f (x) + g (x)] = ddx [f (x)] + ddx [g (x)] \ frac {d} {dx} [f (x) + g (x)] = \ frac {d} {dx} [f (x)] + \ frac {d} {dx} [g (x)] dxd [f (x) + g (x)] = dxd [f (x)] + dxd [ g (x)] ddx [f (x) — g (x)] = ddx [f (x)]… Экспонент правила и практика. 1. ПРАВИЛО ПРОДУКТА: умножать, когда два основания одинаковы. 2. ЧАСТНОЕ ПРАВИЛО: чтобы разделить, когда два основания одинаковы, запишите основание и ВЫЧИТАЙТЕ …
Таким же образом применяется и к вычитанию. Постоянные кратные — это частный случай правила сумм. ddx [f (x) + g (x)] = ddx [f (x)] + ddx [g (x)] \ frac {d} {dx} [f (x) + g (x)] = \ frac {d} {dx} [f (x)] + \ frac {d} {dx} [g (x)] dxd [f (x) + g (x)] = dxd [f (x)] + dxd [ g (x)] ddx [f (x) — g (x)] = ddx [f (x)]… Экспонент правила и практика. 1. ПРАВИЛО ПРОДУКТА: умножать, когда два основания одинаковы. 2. ЧАСТНОЕ ПРАВИЛО: чтобы разделить, когда два основания одинаковы, запишите основание и ВЫЧИТАЙТЕ …
Производная экспоненциальной функции может быть получена с использованием определения производной. Производные экспоненциальных функций включают функцию натурального логарифма, которая сама по себе является важным пределом в исчислении, а также исходную экспоненциальную функцию. Производная — это натуральный логарифм основания, умноженного на исходную функцию.30 апреля 2018 г. · «Производная произведения двух функций — это первое, умноженное на производное второго, плюс второе, умноженное на производное первого».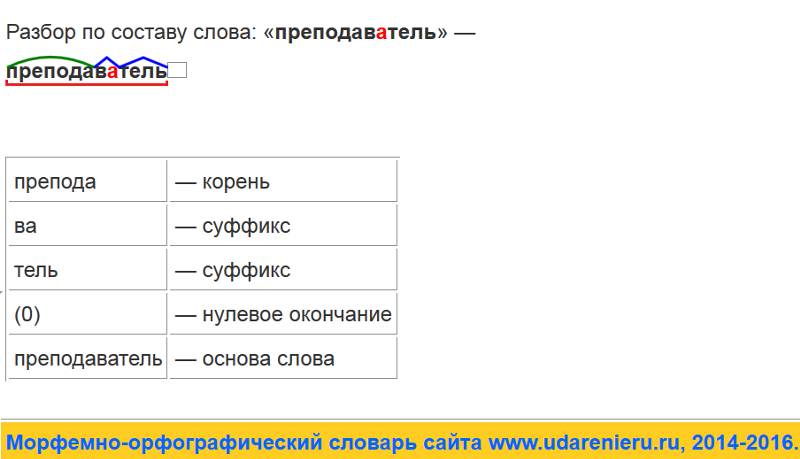 Откуда взялась эта формула? Как и все встречающиеся нам формулы дифференцирования, она основана на производных от первых принципов. Пример 1. Если у нас товар понравился. y = (2x 2 + 6x) (2x 3 + 5x 2)
Откуда взялась эта формула? Как и все встречающиеся нам формулы дифференцирования, она основана на производных от первых принципов. Пример 1. Если у нас товар понравился. y = (2x 2 + 6x) (2x 3 + 5x 2)
Рабочий лист: Правила дифференцирования Вычисляйте производные, используя правила дифференцирования, особенно правила произведения, частного и цепочки.Упростите свои ответы, чтобы мы могли сравнить результаты. 1. f (x) = 2×2 k c + x3, где c и k постоянные 2. y = 2 (2 sinx) cosx 3. f (x) = 7 + x3 5 4. y = ln 1 + ex 1 xe Решение : Используя правила производной, f ‘(x) = 6x — 4. При x = 1 производная равна 6 (1) — 4 = 2. Может. также можно записать как f ‘(1) = 2. Домашние упражнения. Часть 1: Производные полиномиальных функций.
· Рабочие листы цепных правил с ответами 6 октября 2019 г. 1 октября 2019 г. Некоторые из приведенных ниже рабочих листов представляют собой рабочие листы цепных правил с ответами, использование цепного правила для получения производных функций, несколько интересных упражнений на цепные правила с пошаговыми решениями и викторины с ответами,… Определение производных рабочих листов Это исчисление — рабочий лист правил дифференциации будет производными правилами — постоянным правилом, постоянным множественным правилом, правилом мощности, правилом суммы, правилом разности. ..
..
Рабочий лист правил делимости — Концепция — Проблемы с пошаговыми решениями. Рабочий лист правил делимости. Проблема 1. Проверьте, делится ли 16 на 2,1. Глава 2 Производные правила 2. Следующее слайд-шоу содержит основные производные правила из Главы 2. В ходе курса будет больше. Сначала вам дается выражение
Мэриленд, бюджет штата
ТСЖ, по закону, поддерживает улицы, благоустройство и благоустройство. Наиболее значительные ежегодные расходы идут на поддержание парковой обстановки.Поскольку правление входит в бюджетный сезон, мы подумали, что для домовладельцев будет полезно иметь больше…
Olney, MD 20832 (301) 924-4050 (301) 924-9389 (факс) Управляющий агент: Лаура Этчисон (доб. 324) [адрес электронной почты защищен] Помощник менеджера: Эмбер Бекманн (вн. тел. 317) [адрес электронной почты защищен] Заседания Совета ТСЖ проводятся ежемесячно в первый понедельник месяца, 19:00 в начальной школе Гиббса в Медиацентре.
Summit Ridge Homeowners Association% Управление общественными ассоциациями 3655A Old Court Road, Suite 15 Pikesvile, MD 21208 После получения запроса он рассматривается членами комитета по архитектурной проверке. Этот комитет состоит из трех директоров и четырех других домовладельцев, которые потратили свое время.
Этот комитет состоит из трех директоров и четырех других домовладельцев, которые потратили свое время.
21 ноября 2020 г. · Community Association Services, Inc., AAMC 18401 Woodfield Road, Suite H Gaithersburg, MD 20879 Офис: 301-840-1800 Бесплатный звонок: 800-883-0799 Экстренная помощь в нерабочее время: 888-788-2678
Совет директоров Ассоциации домовладельцев Потомакского хребта утвердил операционный бюджет на финансовый год, начинающийся 1 января 2020 года. С утвержденным кратким изложением бюджета и описательными примечаниями можно ознакомиться, перейдя по ссылке ниже.
The View at Crystal Falls — официальный информационный бюллетень ТСЖ Crystal Falls The View at Crystal Falls объединяет актуальные общественные новости и информацию, освещает дома и предприятия района и освещает местные события. Основанный в 2006 году, The View at Crystal Falls является «официальным» информационным бюллетенем сообщества HOA Crystal Falls. Он опубликован CFHOA. Продолжить чтение
ATLANTA (PRWEB) 3 октября 2018 г.
Разбор предложения и слов по составу
В русском языке, в практической его части, существует большое количество самых разных разборов: от разбора слова, словосочетания до разбора предложения. Ни один диктант или контрольная работа не обходится без них. Объяснить это достаточно просто: при выполнении того или иного разбора проверяются базовые знания по предмету, будь то фонетика, морфология, синтаксис или пунктуация.
Как же быть ученику, ведь количество этих разборов достаточно большое и все они такие разные? А сколько знаний нужно, чтобы грамотно их делать!
По этому поводу можно сказать только одно: не нужно драматизировать. Все разборы в русском языке можно систематизировать, разобраться в их алгоритме (последовательности), и тогда на протяжении всего обучения в школе каждый ученик будет успешен.
Давайте рассмотрим порядок некоторых из них.
Фонетический разбор слова
- Выписать слово.
- Определить, сколько в слове слогов.
- Определить, на какой слог падает ударение.
- Дать характеристику каждого звука:

- гласные — ударные,
- безударные;
- согласные — звонкие или глухие, твёрдые или мягкие.
- Указать, какой буквой обозначены звуки.
- Определить, сколько в слове букв и звуков.
Образец:
Дальний — 2 слога (1-ый ударный)
д-[д]- согл., зв., тв.
а-[а]- гл., уд.
л-[л`]- согл., зв., мягк.
ь-[-]
н-[н`]- согл., зв., мягк.
и-[и]- гл., безуд.
й-[й`]- согл, зв., мягк.
7 букв, 6 звуков
Разбор слова по составу
Подарки
- Выписать слово.
- Найти окончание (для этого нужно изменить форму слова, изменяемая его часть и будет окончанием: подарки — подарок, подарка, подарку; в данном случае — окончание — и).
- Выделить основу слова (это часть слова без окончания, в данном случае подарки).
- Найти корень (для этого нужно подобрать несколько однокоренных слов и определить их общую часть: подарки-дарить, дарёный, одаривать, дар, в данном случае корень — дар-).
- Найти приставку (для этого нужно сравнить данное слово с однокоренными словами с разными приставками или без них: подарки-одарить, надарить, дарить, приставка в данном случае по-).
- Найти суффикс (для этого нужно подобрать несколько однокоренных слов с разными суффиксами или без него: подарки — подарочки, подарить, в данном случае суффикс -к-).

Состав слова подарки: приставка по-, корень -дар-, суффикс -к-, окончание -и.
Разбор простого предложения по составу грамматической основы
По наличию главных членов простые предложения делятся на двусоставные и односоставные.
Грамматическая основа двусоставных предложений состоит из двух главных членов предложения — подлежащего и сказуемого, например: Белая берёза под моим
окном принакрылась снегом, точно серебром. (С. Есенин).
В односоставных предложениях грамматическая основа состоит из одного главного члена — подлежащего или сказуемого. В зависимости от этого они бывают назывными или глагольными.
В зависимости от этого они бывают назывными или глагольными.
Назывные — это односоставные предложения с одним главным членом — подлежащим. Например: Зима! Крестьянин, торжествуя, на дровнях обновляет путь (А.Пушкин).
В глагольных односоставных предложениях главный член — сказуемое. В зависимости от формы сказуемого все глагольные односоставные предложения делятся на определённо-личные (сказуемое — глагол в форме 1-го и 2-го лица), неопределённо-личные (сказуемое -глагол в форме 3-го лица мн. ч.), обобщённо-личные (сказуемое — глагол в форме 2-го лица, чаще всего это пословицы и поговорки), безличные (предложение со сказуемым, при котором нет и не может быть подлежащего).
Образец разбора: гл. 3 л.
За рекой косили. (Предложение повествовательное, невосклицательное, односоставное, неопределённо-личное, распространённое).
Разбор предложения по составу (второстепенные члены предложения)
По наличию или отсутствию второстепенных членов предложения простые предложения — двусоставные и односоставные делятся на нераспространённые и распространённые.
Нераспространённые простые предложения в своём составе имеют только главные члены предложения. Например: Катятся ядра, свищут пули …(А.Пушкин).
Распространённые простые предложения имеют второстепенные члены предложения. Например: Весело сияет месяц над селом (И.Никитин).
При характеристике предложения обязательно указывается: распространённое предложение или нераспространённое (см. пример выше).
Разбор предложения по наличию или отсутствию необходимых членов предложения
Иногда в предложении могут отсутствовать какие-либо члены предложения, но смысл предложения понятен из контекста. Такие предложения называются неполными.
Пример полных предложений: С запада надвигалась дождевая туча. Постепенно темнело.
Пример неполного предложения: Товарищ шёл в библиотеку, а я — в бассейн. В данном случае во втором простом предложении пропущен глагол «шёл», но смысл предложения абсолютно понятен из предыдущего. При характеристике неполного предложения на этот факт обязательно указывается.
При характеристике неполного предложения на этот факт обязательно указывается.
Слова «семья» морфологический и фонетический разбор
Объяснение правил деление (разбивки) слова «семья» на слоги для переноса.
Онлайн словарь Soosle.ru поможет: фонетический и морфологический разобрать слово «семья» по составу, правильно делить на слоги по провилам русского языка, выделить части слова, поставить ударение, укажет значение, синонимы, антонимы и сочетаемость к слову «семья».
Содержимое:
- 1 Слоги в слове «семья» деление на слоги
- 2 Как перенести слово «семья»
- 3 Морфологический разбор слова «семья»
- 4 Разбор слова «семья» по составу
- 5 Сходные по морфемному строению слова «семья»
- 6 Синонимы слова «семья»
- 7 Ударение в слове «семья»
- 8 Фонетическая транскрипция слова «семья»
- 9 Фонетический разбор слова «семья» на буквы и звуки (Звуко-буквенный)
- 10 Предложения со словом «семья»
- 11 Сочетаемость слова «семья»
- 12 Значение слова «семья»
- 13 Склонение слова «семья» по подежам
- 14 Как правильно пишется слово «семья»
- 15 Ассоциации к слову «семья»
Слоги в слове «семья» деление на слоги
Количество слогов: 2
По слогам: семь-я
ь всегда примыкает к предшествующей согласной, смягчая её
Как перенести слово «семья»
се—мья
Морфологический разбор слова «семья»
Часть речи:
Имя существительное
Грамматика:
часть речи: имя существительное;
одушевлённость: неодушевлённое;
род: женский;
число: единственное;
падеж: именительный;
отвечает на вопрос: (есть) Что?
Начальная форма:
семья
Разбор слова «семья» по составу
| семь | корень |
| я | окончание |
семья
Сходные по морфемному строению слова «семья»
Сходные по морфемному строению слова
Синонимы слова «семья»
1. родные
родные
2. фамилия
3. дом
4. род
5. династия
6. юрт
7. рой
8. задруга
9. близкие
10. хомут
Ударение в слове «семья»
семья́ — ударение падает на 2-й слог
Фонетическая транскрипция слова «семья»
[с’им’й’а]
Фонетический разбор слова «семья» на буквы и звуки (Звуко-буквенный)
| Буква | Звук | Характеристики звука | Цвет |
|---|---|---|---|
| с | [с’] | согласный, глухой парный, мягкий, шумный | с |
| е | [и] | гласный, безударный | е |
| м | [м’] | согласный, звонкий непарный (сонорный), мягкий | м |
| ь | — | не обозначает звука | ь |
| я | [й’] | согласный, звонкий непарный (сонорный), мягкий | я |
| [а] | гласный, ударный |
Число букв и звуков:
На основе сделанного разбора делаем вывод, что в слове 5 букв и 5 звуков.
Буквы: 2 гласных буквы, 2 согласных букв, 1 буква не означает звука.
Звуки: 2 гласных звука, 3 согласных звука.
Предложения со словом «семья»
В другом варианте реальна серьёзная ссора, в которую невольно втянутся многие члены семьи.
Источник: Татьяна Борщ, Лунный календарь для женщин на 2016 год + календарь стрижек, 2015.
Ибо даже британская королевская семья была немецкого происхождения — она сменила свою немецкую фамилию на английскую только в 1917 году!
Источник: Бернард Хаттон, Секретная миссия Рудольфа Гесса. Закулисные игры мировых держав. 1941-1945.
Сохранились свидетельства разных лиц, в той или иной степени причастных к тайне царской семьи.
Источник: В. Г. Сироткин, Анастасия, или Кому выгоден миф о гибели Романовых, 2010.
Сочетаемость слова «семья»
1. царская семья
2. королевская семья
3. большая семья
4. семьи детей
5. семья друга
6. в семье крестьянина
7. члены семьи
8. глава семьи
9. в кругу семьи
10. семья распалась
11. семья погибла
12. семья жила
13. создать семью
14. родиться в семье
15. происходить из семьи
16. (полная таблица сочетаемости)
Значение слова «семья»
СЕМЬЯ́ , -и́, мн. се́мьи, семе́й, се́мьям, ж. 1. Группа людей, состоящая из мужа, жены, детей и других близких родственников, живущих вместе. (Малый академический словарь, МАС)
Склонение слова «семья» по подежам
| Падеж | Вопрос | Единственное числоЕд.ч. | Множественное числоМн.ч. |
|---|---|---|---|
| ИменительныйИм. | что? | семья | семьи |
РодительныйРод. | чего? | семьи | семей |
| ДательныйДат. | чему? | семье | семьям |
| ВинительныйВин. | что? | семью | семьи |
| ТворительныйТв. | чем? | семьёй, семьёю | семьями |
| ПредложныйПред. | о чём? | семье | семьях |
Как правильно пишется слово «семья»
Орфография слова «семья»Правильно слово пишется: семья́
Гласные: е, я;
Согласные: с, м;
Нумерация букв в слове
Номера букв в слове «семья» в прямом и обратном порядке:
- 5
с
1 - 4
е
2 - 3
м
3 - 2
ь
4 - 1
я
5
Ассоциации к слову «семья»
Домохозяйство
Бедность
Выходец
Член
Домашние
Перепись
Достаток
Иммигрант
Отпрыск
Хозяйство
Благосостояние
Эмигрант
Соболезнование
Губерния
Переселенец
Домохозяйка
Благополучие
Пары
Переселение
Поколение
Пожилое
Первенец
Родство
Доход
Жительство
Ребёнок
Домочадец
Уклад
Мещанин
Воспитание
Землевладелец
Льгота
Наследование
Уезд
Представительница
Крестьянин
Зажиточный
Крестьянский
Потомственный
Языковой
Небогатый
Состоятельный
Аристократический
Супружеский
Дворянский
Дружный
Купеческий
Годовой
Знатный
Военнослужащий
Престарелый
Благополучный
Интеллигентный
Еврейский
Царский
Королевский
Домашний
Фамильный
Полноправный
Влиятельный
Фермерский
Добропорядочный
Алтайский
Тамбовский
Жилищный
Буржуазный
Родственный
Бездетный
Правящий
Служащий
Наследственный
Младший
Родовой
Богатый
Средний
Одинокий
Протестантский
Жилой
Полтавский
Императорский
Родиться
Воспитываться
Переехать
Переселить
Проживать
Эмигрировать
Хорошеть
Насчитываться
Переезжать
Воспитывать
Переселиться
Разориться
Прокормить
Обзавестись
Передаваться
Распасться
Голодать
Родом
Ныне
Совместно
Диктанты на тему «Зима» для 3 класса
-
Опубликовано
26.12.2020
- by Светлана
- в Русский язык, Третий класс

Диктанты на тему «Зима» для 3 класса. Очень много! И все — отличные! Я собрала их из открытых источников, перепроверила, подобрала к каждому грамматическое задание и рассортировала по количеству слов.
Диктанты удобно использовать для контроля знаний дома, а также для подготовки к диктантам, которые будут даны в классе. Если даже просто орфографически прочитать несколько диктантов и разобрать слова, вызвавшие затруднения, оценка за полугодовой диктант может существенно повыситься.
Диктанты на тему «Зима» для 3 класса
Диктант № 1 «Зимой» (41 слово)
Был сильный мороз. Он сковал ледком лесные озерки и речки. На дорожках скользко. Зябко стало дубкам и березкам. Подул резкий ветер. Качнулась гибкая ветка рябины. Испугался робкий зайчишка, прижал от страха ушки. Вдалеке каркнул ворон. Запищали синички.
Грамматическое задание
- Обозначьте над словами второго предложения к каким частям речи они относятся.
- Выпишите из текста два слова с проверяемыми согласными в корне. Рядом напишите проверочные слова.
- Разделите для переноса слова последнего предложения.
Диктант № 2 «Зима» (44 слова)
Прошла ненастная осень. Солнце греет все меньше. Наступила зима. Ударил сильный мороз. В воздухе чувствуется свежесть. Снег покрыл окрестности деревни. Счастливые мальчишки и девчонки бегут на каток. Лопатами чистят они снег, метлой подметают лед. Каток готов. Хорошо будет кататься по скользкому льду на коньках!
Грамматическое задание
- Подчеркните главные члены в восьмом предложении.
- Выпишите из текста два слова с непроизносимыми согласными звуками.
- Проведите фонетический разбор слова «меньше».
Диктант № 3 «Праздничный лес» (44 слова)
Ночная вьюга украсила лес. Он стал как чудесная сказка. Молоденькие кустики укрылись снежными шубками. Еловые лапы надели мягкие рукавицы. Гигантские сугробы напоминали великанов. Вот взошло солнце. Яркими блестками засветились снежинки. Из-под снега вылетел тетерев. Он зорко осмотрел местность и взлетел на гибкую ветвь березы.
Он стал как чудесная сказка. Молоденькие кустики укрылись снежными шубками. Еловые лапы надели мягкие рукавицы. Гигантские сугробы напоминали великанов. Вот взошло солнце. Яркими блестками засветились снежинки. Из-под снега вылетел тетерев. Он зорко осмотрел местность и взлетел на гибкую ветвь березы.
Грамматическое задание
- Сделайте синтаксический разбор первого предложения.
- Найдите в тексте слова с приставками, обозначьте в них приставки.
- Сделайте фонетический разбор слова «гигантские».
Диктант № 4 «Каток» (45 слов)
Мы живем в селе Морозки. В субботу все ребята вышли делать каток. Падал мягкий снежок. Чудесная стояла погодка. Ребята быстро расчистили площадку. Они залили ее водой. За ночь лед окреп. Утром дети уже катались на гладком льду. Легко бежали коньки по скользкой дорожке. Хорош каток!
Грамматическое задание
- Подчеркните основу первого предложения.
- Обозначьте окончания у слов второго предложения.
- Разберите по составу слово «расчистили».

Диктант № 5 «В море» (46 слов)
Зимой море замерзло. Рыбаки собрались на лед ловить рыбу. Рыбак Андрей взял с собой сынишку Володю. Далеко в море выехали рыбаки. Сделали во льду дырки, спустили в воду сети. Светило солнце. Всем было весело. Володя был рад. Он помогал выпутывать из сетей рыбу. Рыбы ловилось много.
Грамматическое задание
- Обозначьте окончания в словах первого предложения.
- Сделайте фонетический разбор слова «солнце».
Диктант № 6 «Снеговик» (47 слов)
Стоит чудесный зимний день. Падает лёгкий снег. Деревья одеты в белые шубки. Спит пруд под ледяной коркой. Яркое солнце на небе. Выбежала группа ребят. Они стали лепить снеговика. Глазки сделали ему из светлых льдинок, рот и нос из морковки, а брови из угольков. Радостно и весело всем!
Радостно и весело всем!
Грамматическое задание
- В первом предложении подчеркните главные члены и укажите части речи.
- Разберите по составу слово «зимний».
- Найдите в тексте и запишите по одному слову с проверяемым безударным гласным в корне, парным по глухости-звонкости согласным, непроизносимым согласным. Рядом запишите проверочные слова.
Диктант № 7«Зимний лес» (48 слов)
Чудесен русский лес зимой! Белый пушистый снег повис на ветвях деревьев. Смолистые шишки украшают вершины елей. Шустрые синицы пищат в сучьях. На сугробах видны узоры заячьих и лисьих следов. Вот бежит через дорогу белка. Прыгнула на сосну, махнула хвостиком. Полетела легкая снежная пыль. Постучал молоточком по стволу дятел.
Грамматическое задание
- Выполните синтаксический разбор второго предложения.
- Выпишите словосочетания из третьего предложения.
- Разберите по составу слово «морозец».

Диктант № 8 «Пернатые друзья» (48 слов)
Сережа с папой делали кормушку для птиц. Сережа подавал гвозди и дощечки. Папа их строгал и сколачивал. Зимой каждое утро Сережа сыпал в кормушку зерно. Воробьи и синички ждали мальчика. Они слетались со всех сторон, спешили позавтракать. Сережа кормил своих пернатых друзей часто. Сытая птица легко переносит холод.
Грамматическое задание
- Сделайте синтаксический разбор последнего предложения.
- Подчеркните разделительный мягкий знак.
- Разберите по составу слово «позавтракать».
Диктант № 9 «Что за зверь?» (48 слов)
Ночью был легкий мороз. Утром выпал мягкий пушистый снег. Деревья, крыши, ступеньки крыльца стали белые. Девочка Катя захотела по снежку погулять. Она вышла на крыльцо. На снегу были маленькие ямки. Какой это зверь ходил по снегу? У зверька теплая шубка и длинные уши. Он любит морковку. Это заяц.
Это заяц.
Грамматическое задание
- Найдите и обведите в тексте слово с разделительным мягким знаком.
- Разберите по составу слово «маленькие».
- Поставьте ударение над словами второго предложения.
Диктант № 10 «Следы на снегу» (48 слов)
Зимний лес. На снегу строчки и точки. Чьи это следы? Вот след лисицы. Не удалось лисе ночью поймать добычу. Здесь проскакал беляк. Зайчишка любит поглодать горькую осиновую кору. Пробежала куница в чащу. Хищница искала белок. Из-под снега вылетел тетерев. Он осмотрел местность и взлетел на гибкую ветвь берёзы.
Грамматическое задание
- Обозначьте основу в последнем предложении.
- Разберите по составу слово «поглодать».
- Обведите в тексте союзы.
Диктант № 11 «В зимнем лесу» (50 слов)
Ночью была метель. Утром лес было не узнать. Ночная вьюга украсила лесные поляны. Маленькие кустики укрылись снежными шубками. Мохнатые лапы колючих елей надели мягкие рукавицы. Гигантские сугробы стояли как сказочные великаны. Взошло солнце. Ясная берёзка заблестела тонкой берестой. Заискрился серебристый иней на гибких веточках. Яркими блёстками засветились снежинки на снегу.
Маленькие кустики укрылись снежными шубками. Мохнатые лапы колючих елей надели мягкие рукавицы. Гигантские сугробы стояли как сказочные великаны. Взошло солнце. Ясная берёзка заблестела тонкой берестой. Заискрился серебристый иней на гибких веточках. Яркими блёстками засветились снежинки на снегу.
Грамматическое задание
- Сделайте синтаксический разбор третьего предложения.
- В четвёртом предложении подчеркните прилагательные.
- Разделите для переноса слова последнего предложения.
Диктант № 12 «Чудесная ёлка» (51 слово)
Скоро Новый год. Ребята решили устроить в лесу праздник. Девочки стали украшать пушистую ёлку ягодками рябины. Коля привязал к ветвям кусочки сала. Петя прикрепил сладкую морковку. Таня посыпала на дощечку зерно. Под ёлку воткнули кочан капусты. Утром прилетели стайки птиц. Они радостно щебетали и угощались подарками. Сладкие капустные листья грыз заяц.
Грамматическое задание
- Во втором предложении подчеркните имена существительные.
- Обведите в тексте все имена собственные.
- Разделите для переноса слова последнего предложения.

Диктант № 13 «Чудесная ёлка» (52 слова)
Идёт мелкий снежок. Пушистые хлопья снега легли на землю и деревья. На поляне стояла молоденькая стройная ёлочка. Ребята решили украсить её. Они повесили на лесную красавицу ягоды рябины. К нижним веткам прикрепили морковку. Под ёлку положили кочан капусты. Утром над ёлкой кружилась стайка птиц. Вечером прибежали два зайца. Они угощались вкусным ужином.
Грамматическое задание
- Выпишите из текста 3 слова с безударной гласной в корне, которая проверяется ударением, укажите проверочное слово.
- Разберите по составу слово «кормушка».
- К данным словам подберите и запишите антонимы: продавать -…, выходить -….
Диктант № 14 «Зимний день» (52 слова)
Стоит чудесный зимний денёк. Под моё окошко летят красивые птички. Смотрю на птиц. Вот они сидят на ветвях кудрявой берёзы. На голове чёрная шапочка. Спина, крылья и хвостик жёлтые. На короткую шейку птичка, словно галстучек повязала. Будто жилет синица надела. Хороша птица! Клювик у птички тоненький. Едят синички вкусное сало. Радостно им.
Смотрю на птиц. Вот они сидят на ветвях кудрявой берёзы. На голове чёрная шапочка. Спина, крылья и хвостик жёлтые. На короткую шейку птичка, словно галстучек повязала. Будто жилет синица надела. Хороша птица! Клювик у птички тоненький. Едят синички вкусное сало. Радостно им.
Грамматическое задание
- Сделайте синтаксический разбор первого предложения.
- Разберите по составу слово «тоненький».
- Поставьте знаки ударения над словами второго предложения.
Диктант 15 «В гостях» (52 слова)
Был поздний вечер. Вокруг лесная глушь. Мороз крепчал. На небе блеснули звёздочки. Месяц осветил окрестность. По тропе пробежал заяц. Мы подъехали к дому лесника. Лесник Игорь Ильич пригласил нас в сторожку. Радостно завилял хвостом пёс Шарик. Лесник затопил печь. Дочь лесника пригласила нас к столу. Мы стали пить душистый чай с медом.
Грамматическое задание
- Обведите в тексте все имена собственные.
- Подчеркните в тексте все местоимения.
- Разделите на слоги слова последнего предложения.

Диктант № 16«На рыбалке» (52 слова)
Мы стали жить на берегу Волги. Зимой река замёрзла. Рыбаки собрались на лёд ловить рыбу. Рыбак Андрей взял с собой сынишку Ваню. Далеко выехали рыбаки. Сделали во льду ямки и спустили в воду сети. Светило солнце. Всем было весело. Ваня был очень рад. Он помогал выпутывать из сетей рыбу. Рыбы наловилось много.
Грамматическое задание
- Укажите части речи в первом предложении.
- Подчеркните в тексте собственные имена существительные.
- Выпишите из текста по два одушевлённых и неодушевлённых существительных.
Диктант № 17 «Зимой» (53 слова)
Был зимний денек. Синичка Зинька прыгала по веткам. Глазок у синички острый. Под корой деревьев она добывала насекомых. Вот Зинька раздолбила дырочку, вытащила букашку и съела. Тут выпрыгнула из снега мышка. Мышка дрожит, вся взъерошилась. Она объяснила Зиньке свой страх. Мышка провалилась в медвежью берлогу. Там крепко спали большая медведица и маленькие медвежата.
Тут выпрыгнула из снега мышка. Мышка дрожит, вся взъерошилась. Она объяснила Зиньке свой страх. Мышка провалилась в медвежью берлогу. Там крепко спали большая медведица и маленькие медвежата.
Грамматическое задание
- Обозначьте основу второго предложения.
- Выпишите из текста однокоренные слова к слову «медведь», обозначьте в них корень.
- Обозначьте окончания в словах третьего предложения.
Диктант № 18 «Весёлая ёлка» (53 слова)
Скоро Новый год. Таня и мама украшали пушистую елку. На праздник пришли гости. Все веселились. Только мама была грустная. Она ждала сына-моряка. Вдруг раздался звонок. Ребята быстро подбежали к дверям. В комнату вошел Дед Мороз. Он стал раздавать детям подарки. Потом Дед Мороз снял седую бороду. Мама увидела сына-моряка. Радостной была эта встреча!
Грамматическое задание
- Проведите синтаксический разбор второго предложения.
- Обведите в тексте все предлоги.
- Разделите для переноса слова последнего предложения.

Диктант № 19 «Зимние чудеса» (54 слова)
Зима вошла в свои права. Мягкий снег укрыл землю. Мороз сковал реки и озера. Крепкий гладкий лед блестит на пруду. Красив стал зимний лес. Пушистые варежки зимушка надела на ели и сосны. Снежком укрылись молодые березки и осинки. Старые пни надели на головы шапки. На полях волнистыми коврами лег снег. Перестал зябнуть озимый хлеб.
Грамматическое задание
- Сделайте синтаксический разбор второго предложения.
- Подчеркните буквы, обозначающие мягкие согласные звуки в третьем предложении.
- Обведите все имена существительные в предпоследнем предложении.
Диктант № 20 «Снежные фигурки» (56 слов)
С неба летел мокрый снежок. Ребята выбежали во двор и стали лепить из снега фигурки. Коля слепил снеговика. Хорош снеговик! В носу краснела морковка. В руке метла, а на голове ведро. Женя строил башню с ледяными окошками. Толя и Илья лепили Деда Мороза и Снегурочку. Дед Мороз был с бородой. Снегурочка в руках держала зеленую ёлочку.
Хорош снеговик! В носу краснела морковка. В руке метла, а на голове ведро. Женя строил башню с ледяными окошками. Толя и Илья лепили Деда Мороза и Снегурочку. Дед Мороз был с бородой. Снегурочка в руках держала зеленую ёлочку.
Грамматическое задание
- Обведите в тексте имена собственные.
- Сделайте синтаксический разбор последнего предложения.
- Разберите по составу слово «ёлочка».
Диктант № 21 «Как белочка зимует» (56 слов)
Закрутила метель. Белка поспешила к гнезду. Гнездо у зверька теплое. Забралась белка в гнездо и прикрылась пушистым хвостиком. Холодный ветер и мороз не страшны зверьку. Утихла непогода. Вылезла белка из гнезда. Она ловко поскакала с дерева на дерево. Вот белка взяла в лапки еловую шишку. Она стала ее обгрызать. Вкусная еда у зверька запасена в кладовой.
Грамматическое задание
- Обведите в тексте все союзы.
- Сделайте синтаксический разбор четвёртого предложения.
- Разделите для переноса слова последнего предложения.
Диктант № 22 «Зима» (57 слов)
Трещали морозы. Иней запушил деревья. На ели и сосне — шапки снега. Мороз затянул пруд льдом. Мягкий белый ковёр покрыл холодную землю. Школьники полили водой горку, размели каток. На катке шум и веселье. Вот покатил на коньках Юра. К Юре подъехал Вася. По дороге к деревне проезжал на лошади дядя Егор. Он с улыбкой смотрел на ребят.
Грамматическое задание
- Подчеркните основу второго предложения.
- Обведите все имена собственные.
- Подчеркните все существительные в четвёртом предложении.
Диктант № 23 Зимние забавы (57 слов)
На дворе зима. Ребята с нетерпением ждут первого снега. Так хочется прокатиться на санях или поиграть в снежки. Зима – самое веселое время года. Дети строят снежные горки. Как здорово скатиться с горки на санках! Ветер свистит в ушах, а мороз обжигает щеки. Весело! В лесу уже проложили лыжню. В воскресенье мы всем классом отправимся кататься на лыжах.
Весело! В лесу уже проложили лыжню. В воскресенье мы всем классом отправимся кататься на лыжах.
Грамматическое задание
- Обозначьте подлежащее и сказуемое во втором предложении.
- Придумайте и запишите однокоренные слова к слову «мороз».
- В третьем предложении подчеркните буквы, обозначающие твёрдые согласные звуки.
Диктант № 24 «Во льдах» (58 слов)
Ледокол «Серов» шел во льдах. Кругом ледяное море, ослепительный снег и голубые на снегу тени. Вдруг мы увидели семейство медведей. Они шли по льду друг за дружкой гуськом. Впереди медведица, позади два медвежонка. Медведица часто останавливалась, покачивала головой, нюхала воздух. Медвежата покорно следовали за ней. Звери шли неторопливо рядом с кораблем. Видимо, они приняли ледокол за большой айсберг.
Грамматическое задание
- Определите падежи всех имён существительных шестого предложения.
- Сделайте синтаксический разбор предпоследнего предложения.
- Обозначьте окончания в словах второго предложения.

Диктант № 25 «Снегири» (58 слов)
Это случилось в один погожий зимний день. Я возвращался из школы. Возле самого дома я увидел необычных птичек. Они были похожи на маленькие пушистые шарики. Их черные головки ярко выделялись на фоне белоснежных сугробов. Перышки на спине у них серые, а грудки – ярко-розовые. Это были снегири. Короткими клювиками снегири вытаскивали из снега веточки рябины и клевали алые ягоды.
Грамматическое задание
- Подчеркните основу первого предложения.
- Разберите по составу слово «головки».
- Придумайте и запишите однокоренные слова к слову «снег».
Диктант № 26 «Как заяц зимой живёт» (59 слов)
Зима. Мороз. Все животные от лютой стужи попрятались. А у зайца норы нет. Зато у него шубка тёплая. Днём заяц спит, а ночью выходит покормиться. В поле лапками снег разроет и травку найдёт. В лесу с кустов молодые веточки обгрызёт или кору погложет. А иногда в деревне из стога сена надёргает и наестся. Так и живёт заяц всю зиму.
В лесу с кустов молодые веточки обгрызёт или кору погложет. А иногда в деревне из стога сена надёргает и наестся. Так и живёт заяц всю зиму.
Грамматическое задание
- Сделайте фонетический разбор слова «заяц».
- Разберите по составу слово «попрятались».
- Обведите в тексте все предлоги.
Диктант № 27 «Снежинки» (59 слов)
Подул холодный ветер. С неба посыпались снежинки. Они кружатся в воздухе и падают на землю. Одна красивее другой! Вот цветок с шестью лепестками. Вот звёздочка с шестью лучами. Вот тонкая пластинка с шестью гранями! В тихую морозную погоду снежинки не теряют красоту. При сильном морозе у них обламываются лучи и грани. Белые цветы и звёзды обращаются в снежную пыль.
Грамматическое задание
- Сделайте синтаксический разбор первого предложения.
- Разберите по составу слово «звёздочка».
- Обведите имена существительные в последнем предложении.

Диктант № 28 «Настоящий друг» (59 слов)
Витя и Илья шли из школы домой. Они спустились к реке. Ночью мороз сковал речку льдом точно зеркальным стеклом. Мальчики решили поиграть на льду. Вдруг под ногами Ильи хрупкий лед треснул. Илья очутился в холодной воде. Бежать за помощью было далеко. Витя нашел длинную палку и протянул ее другу. Илья схватился за палку. Так Витя вытащил товарища на берег.
Грамматическое задание
- Подчеркните в третьем предложении все существительные.
- Проведите синтаксический разбор восьмого предложения.
- Сделайте фонетический разбор слова «Илья».
Диктант № 29 «Волшебный лес» (59 слов)
Отбушевала метель. Лес как в сказке. Ель стоит как молчаливый витязь. На плечах хвойная кольчуга. На колючую хвою села птичка. На снежной поляне маленькие ёлочки. Их совсем занесло снегом. Как они хороши и пригожи! Вьюга сделала деревьям модные прически. Нарядная снежная шапка украсила макушку сосны. Ясная берёзка распустила серебряные от инея косы. Блестит на солнце нежно-розовая тонкая береста берёз.
Нарядная снежная шапка украсила макушку сосны. Ясная берёзка распустила серебряные от инея косы. Блестит на солнце нежно-розовая тонкая береста берёз.
Грамматическое задание
- Сделайте синтаксический разбор пятого предложения.
- Сделайте фонетический разбор слова «инея».
- Обведите слово с разделительным мягким знаком.
Диктант № 30 «Лесная сказка» (59 слов)
Виталик остался ночевать в сторожке. Наступила ночь. Месяц освещал сосны и ели. Снег заискрился блестками. Из окна мальчик смотрел на лесную сказку. Вот белый комок оторвался от ели. Это голодный заяц-беляк поскакал к кустарнику. Он начал строгать зубами кору. К сторожке приблизились лесные великаны. Ветвистые рога украшали головы оленей. Из оленьих ноздрей клубился пар. Лесные красавцы стали есть сено.
Грамматическое задание
- Обозначьте основу первого предложения.
- Разберите по составу слово «лесные».
- Поставьте знак ударения над словами последнего предложения.

Диктант № 31 «Помощь зайцу» (59 слов)
Ребята были в лесу. Под кустом они нашли зайца. Заяц прижался к земле. У зверька была сломана лапка. Гоша принес зайца домой. Мальчик забинтовал зайцу лапку и посадил в ящик. Мама дала зверьку морковку и водички. Заяц прожил в доме месяц. Ножка у зверька срослась. Скоро выпал снег. Наступила зима. Ребята отнесли зайчика в лесок и выпустили на волю.
Грамматическое задание
- Сделайте синтаксический разбор второго предложения.
- Поставьте знак ударения над словами четвёртого предложения.
- Разберите по составу слово «морковку».
Диктант № 32 «Гостья» (60 слов)
На лесной опушке ребята устроили птичью столовую. Ветки ели защищали кормушку от вьюги. Пищу для птиц дети заготовили ещё осенью. Сегодня ребята шли по узкой тропинке навестить друзей. На снегу есть лисьи, заячьи, птичьи следы. В домике уже завтракала синичка. Из чащи прилетела стайка щеглов. На верхушке ёлки появилась белка. Зверёк спрыгнул на птичий домик. Гостья ловко стала объедать ягодки.
Грамматическое задание
- Сделайте синтаксический разбор первого предложения.
- Обведите слово с разделительным мягким знаком.
- Разделите для переноса слова последнего предложения.
Диктант № 33 «Белые фигурки» (60 слов)
В зимнем лесу можно встретить странные белые фигурки. Каждая на что-то похожа. Тут вылез из сугроба лесной человечек в огромной белой папахе. Там на пеньке сидит белочка или зайчик. На камне у речки плачет белая Алёнушка. Она склонила голову на плечо, подпёрла белой ладошкой белую щёчку. Обласкало солнце грустную Алёнушку. С хвойных ресниц её закапали слёзы. Много в лесу диковин!
Грамматическое задание
- В первом предложении подчеркните прилагательные.
- Сделайте фонетический разбор слова «Алёнушка».
- Сделайте синтаксический разбор предпоследнего предложения.

Диктант № 34 «Декабрь» (60 слов)
Декабрь. Мороз. Кругом лежит снег. На деревьях пушистые снежинки. С берёз и осин листья давно опали. Только на дубах желтеют листья. Тишина. Вдруг донеслась весёлая песня. Я оглянулся. На берегу реки сидела птичка. Я шагнул к ней. Птичка кинулась в воду и пропала. Я решил, что она утонула. Но кто это бежит по дну речки? Это оляпка или водяной воробей.
Грамматическое задание
- Разберите третье предложение по членам.
- Разберите по составу слово «осинка».
- Найдите в тексте и подчеркните два слова с безударными гласными в корне, которые можно проверить.
Диктант № 35 «Синичка» (61 слово)
Стояли трескучие морозы. Каждое утро Костя выносил птицам свежий корм. Однажды у кормушки мальчик увидел синичку. Она лежала на снегу. Костя принес птичку домой и поместил ее в клетку. Синичка открыла глазки и ожила. Костя поставил в клетку чашку с пшеном и блюдце с водой. Рядом положил кусочек сала. Синичка прожила у мальчика всю зиму. Весной Костя выпустил птичку на волю.
Она лежала на снегу. Костя принес птичку домой и поместил ее в клетку. Синичка открыла глазки и ожила. Костя поставил в клетку чашку с пшеном и блюдце с водой. Рядом положил кусочек сала. Синичка прожила у мальчика всю зиму. Весной Костя выпустил птичку на волю.
Грамматическое задание
- Подчеркните подлежащее и сказуемое во втором предложении.
- В первом предложении выделите у всех слов окончания.
- Поставьте ударение над словами последнего предложения.
Диктант № 36 «Загадки леса» (62 слова)
Хорошая погодка была в декабре. Лесная дорожка вела нас в лес. Всюду загадки. Под сосной и елью пустые шишки. Здесь крепким клювом поработал дятел. А кто грыз орешки и бросал скорлупки вниз? Это белка. Куда ведут заячьи следы? В осинник. Робкий зверек в белой шубке сидит под осинкой, прижал ушки. На макушку дуба уселся ворон. Под елью большой сугроб. Там берлога медведя.
Грамматическое задание
- Обведите в первом предложении все имена существительные.
- Обозначьте окончания в словах второго предложения.
- Разделите на слоги слова последнего предложения.

Диктант № 37 «Чудесная ёлка» (62 слова)
Мой дед был лесником. Однажды поздним вечером он возвращался домой. Вдруг дед услышал вой волков. Он побежал по дороге. Волки всё ближе. Дед влез на сук ёлки. Волки стали ждать под деревом. Наступила тёмная ночь. Ветром качало ветки. У деда слетела шапка. Голодные волки схватили шапку и разорвали её в клочья. Утром волки скрылись. Дед спустился с дерева и пошёл в сторожку.
Грамматическое задание
- Сделайте фонетический разбор слова «ёлки».
- Проведите синтаксический разбор третьего предложения.
- Разделите для переноса слова последнего предложения.
Диктант № 38 «Зимний сон» (64 слова)
Наступила зима. Все вокруг как будто уснуло. Не слышно птичьего гомона. Только редкие вороны хрипло каркают в парке. Земля укрыта огромным белоснежным покрывалом. Сосны и ели накинули снежные шубы, а тоненькие березы укутали свои веточки серебристым пледом. Тихо вокруг. Не слышно ни звука. Только на полянке видны следы редких прохожих. Речка тоже погрузилась в спячку. Её сковал мороз. Буйные волны замерли под толщей льда.
Земля укрыта огромным белоснежным покрывалом. Сосны и ели накинули снежные шубы, а тоненькие березы укутали свои веточки серебристым пледом. Тихо вокруг. Не слышно ни звука. Только на полянке видны следы редких прохожих. Речка тоже погрузилась в спячку. Её сковал мороз. Буйные волны замерли под толщей льда.
Грамматическое задание
- Подчеркните основу третьего предложения.
- Найдите и обведите разделительный мягкий знак.
- Разберите по составу слово «тоненькие».
Диктант № 39 «Зимний вечер» (64 слова)
Короток зимний вечер. Синий сумрак выполз из леса и повис над сугробами. Резко хрустел под ногами. На звёздном небе появилась луна. Мороз крепчал. Вьюга намела большие сугробы. Деревья и кусты укрылись хлопьями снега. Старые пни надели на головы пушистые шапки. Поздним вечером мы подъехали к сторожке лесника. Маленький домик было чуть видно. Мы затопили печь. Ярко запылал огонь. В избушке стало тепло и уютно.
Грамматическое задание
- Выпишите из текста два слова с безударной гласной, подберите к ним проверочные слова.
- Выпишите из текста два слова с парной согласной, подберите к ним проверочные слова.
- Разделите слова для переноса: звёздном, деревья, маленький.
Диктант № 40 «Новый год» (65 слов)
Скоро наступит новый год. Папа принес домой красивую ёлку. Лесная красавица еще вчера росла в лесу, укрыв снегом свои пушистые ветви. За вечер елочка отогрелась в тепле, расправила иголочки. Мы украсили ёлку. Теперь на веточках покачиваются разноцветные шары. Золотой дождик сверкает в лучах света. А на самую верхушку папа прикрепил большую рубиновую звезду. Стемнело. На нашей елочке зажглись разноцветные лампочки. Вся комната осветилась праздничным светом.
Грамматическое задание
- Найдите в тексте предложения с однородными членами.
- Сделайте фонетический разбор слова «ёлка».
- Подчеркните основу предпоследнего предложения.

Диктант № 41 «Первый снег» (65 слов)
Ночью выпал первый снег. Еще вчера было пасмурно, шел холодный дождь. А небо было затянуто свинцовыми тучами. За ночь все вокруг преобразилось. Дороги и крыши домов укрыты белой пеленой. Снег, будто огромное пушистое одеяло, укрыл всю землю. Деревья похожи на нарядных девушек, которые примерили свои зимние шубки. Мороз – затейник нарисовал на стеклах забавные узоры. Все вокруг стало нарядным и праздничным. Не случайно зиму называют волшебницей.
Грамматическое задание
- Подчеркните основу в последнем предложении.
- Сделайте фонетический разбор слова «вчера».
- Разделите для переноса слова первого предложения.
Диктант № 42 «Метель» (66 слов)
Все началось со снегопада. Снег валил большими белыми хлопьями. За несколько часов выросли огромные сугробы. Мороз крепчал. Пушистые снежинки превратились в острые колючки. Неожиданно подул ветер. По земле закружила поземка. Скоро снежная пелена стала такой густой, что не было видно даже вытянутой руки. Яростный ветер трепал сугробы, разбрасывая снег вокруг, поднимая его в воздух и швыряя обратно на дорогу. Стало темно и очень холодно. Началась метель.
Мороз крепчал. Пушистые снежинки превратились в острые колючки. Неожиданно подул ветер. По земле закружила поземка. Скоро снежная пелена стала такой густой, что не было видно даже вытянутой руки. Яростный ветер трепал сугробы, разбрасывая снег вокруг, поднимая его в воздух и швыряя обратно на дорогу. Стало темно и очень холодно. Началась метель.
Грамматическое задание
- Во втором предложении над существительными напишите их падеж.
- Выпишите из текста все глаголы с приставками.
- В пятом предложении обозначьте второстепенные члены предложения.
Диктант № 43 «Зимой» (68 слов)
В лютый зимний холод всякая птица спешит к жилью человека. Там легче добыть пишу. Голод убивает обычный страх. Люди перестают пугать осторожную лесную живность. Тетерев и куропатка залетают в хранилища с зерном. Русаки ночью постоянно скачут в сад и огород. Ласки устраивают охоту на мышей в подвалах. Беляки прибегают щипать душистое сено из стогов. Однажды через открытую дверь в сторожку влетела синичка. Она стала клевать крошки на столе.
Беляки прибегают щипать душистое сено из стогов. Однажды через открытую дверь в сторожку влетела синичка. Она стала клевать крошки на столе.
Грамматическое задание
- Разберите по составу слово «сторожка».
- Придумайте и запишите однокоренные слова к слову «лесную».
- Выпишите из текста два глагола с приставками, приставки выделите.
Диктант № 44 «Зима» (73 слова)
Земля покрылась белым снежным ковром. Ранним утром я и мой товарищ Андрей отправились в зимний лес. Ночью вьюга занесла все дорожки и тропинки. Мы скользили на лыжах по лесным полянам. Вокруг стояли красивые деревья в зимнем убранстве. Стройные берёзы и осины покрыты пушистым снегом. С ветки летит на шапки мягкий снежок. Мы подъехали к избушке лесника. Она стоит на опушке большой сосновой рощи. Вдруг выглянуло зимнее солнце. Яркий луч его осветил стройные деревья.
Грамматическое задание
- Сделайте синтаксический разбор первого предложения.
- Во втором предложении обведите все имена существительные.
- Сделайте фонетический разбор слова «яркий»

Диктант № 45 «Масленица» (78 слов)
Масленица – самый веселый зимний праздник. Масленицей зиму провожают, а весну встречают. Праздник отмечают в последние дни февраля. Целую неделю устраивают народные гуляния. Все люди выходят на улицу. Дети играют в снежки и катаются на санках. А самое интересное занятие – построить снежную крепость. Теперь нужно запасти много снежков, чтобы не сдать свою крепость команде противников. На Масленицу принято печь блины. Их едят со сметаной, с творогом, с икрой, с вареньем, с медом. Не случайно Масленицу в народе называют Объедухой.
Грамматическое задание
- Найдите в тексте предложения с однородными членами.
- Сделайте фонетический разбор слова «праздник».
- Найдите в тексте однокоренные слова к слову «снег», укажите к каким частям речи они относятся.

Тэги: 3 класс, диктанты, зима, русский язык
О Светлана
Копирайтер. Фрилансер. Мама ребёнка на семейном обучении.
Посмотреть все публикации созданные Светлана →
Словосочетания: признаки, виды связи, синтаксический разбор
- Смысловая и грамматическая связь
- Признаки словосочетания
- Простые и сложные
- Глагольные, именные и наречные
- Типы подчинительной связи в словосочетаниях
- Неделимые словосочетания
- Синтаксический разбор словосочетания
Словосочетание — это сочетание двух или нескольких самостоятельных слов, связанных между собой по смыслу и грамматически. Словосочетание более точно, чем слово, называет предмет, признак, явление или действие:
| слово | словосочетание |
|---|---|
| стол | обеденный стол письменный стол |
| лист | лавровый лист лист малины |
| гулять | гулять в парке гулять с друзьями |
Словосочетание состоит из главного слова и зависимого. Зависимое слово уточняет смысл главного. От главного слова к зависимому всегда можно поставить вопрос:
Зависимое слово уточняет смысл главного. От главного слова к зависимому всегда можно поставить вопрос:
стол (какой?) обеденный,
гулять (с кем?) с друзьями.
Словосочетания прилагательное+существительное
и существительное+существительное
могут быть синонимичными, например:
золотая монета – монета из золота,
меховой воротник – воротник из меха,
бритвенный станок – станок для бритья.
Не являются словосочетаниями:
- Подлежащее и сказуемое:
ветер дует.
- Любые слова, стоящие рядом в предложении, но не связанные между собой по смыслу:
сильный дует.
- Слово с предлогом:
у окна, возле дома.
- Члены предложения, отвечающие на одинаковые вопросы (однородные члены предложения):
(какой?) уставший и (какой?) сонный.
- Устойчивые выражения (фразеологизмы):
спустя рукава.
- Сложные грамматические формы:
будет играть, менее дорогой.
- Обособленные члены в сочетании с определяемым словом:
… ветка, сломанная ветром,…

Смысловая и грамматическая связь
Смысловая связь слов в словосочетаниях устанавливается по вопросам, которые задаются от главного слова к зависимому:
| Смысловая связь | Примеры |
|---|---|
| предмет и его признак | засохший цветок цветок (какой?) засохший |
| действие и предмет | лететь к солнцу лететь (к чему?) к солнцу |
| действие и его признак | заинтересованно разглядывать разглядывать (как?) заинтересованно |
| признак и его степень | по-весеннему свежий свежий (как?) по-весеннему |
Зависимое слово связывается с главным не только по смыслу, но и грамматически. Грамматическая связь слов в словосочетании чаще всего выражается с помощью окончания или с помощью окончания и предлога:
Грамматическая связь слов в словосочетании чаще всего выражается с помощью окончания или с помощью окончания и предлога:
домашние тапочки, тапочки (какие?) домашние;
ходить по лезвию, ходить (по чему?) по лезвию.
Признаки словосочетания
Как и любая единица языка, словосочетания имеют свои признаки:
- Состав словосочетания — два и более самостоятельных слова.
- Смысловое единство этих слов.
- Подчинительная грамматическая связь между компонентами в словосочетании.
Простые и сложные
Все словосочетания состоят из главной и зависимой частей. Например, в словосочетании чистая вода главная часть — вода, зависимая — чистая.
По своему составу словосочетания могут быть простыми и сложными. Простое словосочетание — это словосочетание, состоящее из двух знаменательных слов:
мокрые пятна, всегда весёлый, гулять в саду.
Сложное словосочетание — это словосочетание, состоящее из трёх и более знаменательных слов:
гулять в зимнем саду, стол из красного дерева.
Сложное словосочетание образуется путём распространения компонентов простого словосочетания и, следовательно, может быть разделено на простые:
папин стол из красного дерева – папин стол, стол из дерева, из красного дерева.
Глагольные, именные и наречные
По характеру главного слова словосочетания делятся на глагольные, именные и наречные:
| глагольные | главным словом является глагол | бегать по траве, листать книгу, решать задачу |
|---|---|---|
| именные | главным словом является имя существительное, имя прилагательное, имя числительное или местоимение | свежая краска, очень скучный, пять уроков, кто-то знакомый |
| наречные | главным словом является наречие | навстречу мечте, рядом с домом |
Типы подчинительной связи в словосочетаниях
Слова в словосочетаниях связаны подчинительной связью. В русском языке существует три основных типа (вида) подчинительной связи слов в словосочетаниях: согласование, управление и примыкание.
В русском языке существует три основных типа (вида) подчинительной связи слов в словосочетаниях: согласование, управление и примыкание.
Согласование — это тип подчинительной связи, при котором зависимое слово стоит в том же роде, числе и падеже, что и главное. Например: спелое яблоко — зависимое слово спелое стоит в тех же формах (в единственном числе, среднем роде и именительном падеже), что и главное слово яблоко.
При согласовании с изменением формы главного слова соответственно изменяются и формы зависимого слова. Например:
большая яма (Им. п.), большой ямы (Р. п.), о большой яме (П. п.).
Грамматическая связь при согласовании осуществляется с помощью окончаний. В роли зависимых слов в таких словосочетаниях могут выступать:
- имена прилагательные: надувной шарик;
- местоимения-прилагательные: наши друзья;
- причастия: просмотренный фильм;
- порядковые числительные: второй год.

Управление — это тип подчинительной связи, при котором главное слово управляет
падежной формой зависимого слова. Например:
прочитать книгу (В. п.), журнал (В. п.), страницу (В. п.).
Главное слово прочитать требует
поставить зависимые от него слова — книга, журнал, страница, в винительный падеж.
При управлении с изменением формы главного слова форма зависимого слова не изменяется (зависимое слово остаётся в том же падеже). Например:
прочитать книгу (В. п.), прочитал книгу (В. п.), прочитаю книгу (В. п.).
Грамматическая связь при управлении выражается с помощью окончаний или окончаний и предлогов. Например, в словосочетании
гулять по парку
грамматическая связь передаётся с помощью окончания -у и предлога по.
Примыкание — это тип подчинительной связи, при котором в роли зависимого слова выступают неизменяемые слова (глаголы неопределённой формы, деепричастия, наречия):
учиться читать, бежать прихрамывая, писать грамотно.
При примыкании связь выражается не формами слов, а порядком слов и интонацией.
Неделимые словосочетания
Неделимые словосочетания — это словосочетания, которые в предложениях выполняют роль одного члена предложения:
сломанный ветром, пара часов, два цветка, брат с сестрой.
Синтаксический разбор словосочетания
Порядок разбора словосочетания:
- Назвать главное и зависимое слово. Поставить вопрос от главного слова к зависимому.
- Определить, какой частью речи является главное слово.
- Определить, какой частью речи выражено зависимое слово.
- Назвать средства грамматической связи (выделить окончание у зависимого слова или окончание и предлог).
- Определить вид словосочетания по главному слову.
- Указать тип подчинительной связи (согласование, управление, примыкание).
При письменном разборе словосочетания, над главным словом ставится символ – ×
.
Пример. Образец разбора словосочетания: засохший лист.
Устный разбор.
В словосочетании засохший лист главное слово — лист. Лист (какой?) засохший. Засохший — зависимое слово. Главное слово выражено именем существительным. Зависимое слово выражено именем прилагательным. Зависимое слово связано с главным с помощью окончания -ий. Вид словосочетания по главному слову — именное. Тип подчинительной связи — согласование (зависимое слово согласуется с главным в мужском роде, единственном числе, именительном падеже; при изменении главного слова изменяется и зависимое).
Письменный разбор.
Информация Синонимы: 55 синонимов и антонимов для информации
См. Определение Информировать на Dictionary.com
- Глагол Знания общего
- кратко
- предупредительно
- обучать
- инструктировать
- уведомлять
- рассказывать
- обновлять
- предупреждать
- знакомить
- betray
- blab
- clue
- edify
- endow
- endue
- enlighten
- familiarize
- forewarn
- illuminate
- inspire
- invest
- leak
- level
- post
- relate
- стукачок
- визг
- болтовня
- учить
- подсказка
- зазывать
- заполнить
- дать указатель
- Дайте TIP
- Отдайте
- .
Дайте два цента- Let In on
- Дайте знать
- Сделайте со счетом
- Word
- Покажите Ropes
- . для: информированный / информирующий / информирует
антонимы к слову информировать
НАИБОЛЕЕ АКТУАЛЬНЫЕ
- скрыть
- узнать
- молчать
- скрывать
- 2
- 2обманывать
- хранить в секрете
- вводить в заблуждение
- неясно
- утаивать
- в секрете
Тезаурус 21 века Роже, третье издание Copyright © 2013 by Philip Lief Group.
ПОПРОБУЙТЕ ИСПОЛЬЗОВАТЬ inform
Посмотрите, как выглядит ваше предложение с разными синонимами.
Символы: 0/140
ВИКТОРИНА
Расслабьтесь в шезлонге и пройдите викторину «Слово дня»!
НАЧАТЬ ВИКТОРИНУКак использовать информировать в предложении
Он сообщил им, что является студентом и получил результаты довольно быстро, в течение двух дней, прежде чем сесть в самолет.
ДАЖЕ В САМЫХ ВНИМАТЕЛЬНЫХ ШКОЛАХ БУДЕТ ВСПЫШКА СИ МУХЕРДЖИ СЕНТЯБРЯ 17, 2020FORTUNE
Если бы она была проинформирована о том, что делается с ее фаллопиевыми трубами, она смогла бы справедливо отреагировать на это.
ЖЕНЩИНА В ЛЕДОВОМ ЗАСЛУЖИВАНИИ ГОВОРИТ, ЧТО ЕЕ ФАЛЛОПИЕВУ ТРУБУ УДАЛИЛИ БЕЗ ЕЕ СОГЛАСИЯ NICOLE NAREASE17 СЕНТЯБРЯ 2020 ГОДАVOX
Вместо этого врач сообщил ей, что он также удалил одну из ее фаллопиевых труб и что у нее могут возникнуть трудности с зачатием, если она захочет иметь больше детей.
ЖЕНЩИНА В ЛЕДОВОМ ЗАСЛУЖИВАНИИ УТВЕРЖДАЕТ, ЧТО ЕЕ ФАЛЛОПИЕВУ ТРУБУ УДАЛИЛИ БЕЗ ЕЕ СОГЛАСИЯ 17 СЕНТЯБРЯ 2020 ГОДАVOX
Его всеобъемлющий отчет, выпущенный в 2018 году, послужил основой для моего исследования и приведенного ниже списка.
ХОТИТЕ ЭФФЕКТИВНО БОРАТЬСЯ С ИЗМЕНЕНИЕМ КЛИМАТА? ЗДЕСЬ ПОЖЕРТВОВАТЬ СВОИ ДЕНЬГИ.
СИГАЛ САМУЭЛЬ 17 СЕНТЯБРЯ 2020 ГОДАVOXВскоре престижный колледж Уэлсли сообщил ей, что она включена в список ожидания для поступления.
КОГДА В ЛИНКОЛЬНЕ ХАЙСКОТТЕ-ЛЬЮИССЕ КЛАСС ВНЕЗАПНО СТАЛ КЕРАМИЧЕСКИМ 16 СЕНТЯБРЯ 2020 ГОЛОС САН-ДИЕГО
Эти опасения «не были должным образом рассмотрены», и компания «не проинформировала FAA», говорится в отчете.
АВАРИИ BOEING БЫЛИ «УЖАСНОЙ КУЛЬМИНАЦИЕЙ» МНОЖЕСТВЕННЫХ ОШИБОК, ОТЧЕТ ДОМА SAYSKDUNN6, 16 СЕНТЯБРЯ 2020 г. FORTUNE
Хотя это предназначено для того, чтобы снять часть нагрузки с рекламного бизнеса издателя, онлайн-рынок также информирует обо всех первых- Стратегия партийных данных, сказал он, таким образом, обратная связь с ее контентной стратегией и рекламным бизнесом.
«ОДИН БЕСКОНЕЧНЫЙ ЦИКЛ»: КАК GOLF ИСПОЛЬЗУЕТ СВОЙ НОВЫЙ РОЗНИЧНЫЙ РЫНОК В КАЧЕСТВЕ ПЕРВЫХ ДАННЫХ PLAYKAYLEIGH BARBERS 16 сентября 2020 г.
содержание.«БЕЗ РОСКОШИ ВРЕМЕНИ»: WALL STREET JOURNAL ПРЕДСТАВЛЯЕТ НОВЫЕ ПРОДУКТЫ ДЛЯ БЫСТРОГО РЕКЛАМНОГО РЫНКА LUCINDA SOUTHERN15 сентября 2020 г. .
5 СОВЕТОВ ПО ДОБАВЛЕНИЮ ПОДКЛЮЧЕННОГО ТЕЛЕВИДЕНИЯ В ВАШ ПРАЗДНИК РЕКЛАМНАЯ СТРАТЕГИЯ КОНТЕНТ, СПОНСИРУЕМЫЙ КОНТЕНТОМ: STEELHOUSE 14 СЕНТЯБРЯ 2020 г.SEARCH ENGINE LAND
Бенчик и его команда считают, что матка гудит, чтобы сообщить рабочим пчелам, что она вылупилась.
Стжек и Туты помогают молодым пчелам избегать смертельных дуэлсшаронов Oostekseptepteptepteptepteptepteptepteptepteptepteptepteptepteptepteptepteptepteptepteptepteptepteptepteptepteptepteptepteptepteptepteptepteptepteptepteptepteptepteptepteptepteptepteptepteptepteptepteptepteptept.0012
- bring out
- clue
- come out with
- disclose
- divulge
- enlighten
- familiarize
- fill in
- fix up
- get-together
- habituate
- inform
- intro
- introduce
- сбить с ног
- сообщить
- познакомить
- уведомить
- опубликовать
- подарить
- раскрыть
- рассказать
- warn
 Дайте два цента
Дайте два цента
 СИГАЛ САМУЭЛЬ 17 СЕНТЯБРЯ 2020 ГОДАVOX
СИГАЛ САМУЭЛЬ 17 СЕНТЯБРЯ 2020 ГОДАVOX содержание.
содержание.- accustoming
- advising
- apprising
- bringing out
- cluing
- coming out with
- disclosing
- divulging
- enlightening
- familiarizing
- filling in
- fixing up
- getting together
- привыкание
- информирование
- знакомство
- знакомство
- сбивание с ног
- letting knowing
- making familiar
- notifying
- posting
- presenting
- revealing
- telling
- warning
- acquaint
- advance
- advise
- announce
- apprise
- ballyhoo
- beat the drum для
- счет
- герб
- повышение
- создание
- распространение
- связь
- declare
- disclose
- display
- divulge
- drum
- endorse
- exhibit
- expose
- flaunt
- get on soapbox for
- hard sell
- herald
- hype
- inform
- make a шаг
- сообщить
- уведомить
- шаг
- заглушить
- пресс-агент
- объявить
- продвигать
- promulgate
- puff
- push
- put on the map
- reveal
- show
- soft sell
- splash
- sponsor
- spot
- tout
- uncover
- unmask
- acquaints
- advances
- советует
- объявляет
- сообщает
- шумит
- бьет в барабан
- купюры
- гербы
- boosts
- builds up
- circularizes
- communicates
- declares
- discloses
- displays
- divulges
- drums
- endorses
- exhibits
- exposes
- flaunts
- gets on soapbox for
- hard sell
- предвещает
- шумиха
- сообщает
- делает шаг
- делает известным
- уведомляет
- pitches
- plugs
- press agent
- proclaims
- promotes
- promulgates
- puffs
- pushes
- puts on the map
- reveals
- shows
- soft sell
- splashes
- sponsors
- spots
- рекламирует
- раскрывает
- разоблачает
- знакомит
- сообщает
- подсказка
- подсказка в
- Заполните
- Дайте слово
- Информация
- Keep Oped
- Lay Out
- Let In On
- Сделайте известные
- Notify
- POST
- . на
- отчет
- показать
- рассказать
- напомнить
- обновить
- предупредить
 на
на- предостеречь 9019 предостережения
- 2s9 charges
- commends
- counsels
- directs
- dissuades
- encourages
- enjoins
- exhorts
- forewarns
- gives a pointer
- gives a tip
- guides
- instructs
- kibitzes
- levels with
- движется
- думает
- указывает
- проповедует
- готовит
- предписывает
- подсказывает
- Поставки ошибки в уш
- Поставки в два цента
- Рекомендуются
- Управление
- Предлагает
- TOUT Группа.
Промежуточные филаменты — Клетка
Промежуточные филаменты имеют диаметр около 10 нм, что является промежуточным между диаметрами двух других основных элементов цитоскелета, актиновых филаментов (около 7 нм) и микротрубочек (около 25 нм).
В отличие от актиновых филаментов и микротрубочек промежуточные филаменты не принимают непосредственного участия в клеточных движениях. Вместо этого они, по-видимому, играют в основном структурную роль, обеспечивая механическую прочность клеток и тканей.Белки промежуточных филаментов
В то время как актиновые филаменты и микротрубочки представляют собой полимеры белков одного типа (актина и тубулина соответственно), промежуточные филаменты состоят из множества белков, которые экспрессируются в разных типах клеток. Было идентифицировано более 50 различных белков промежуточных филаментов, которые были разделены на шесть групп на основе сходства между их аминокислотными последовательностями (4). Типы I и II состоят из двух групп кератинов, каждая из которых состоит примерно из 15 различных белков, которые экспрессируются в эпителиальных клетках. Каждый тип эпителиальных клеток синтезирует по крайней мере один тип I (кислый) и один тип II (нейтральный/основной) кератин, которые сополимеризуются с образованием филаментов.
Некоторые кератины типа I и II (называемые твердые кератины ) используются для изготовления таких структур, как волосы, ногти и рога. Другие кератины I и II типов ( мягкие кератины ) в изобилии присутствуют в цитоплазме эпителиальных клеток, при этом разные кератины экспрессируются в различных типах дифференцированных клеток.Таблица 11.1
Белки промежуточных филаментов.
Белки промежуточных филаментов типа III включают виментин , который содержится в различных типах клеток, включая фибробласты, гладкомышечные клетки и лейкоциты. Другой белок типа III, десмин , специфически экспрессируется в мышечных клетках, где он соединяет Z-диски отдельных сократительных элементов. Третий белок промежуточных филаментов III типа специфически экспрессируется в глиальных клетках, а четвертый экспрессируется в нейронах периферической нервной системы.
Белки промежуточных филаментов типа IV включают три белка нейрофиламентов (NF) (обозначенные как NF-L, NF-M и NF-H для l light, m edium и h eavy, соответственно).
). Эти белки образуют основные промежуточные филаменты многих типов зрелых нейронов. Их особенно много в аксонах двигательных нейронов, и считается, что они играют решающую роль в поддержании этих длинных тонких отростков, длина которых может достигать более метра. Другой белок типа IV (α-интернексин) экспрессируется на более ранней стадии развития нейронов, до экспрессии белков нейрофиламентов. Единственный белок промежуточных филаментов VI типа (нестин) экспрессируется еще раньше в процессе развития нейронов, в стволовых клетках центральной нервной системы.Белки промежуточных филаментов типа V представляют собой ядерные ламины, обнаруженные в большинстве эукариотических клеток. Ядерные ламины являются не частью цитоскелета, а компонентами ядерной оболочки (see). Они также отличаются от других белков промежуточных филаментов тем, что они собираются с образованием ортогональной сети, лежащей в основе ядерной мембраны.
Несмотря на значительные различия в размерах и аминокислотной последовательности, различные белки промежуточных филаментов имеют общую структурную организацию (4).
Все белки промежуточных филаментов имеют центральный α-спиральный стержневой домен примерно из 310 аминокислот (350 аминокислот в ядерных ламинах). Этот центральный стержневой домен окружен амино- и карбоксиконцевыми доменами, которые варьируют среди различных белков промежуточных филаментов по размеру, последовательности и вторичной структуре. Как обсуждается далее, α-спиральный стержневой домен играет центральную роль в сборке филаментов, в то время как вариабельные головные и хвостовые домены, по-видимому, определяют специфические функции различных белков промежуточных филаментов.Рисунок 11.31
Структура белков промежуточных филаментов. Белки промежуточных филаментов содержат центральный α-спиральный стержневой домен примерно из 310 аминокислот (350 аминокислот в ядерных ламинах). N-концевой головной и С-концевой хвостовой домены различаются по (подробнее…)
Сборка промежуточных филаментов
Первая стадия сборки филаментов – образование димеров, в которых закручены центральные стержневые домены двух полипептидных цепей вокруг друг друга в спирально-спиральной структуре, похожей на структуру, образованную тяжелыми цепями миозина II (11).
Затем димеры объединяются в шахматном порядке антипараллельно с образованием тетрамеров, которые могут собираться конец к концу, образуя протофиламенты. Последний промежуточный филамент содержит примерно восемь протофиламентов, намотанных друг вокруг друга в виде веревкообразной структуры. Поскольку они собраны из антипараллельных тетрамеров, оба конца промежуточных филаментов эквивалентны. Следовательно, в отличие от актиновых филаментов и микротрубочек промежуточные филаменты неполярны; у них нет отчетливых плюсовых и минусовых концов.Рисунок 11.32
Сборка промежуточных нитей. Центральные стержневые домены двух полипептидов наматываются друг на друга в спирально-спиральной структуре, образуя димеры. Затем димеры связываются в шахматном порядке антипараллельно с образованием тетрамеров. Тетрамеры связываются конец к концу (подробнее…)
Сборка филаментов требует взаимодействия между определенными типами белков промежуточных филаментов. Например, кератиновые филаменты всегда собираются из гетеродимеров, содержащих один полипептид типа I и один полипептид типа II.
Напротив, белки типа III могут собираться в филаменты, содержащие только один полипептид (например, виментин) или состоящие из двух разных белков типа III (например, виментин плюс десмин). Однако белки типа III не образуют сополимеров с кератинами. Среди белков типа IV α-интернексин может сам собираться в филаменты, тогда как три белка нейрофиламентов сополимеризуются с образованием гетерополимеров.Промежуточные филаменты, как правило, более стабильны, чем актиновые филаменты или микротрубочки, и не демонстрируют динамического поведения, связанного с этими другими элементами цитоскелета (например, беговая дорожка актиновых филаментов, показанная на рис. ). Однако белки промежуточных филаментов часто модифицируются фосфорилированием, которое может регулировать их сборку и разборку внутри клетки. Наиболее ярким примером является фосфорилирование ядерных ламинов (см.), что приводит к разборке ядерной ламины и разрушению ядерной оболочки во время митоза. Цитоплазматические промежуточные филаменты, такие как виментин, также фосфорилируются при митозе, что может приводить к их разборке и реорганизации в делящихся клетках.
Внутриклеточная организация промежуточных филаментов
Промежуточные филаменты образуют сложную сеть в цитоплазме большинства клеток, простирающуюся от кольца, окружающего ядро, до плазматической мембраны (). И кератиновые, и виментиновые филаменты прикрепляются к ядерной оболочке, по-видимому, служащие для позиционирования и закрепления ядра внутри клетки. Кроме того, промежуточные филаменты могут связываться не только с плазматической мембраной, но и с другими элементами цитоскелета, актиновыми филаментами и микротрубочками. Таким образом, промежуточные филаменты образуют каркас, объединяющий компоненты цитоскелета и организующий внутреннюю структуру клетки.
Рисунок 11.33
Внутриклеточная организация кератиновых филаментов. Микрофотография эпителиальных клеток, окрашенных флуоресцентными антителами к кератину (зеленый). Ядра окрашены в синий цвет. Кератиновые филаменты простираются от кольца, окружающего ядро, до плазматической мембраны. (Нэнси (подробнее.
..)Кератиновые филаменты эпителиальных клеток прочно прикреплены к плазматической мембране в двух областях специализированных клеточных контактов, десмосомах и полудесмосомах (). Десмосомы представляют собой соединения между соседними клетками, в которых происходит межклеточный контакт. опосредованы трансмембранными белками, родственными кадгеринам. На цитоплазматической стороне десмосомы связаны с характерной плотной бляшкой внутриклеточных белков, к которой прикреплены кератиновые филаменты. Эти присоединения опосредуются десмоплакином, членом семейства белков, называемых плакины , которые связывают промежуточные филаменты и связывают их с другими клеточными структурами. Гемидесмосомы представляют собой морфологически сходные соединения между эпителиальными клетками и подлежащей соединительной тканью, в которых кератиновые филаменты связаны с интегринами различными членами семейства плакинов (например, плектинами). Таким образом, десмосомы и гемидесмосомы прикрепляют промежуточные филаменты к областям межклеточных и клеточно-субстратных контактов, соответственно, аналогично прикреплению актинового цитоскелета к плазматической мембране в местах слипчивых соединений и фокальных адгезий.
Важно отметить, что кератиновые филаменты, прикрепленные к обеим сторонам десмосом, служат механической связью между соседними клетками в эпителиальном слое, тем самым обеспечивая механическую стабильность всей ткани.Рисунок 11.34
Прикрепление промежуточных филаментов к десмосомам и гемидесмосомам. (A) Электронная микрофотография, иллюстрирующая кератиновые филаменты (стрелки), прикрепленные к плотным бляшкам внутриклеточного белка по обеим сторонам десмосомы. (B) Схема десмосомы. Промежуточные (подробнее…)
Помимо связывания промежуточных филаментов с клеточными соединениями, некоторые плакины связывают промежуточные филаменты с другими элементами цитоскелета. Плектин, например, связывает актиновые филаменты и микротрубочки в дополнение к промежуточным филаментам, так что он может создавать мосты между этими компонентами цитоскелета (14). Считается, что эти мостики к промежуточным филаментам скрепляют и стабилизируют актиновые филаменты и микротрубочки, повышая тем самым механическую стабильность клетки.
Рисунок 11.35
Электронная микрофотография пектиновых мостиков между промежуточными филаментами и микротрубочками. Микрофотография фибробласта, окрашенного антителами против плектина. Микрофотография искусственно окрашена, чтобы показать плектин (зеленый), антитела против плектина (подробнее…)
Два типа промежуточных филаментов, десмин и нейрофиламенты, играют особую роль в мышечных и нервных клетках соответственно. Десмин связывает отдельные актин-миозиновые ансамбли мышечных клеток как друг с другом, так и с плазматической мембраной, тем самым связывая действия отдельных сократительных элементов. Нейрофиламенты являются основными промежуточными филаментами в большинстве зрелых нейронов. Их особенно много в длинных аксонах двигательных нейронов, где они, по-видимому, прикреплены к актиновым филаментам и микротрубочкам нейронами семейства плакинов. Считается, что нейрофиламенты играют важную роль в обеспечении механической поддержки и стабилизации других элементов цитоскелета в этих длинных тонких отростках нервных клеток.
Функции кератинов и нейрофиламентов: заболевания кожи и нервной системы
Хотя долгое время считалось, что промежуточные филаменты обеспечивают структурную поддержку клетки, только недавно были получены прямые доказательства их функции. Некоторые клетки в культуре не производят белков промежуточных филаментов, что указывает на то, что эти белки не требуются для роста клеток in vitro . Аналогично, инъекция культивируемым клеткам антител против виментина разрушает сети промежуточных филаментов, не влияя на рост или движение клеток. Поэтому считалось, что промежуточные филаменты наиболее необходимы для укрепления цитоскелета клеток в тканях многоклеточных организмов, где они подвергаются различным механическим воздействиям, не влияющим на клетки в изолированной среде культуральной чашки.
Экспериментальные доказательства такой in vivo роли промежуточных филаментов были впервые получены в 1991 г. в ходе исследований в лаборатории Элейн Фукс. Эти исследователи использовали трансгенных мышей для исследования in vivo эффектов экспрессии мутанта с делецией кератина, кодирующего укороченный полипептид, который нарушал образование нормальных кератиновых филаментов (4).
Этот мутантный ген кератина вводили трансгенным мышам, где он экспрессировался в базальных клетках эпидермиса и нарушал формирование нормального кератинового цитоскелета. Это привело к развитию тяжелых кожных аномалий, в том числе волдырей из-за лизиса эпидермальных клеток после легкой механической травмы, такой как растирание кожи. Таким образом, кожные аномалии этих трансгенных мышей напрямую подтверждают предполагаемую роль кератинов в обеспечении механической прочности эпителиальных клеток в тканях.Рисунок 11.36
Экспериментальная демонстрация функции кератина. Плазмиду, кодирующую мутантный кератин, препятствующий нормальной сборке кератиновых филаментов, микроинъецировали в один пронуклеус оплодотворенной яйцеклетки. Затем микроинъецированные эмбрионы были перенесены в (подробнее…)
. Эти эксперименты также указали на молекулярную основу генетического заболевания человека, простого буллезного эпидермолиза (EBS). Подобно трансгенным мышам, экспрессирующим мутантные гены кератина, у пациентов с этим заболеванием появляются волдыри на коже в результате лизиса клеток после незначительной травмы.
Это сходство вызвало исследования генов кератина у пациентов с СЭБ, что привело к демонстрации того, что СЭБ вызывается мутациями гена кератина, которые мешают нормальной сборке кератиновых филаментов. Таким образом, как экспериментальные исследования на трансгенных мышах, так и молекулярный анализ генетического заболевания человека продемонстрировали роль кератинов в обеспечении устойчивости клеток кожи к механическим воздействиям. Продолжающиеся исследования показали, что мутации в других кератинах ответственны за несколько других наследственных заболеваний кожи, которые также характеризуются аномальной хрупкостью эпидермальных клеток.Другие исследования на трансгенных мышах выявили причастность аномалий нейрофиламентов к заболеваниям двигательных нейронов, в частности к боковому амиотрофическому склерозу (БАС). БАС, известный как болезнь Лу Герига и заболевание, поразившее известного физика Стивена Хокинга, возникает в результате прогрессирующей потери двигательных нейронов, что, в свою очередь, приводит к атрофии мышц, параличу и, в конечном итоге, к смерти.
БАС и другие типы заболеваний двигательных нейронов характеризуются накоплением и аномальной сборкой нейрофиламентов, что позволяет предположить, что аномалии нейрофиламентов могут способствовать этим патологиям. В соответствии с этой возможностью было обнаружено, что сверхэкспрессия NF-L или NF-H у трансгенных мышей приводит к развитию состояния, сходного с БАС. Хотя задействованный механизм еще предстоит понять, эти эксперименты ясно указывают на участие нейрофиламентов в патогенезе заболевания двигательных нейронов.Терминология аутригеров и подъемных кранов
Специально для аутригеров, крановых площадок и фундаментных опор.
Допустимое давление на грунт: Максимально допустимое давление, обычно выражаемое в фунтах на квадратный фут (psf) или Паскалях (Па), которое может воздействовать на опорную поверхность. Это значение может быть равно несущей способности грунта, деленной на подходящий коэффициент безопасности, или может быть меньшим значением, ограниченным другими соображениями, такими как прочность подземных труб.
См. Давление на грунт; см. Несущая способность почвы.Директор A/D: (директор по сборке/демонтажу) означает лицо, которое соответствует требованиям настоящего подраздела для директора A/D, независимо от официальной должности лица или того, является ли это лицо неуправленческим или управленческим персоналом. (Источник: Глоссарий общих терминов по кранам и такелажу . Специализированные носители и такелажный фонд)
Сборка/разборка: Означает сборку и/или разборку оборудования, подпадающего под действие настоящего стандарта. Что касается башенных кранов, термин «монтаж» заменяет термин «сборка», а термин «демонтаж» заменяет термин «демонтаж». Независимо от того, монтируется ли кран изначально на полную высоту или поднимается поэтапно, процесс увеличения высоты крана является процессом возведения. (Источник: Глоссарий общих терминов по кранам и такелажу . Specialized Carriers & Rigging Foundation)
Среднее давление на грунт (AGBP): Сила/площадь = AGBP.
Среднее давление, оказываемое на землю под определенной площадью.Напряжение при изгибе: Напряжение, возникающее в несущем элементе из-за изгибающего действия этого элемента, в отличие от действия на растяжение или сжатие. (Источник: Глоссарий общих терминов по кранам и такелажу . Фонд Specialized Carriers & Rigging Foundation)
Блокировка: Материал, используемый для поддержки оборудования или компонента и распределения нагрузки на землю. Обычно он используется для поддержки секций решетчатой стрелы во время сборки/разборки, а также под поплавками аутригеров и стабилизаторов. Также называется dunnage, cribbing, пусковыми площадками, и циновками.
Выносная опора бампера: Гидравлический цилиндр или ручной домкрат, расположенный на переднем бампере тележки автокрана для обеспечения дополнительной устойчивости и увеличения рабочего диапазона крана над передней частью тележки. Также: Пятая выносная опора.
(Источник: Глоссарий общих терминов по кранам и такелажу . Specialized Carriers & Rigging Foundation)Композит: Композитный материал (также называемый композиционным материалом или сокращенно композитным, что является общепринятым названием) изготовлен из двух или более составляющих материалов со значительно различающимися физическими или химическими свойствами, которые при комбинированные, производят материал с характеристиками, отличными от отдельных компонентов. Отдельные компоненты остаются отдельными и отчетливыми в готовой структуре, что отличает композиты от смесей и твердых растворов. Запатентованный материал DICA для колодок выносных опор SafetyTech и домкратов RoadWarrior RV не является композитным. Все подушки кранов и выносных опор FiberMax являются композитными материалами.
Коврик для крана: Совокупность конструктивных элементов, установленных или соединенных и используемых для поддержки гусеничных кранов и/или оборудования для распределения нагрузки и устойчивости.
Крановая подушка: Изготовленные цельные решения, используемые для поддержки выносных опор под оборудованием с выносными опорами для распределения нагрузки и устойчивости. См. крановые подушки DICA.
Ребрышки: Блокирующие материалы, которые используются для увеличения опорной площади и высоты. См. пластиковые подпорки DICA.
Критический подъем: Операция подъема краном, связанная с исключительным уровнем риска из-за таких факторов, как вес груза, высота подъема, процедурные сложности или близость к ситуационным опасностям. Критические подъемы могут быть идентифицированы условиями, превышающими определенный процент номинальной грузоподъемности крана; однако может быть задействовано много более сложных вопросов. (Источник: Глоссарий общих терминов по кранам и такелажу . Фонд Specialized Carriers & Rigging Foundation)
Степень раздавливания: Допустимое давление, на которое рассчитана подушка выносной опоры, подушка выносной опоры крана, опора или подушка крана, в зависимости от прочности материала.
Прогиб: Изгибание опорных материалов при приложении прижимной силы. См. Прижимная сила.
Вышка: Подъемное устройство, состоящее из вертикальной мачты, закрепленной наверху растяжками или скобами, с использованием подъемных механизмов и рабочих канатов. Вышка может использовать или не использовать стрелу. (Источник: Глоссарий общих терминов по кранам и такелажу . Specialized Carriers & Rigging Foundation)
Вышка: Операция по изменению угла наклона стрелы путем изменения длины подвесных канатов стрелы. (Источник: Глоссарий общих терминов по кранам и такелажу . Specialized Carriers & Rigging Foundation)
Водоизмещение: Разница между горизонтальными положениями опоры выносной опоры без нагрузки по сравнению с горизонтальным положением опоры опоры при полной нагрузке.
Прижимная сила: Сила, создаваемая в направлении вниз от выносных опор через поплавок выносных опор и на опоры выносных опор.
Dunnage: Сыпучие материалы, используемые для поддержки, разделения и защиты оборудования, деталей и принадлежностей во время хранения и стабилизации.
Эффективная площадь опоры: Площадь полного контакта, эффективно распределяющая приложенную нагрузку на подстилающую поверхность.
Модуль упругости (жесткость при изгибе): Отношение напряжения к деформации, определяющее склонность материала к изгибу.
Поплавок: Жесткий чашеобразный конструктивный элемент, который крепится к внешнему концу выносной опоры мобильного крана для обеспечения некоторого распределения нагрузки от выносной опоры на опорную поверхность. Поплавки предоставляются производителем крана. Также обычно называется поплавком аутригера, подушкой аутригера, поддоном аутригера, ногой или подушкой.
Давление на грунт (GBP): Давление, обычно выражаемое в фунтах на квадратный фут (psf) или Паскалях (Па), которое кран оказывает на опорную поверхность.
См. Допустимое давление на грунт.Несущая способность грунта (GBC): Прочность грунта или его способность выдерживать давление.
Состояние грунта: Способность грунта поддерживать оборудование (включая уклон, уплотнение и твердость) (29 CFR 1926.1402)
Заземляющий мат: Заземляющее устройство, используемое, когда кран месте, которое находится вблизи опасности поражения электрическим током. Оборудование помещается на этот мат и приклеивается к нему. Зона заземляющего мата обычно огорожена забором, чтобы персонал не наступал на мат и не спускался с него во время работы машины. (Источник: Глоссарий общих терминов по кранам и такелажу . Specialized Carriers & Rigging Foundation)
Заземляющий стержень: Заземляющее устройство, используемое, когда кран будет работать в относительно фиксированном месте, близком к опасности поражения электрическим током. Заземляющий провод подсоединяется к стержню и к машине, при этом стержень размещается как можно ближе к машине.
(Источник: Глоссарий общих терминов по кранам и такелажу . Фонд Specialized Carriers & Rigging Foundation)Домкрат: Механическое или гидравлическое устройство для подъема тяжестей на короткие расстояния. СВЯЗАННЫЕ: Домкрат для выносных опор. (Источник: Глоссарий общих терминов по кранам и такелажным работам . Фонд Specialized Carriers & Rigging Foundation). (Источник: Глоссарий общих терминов по кранам и такелажу . Фонд Specialized Carriers & Rigging Foundation)
Грузоподъемность: Максимальный вес брутто-груза, который производитель крана определил для безопасной работы крана при определенных условиях противовеса, уровня, положения выносных опор, длины стрелы, радиуса и т. д. Грузоподъемность публикуется в таблицах, известных как диаграммы грузоподъемности. Грузоподъемность НЕ является максимальным грузом, который кран может поднять до того, как он опрокинется или рухнет. (Источник: Глоссарий общих терминов по кранам и такелажу .
Фонд Specialized Carriers & Rigging Foundation)Мат: Отдельные брусья, скрепленные вместе в блоки, стальные пластины или плетеные проволоки, размещенные под гусеницами, колесами или подушками выносных опор для предотвращения опускания за счет увеличения площади опоры, соприкасающейся с землей, что снижает опорное давление. (Источник: Глоссарий общих терминов по кранам и такелажу . Specialized Carriers & Rigging Foundation)
Максимальная нагрузка: Самый тяжелый груз, который, как показывает таблица грузоподъемности крана, он способен поднять в данной конфигурации. (Источник: Глоссарий общих терминов по кранам и такелажу . Specialized Carriers & Rigging Foundation)
Ограничение максимального радиуса нагрузки: Наибольшее расстояние от центральной линии поворота крана (радиус), на котором кран с заданной длиной стрелы может поднять любой груз. (Источник: Глоссарий общих терминов по кранам и такелажу .
Specialized Carriers & Rigging Foundation)Максимальное усилие реакции выносных опор: Максимальный вес, который оборудование может оказывать через свои выносные опоры.
Максимальный радиус: Наибольшее расстояние от центральной линии вращения крана (радиус), на котором может быть поднят максимальный груз в пределах номинальной грузоподъемности крана. (Источник: Глоссарий общих терминов по кранам и такелажу . Специализированные носители и такелажный фонд) См. «Максимальная нагрузка на аутригеры».
Минимальный радиус: Кратчайшее расстояние от центральной линии вращения крана (радиус), на котором кран с заданной длиной стрелы может поднять любой груз. (Источник: Глоссарий общих терминов по кранам и такелажу . Specialized Carriers & Rigging Foundation)
На выносных опорах: (1) Состояние, при котором кран работает со всеми выносными опорами, полностью выдвинутыми и установленными для выравнивания корпуса крана, а его шины или гусеницы не касаются земли.
(2) Номинальные грузоподъемности крана, которые применимы, когда кран работает со всеми выносными опорами, полностью выдвинутыми и установленными так, чтобы выровнять корпус крана с его шинами или гусеницами, не касающимися земли. Связанный: На резине. (Источник: Глоссарий общих терминов по кранам и такелажу . Specialized Carriers & Rigging Foundation)На резине: (1) Состояние, при котором кран работает с любой из своих шин, соприкасающихся с землей. (2) Номинальная грузоподъемность крана, применимая при работе крана с любой из его шин, соприкасающихся с землей. Также: На шинах. Связанный: На выносных опорах. (Источник: Глоссарий общих терминов по кранам и такелажу . Фонд Specialized Carriers & Rigging Foundation)
Выносная опора: Балка, выходящая из нижней части крана для повышения устойчивости крана. Балки могут быть либо выдвижными/убирающимися, либо фиксированной длины, и обычно используют какой-либо тип прокладок для распределения нагрузок на поверхность земли.
(Источник: Глоссарий общих терминов по кранам и такелажу . Specialized Carriers & Rigging Foundation)Балка аутригера: Часть системы аутригеров, которая проходит горизонтально от коробки аутригеров для поддержки вертикального домкрата аутригеров. (Источник: Глоссарий общих терминов по кранам и такелажу . Specialized Carriers & Rigging Foundation)
Домкрат аутригера: Гидравлический цилиндр на балке аутригера, который выдвигается вертикально для подъема и опускания крана. (Источник: Глоссарий общих терминов по кранам и такелажу . Специализированные носители и такелажный фонд)
Площадка/поплавок/ножка выносной опоры: Металлическая пластинчатая конструкция, поставляемая производителем оборудования, распределяющая общую нагрузку от нижележащая блокировка и земная поверхность. Типичные формы круглые, квадратные, прямоугольные или восьмиугольные. СМОТРИТЕ: плавайте.
Подушка выносной опоры: Деревянный, металлический или синтетический конструктивный элемент, который размещается на опорной поверхности и на который опирается плавучее устройство крана, используемое для распределения нагрузки на выносную опору на большую площадь.
См. опорные подушки DICA.Система штифтов выносных опор: Доступна гидравлическая система для облегчения снятия коробки выносных опор с помощью гидравлических цилиндров, используемых вместо стандартных монтажных штифтов коробки выносных опор. (Источник: Глоссарий общих терминов по кранам и такелажу . Specialized Carriers & Rigging Foundation)
Подушка: Уплотненный грунт, бетон, деревянные брусья или маты или стальные пластины, собранные в систему для поддержки крана с минимальной осадкой. (Источник: Глоссарий общих терминов по кранам и такелажу . Специализированные носители и такелажный фонд)
Понтон: (1) Чашеобразная конструкция, которая крепится к нижней части вертикального домкрата с выносной опорой для распределения нагрузки по опорной поверхности. СМОТРИТЕ: плавайте. (2.) Большие стальные подушки, обычно изготавливаемые для поддержки грузов крана или выносных опор на мягком грунте.
Понтоны — это стальной эквивалент деревянных матов крана. (Источник: Глоссарий общих терминов по кранам и такелажу . Specialized Carriers & Rigging Foundation)Контрольное испытание: Неразрушающее испытание, при котором к системе или компоненту применяется пробная нагрузка. (Источник: Глоссарий общих терминов по кранам и такелажу . Специализированные носители и такелажный фонд)
Номинальная грузоподъемность: Максимальная рабочая нагрузка на кран или компонент такелажа, разрешенная производителем при определенных условиях работы. ТАКЖЕ: предел рабочей нагрузки. (Источник: Глоссарий общих терминов по кранам и такелажу . Specialized Carriers & Rigging Foundation)
Винтовой домкрат: Резьбовой вертикальный опорный цилиндр, обычно используемый на выносных опорах или кольцевых навесных устройствах для регулировки уровня. СВЯЗАННЫЙ: Джек.
Площадка винтового домкрата: Конструкция чашеобразного типа, прикрепленная к нижней части винтового домкрата для распределения нагрузки на землю, аналогичная опорной стойке.
(Источник: Глоссарий общих терминов по кранам и такелажу . Specialized Carriers & Rigging Foundation)Начальник участка: Осуществляет надзор за рабочим участком, на котором используется кран, и работами, которые выполняются на этом участке. (Источник: Глоссарий общих терминов по кранам и такелажу . Специализированные носители и такелажное основание)
Стабилизатор: Выдвижные или фиксированные элементы, прикрепленные к монтажному основанию для повышения устойчивости крана, но которые могут не иметь возможности снять весь вес с колес гусениц. (Источник: ASME B30.5 & B30.22 )
Вспомогательные материалы: OSHA 1926.1402(a)(2) «Вспомогательные материалы» означают блоки, маты, опоры, поддерживающие тележки (на болотах/заболоченных территориях) или аналогичные материалов или устройств.
Выход: Точка, в которой материал начинает деформироваться, изгибаться или ломаться.
Как только происходит текучесть, материалы не возвращаются к своей первоначальной форме, а свойства материала изменяются навсегда.Информация об очистке компьютера и шаги
Обновлено: 02.08.2020, автор: Computer Hope
Чистка компьютера, компонентов и периферийных устройств помогает поддерживать все в хорошем рабочем состоянии, помогает предотвратить распространение микробов и обеспечивает надлежащий поток воздуха. На картинке показан хороший пример того, насколько грязной может быть внутренняя часть корпуса вашего компьютера. Глядя на это изображение, сразу становится очевидным, что вся пыль и грязь будут препятствовать надлежащему потоку воздуха и даже могут помешать работе вентилятора.
- Как часто нужно чистить компьютер?
- Общие советы по уборке.
- Инструменты для чистки.
- Очистка корпуса.
- Очистка дисководов CD-ROM, DVD и других дисков.
- Очистка CD, DVD и других дисков.
- Очистка жесткого диска.
- Чистка наушников.
- Чистка клавиатуры.
- Очистка ЖК/светодиодов.
- Очистка ЭЛТ-монитора.
- Очистка материнской платы.
- Чистка мышей.
- Очистка принтера.
- Очистка сканера.
- Как почистить смартфон.
- Как почистить ноутбук.
- Как почистить системный вентилятор компьютера.
- Различные этапы очистки.
Как часто нужно чистить компьютер?
Частота очистки компьютера зависит от различных факторов. Чтобы помочь вам определить, как часто вам нужно чистить компьютер, мы составили контрольный список ниже. Отметьте каждый из приведенных ниже флажков, которые относятся к состоянию вашего компьютера, чтобы определить, как часто нужно чистить компьютер.
Где находится компьютер? В домашних условиях. В чистом офисе. В строительстве или промышленности. В школьной среде. Компьютерная среда Держите кошку или собаку в одном здании с компьютером. Дым в том же здании, что и компьютер. Дым рядом с компьютером. Компьютер на полу. В комнате с компьютером есть ковер. Ешьте или пейте за компьютером. Кто им пользуется? Взрослый (18 лет и старше). Молодые люди (в возрасте 10–18 лет) пользуются компьютером. Подростки (младше 10 лет) пользуются компьютером. Компьютером пользуется несколько человек. С тем, что проверено выше, очищайте компьютер каждый месяц.
Советы по общей чистке
Ниже приведены рекомендации по очистке любых компонентов компьютера или периферийных устройств, а также советы по поддержанию чистоты компьютера.
- Никогда не распыляйте жидкость на какие-либо компоненты компьютера. Если необходим спрей, распылите жидкость на ткань.
- Вы можете использовать пылесос для сбора грязи, пыли или волос вокруг компьютера. Однако не используйте пылесос внутри вашего компьютера, так как он создает статическое электричество, которое может повредить ваш компьютер. Если вам нужно использовать пылесос внутри вашего компьютера, используйте портативный пылесос с питанием от батареи или попробуйте сжатый воздух.
- При очистке компонента или компьютера выключите его перед очисткой.
- Будьте осторожны при использовании чистящих растворителей; у некоторых людей бывают аллергические реакции на химические вещества в чистящих растворителях, а некоторые растворители могут даже повредить корпус. Старайтесь всегда использовать воду или сильно разбавленный растворитель.
- При чистке следите за тем, чтобы случайно не отрегулировать какие-либо ручки или элементы управления. Кроме того, при очистке задней панели компьютера, если что-либо подключено, не отсоединяйте вилки.
- При очистке вентиляторов, особенно небольших, держите вентилятор или поместите что-нибудь между лопастями вентилятора, чтобы он не вращался. Распыление сжатого воздуха на вентилятор или очистка вентилятора пылесосом может привести к повреждению или возникновению обратного напряжения.
- Никогда не ешьте и не пейте рядом с компьютером.
- Ограничьте курение возле компьютера.
Инструменты для чистки
Хотя средства для чистки компьютеров доступны, вы также можете использовать предметы домашнего обихода для очистки компьютера и его периферийных устройств. Ниже приведен список предметов, которые могут вам понадобиться или которые вы хотите использовать при очистке компьютера.
- Салфетка . Хлопчатобумажная ткань лучше всего подходит для протирания компонентов компьютера. Бумажные полотенца можно использовать с большинством оборудования, но мы всегда рекомендуем использовать ткань, когда это возможно. Однако при очистке таких компонентов, как корпус, дисковод, мышь и клавиатура, используйте только ткань. Не используйте ткань для очистки каких-либо схем, таких как оперативная память или материнская плата.
- Вода или медицинский спирт — При увлажнении ткани лучше всего использовать воду или медицинский спирт. Другие растворители могут быть вредны для пластика, используемого в вашем компьютере.
- Портативный пылесос . Высасывание пыли, грязи, волос, частиц сигарет и других частиц из компьютера может быть одним из лучших способов очистки компьютера. Однако не используйте пылесос, который подключается к стене, так как он создает много статического электричества, которое может повредить ваш компьютер.
- Ватные тампоны — Ватные тампоны, смоченные спиртом или водой, — отличный инструмент для протирки труднодоступных мест на клавиатуре, мыши и других местах.
- Тампоны из пенопласта — По возможности лучше использовать безворсовые тампоны, например тампоны из пенопласта.
Кончик
Список других инструментов, которые должен иметь каждый технический специалист, см. на нашей странице компьютерных инструментов.
Очистка корпуса
Почему? Очистка корпуса позволяет сохранить внешний вид компьютера как нового. Во время очистки, если вы видите вентиляционные прорези, их можно очистить или очистить, чтобы поддерживать постоянный поток воздуха в компьютер и охлаждать все компоненты.
Процедура: Пластиковый корпус, в котором находятся компоненты ПК, можно протирать слегка влажной безворсовой тканью. Для удаления трудновыводимых пятен добавьте в ткань немного бытового моющего средства. Не используйте чистящие растворители для пластика.
Убедитесь, что на всех вентиляционных и вентиляционных отверстиях нет волос и ворсинок, протирая отверстия и вентиляционные отверстия тканью. Также полезно пропылесосить все отверстия, вентиляционные отверстия и щели на компьютере.
При очистке внешних вентиляционных отверстий компьютера безопасно использовать стандартный пылесос.Если вам нужны инструкции по очистке внутренних частей компьютера, см. раздел о очистке материнской платы.
- Компьютерный корпус (шасси) помощь и поддержка.
Очистка дисководов CD-ROM, DVD и других дисков
Почему? Грязный дисковод для компакт-дисков или другие дисководы могут вызывать ошибки чтения при чтении дисков. Эти ошибки чтения могут вызвать проблемы с установкой программного обеспечения или проблемы во время работы программы.
Процедура: Для очистки дисковода компакт-дисков рекомендуется приобрести средство для очистки компакт-дисков у местного продавца компьютеров. Использование чистящего средства для компакт-дисков должно в достаточной степени очистить лазер для компакт-дисков от пыли, грязи и волос.
- Где купить запчасти для компьютерной техники.
Вы также можете использовать ткань, смоченную водой, для очистки лотка, который выдвигается из дисковода.
Однако убедитесь, что после очистки лоток полностью высох, прежде чем вставлять лоток обратно в дисковод.См. рекомендации по очистке дисков для дальнейших действий по очистке каждого из ваших компакт-дисков.
- Компакт-диск справки и поддержки.
Очистка CD, DVD и других дисков
Почему? Грязные компакт-диски могут вызвать ошибки чтения или привести к тому, что компакт-диски вообще не будут работать.
Процедура: Используйте набор для чистки или влажную чистую хлопчатобумажную ткань для очистки компакт-дисков, DVD-дисков и других дисков. При очистке диска протирайте дорожки, начиная с середины диска и продвигаясь к внешней стороне. Никогда не протирайте дорожками; это может привести к появлению новых царапин на диске.
Кончик
Если вещество на компакт-диске нельзя удалить с помощью воды, можно также использовать чистый спирт.
Очистка жесткого диска
Почему? Жесткие диски компьютера не могут быть физически очищены.
Однако их можно очистить с помощью программных утилит, чтобы они работали быстро и эффективно. Использование этих утилит предотвращает замедление работы жесткого диска.Процедура: См. наш основной раздел по устранению неполадок, чтобы узнать, какие действия для вашей операционной системы помогут повысить производительность компьютера.
- Как очистить жесткий диск компьютера.
- Мой компьютер работает медленно, что я могу сделать, чтобы это исправить?
- Помощь и поддержка жесткого диска.
Чистка наушников
Почему? Наушники и гарнитуры могут использоваться разными людьми, и, возможно, их необходимо часто чистить, чтобы предотвратить распространение микробов и вшей.
Процедура: Если наушники пластиковые или виниловые, смочите ткань теплой водой и протрите головку и наушники наушников.
Примечание
Библиотечные или школьные наушники нельзя чистить никакими дезинфицирующими средствами или чистящими растворителями, так как у некоторых людей могут возникнуть аллергические реакции на содержащиеся в них химические вещества.
Наушники с подушечками также имеют возможность замены подушечек. Замена этих подушечек также поможет содержать наушники в чистоте.
И, наконец, по поводу наушников, распространяющих педикулеза. Если разные учащиеся используют одни и те же наушники, пусть учащиеся используют свои собственные наушники, надевают на наушники пакеты или используют наушники, которые можно протирать теплой водой после каждого использования.
Чистка клавиатуры
Эти действия предназначены для очистки клавиатуры настольного компьютера. О ноутбуках см.: Как почистить ноутбук.
Пыль, грязь и бактерии
Клавиатура компьютера обычно является наиболее зараженным микробами предметом в вашем доме или офисе. На клавиатуре может быть даже больше бактерий, чем на сиденье унитаза. Очистка помогает удалить любые опасные бактерии, исправить или предотвратить залипание клавиш и обеспечить правильную работу клавиатуры.
Процедура: Перед очисткой клавиатуры сначала выключите компьютер или, если вы используете USB-клавиатуру, отсоедините ее от компьютера.
Неотключение клавиатуры может вызвать другие проблемы с компьютером, так как вы можете нажимать клавиши, которые заставят компьютер выполнять задачу, которую вы не хотите, чтобы он выполнял.Многие чистят клавиатуру, переворачивая ее вверх дном и встряхивая. Более эффективным методом является использование сжатого воздуха. Сжатый воздух — это сжатый воздух, находящийся в баллоне с очень длинным соплом. Чтобы очистить клавиатуру сжатым воздухом, направьте его между клавишами и сдуйте всю пыль и мусор. Можно также использовать пылесос, но убедитесь, что на клавиатуре нет выпадающих клавиш, которые могут быть засосаны пылесосом.
Если вы хотите более тщательно очистить клавиатуру, снимите клавиши с клавиатуры.
- Как убрать клавиши на клавиатуре.
После удаления пыли, грязи и волос. Распылите дезинфицирующее средство на ткань или используйте дезинфицирующие салфетки и протрите каждую из клавиш на клавиатуре. Как упоминалось в наших общих советах по очистке, никогда не распыляйте жидкость на клавиатуру.
Вещество пролилось на клавиатуру
Если на клавиатуру что-то пролилось (например, газированные напитки, кола, пепси, кока-кола, пиво, вино, кофе и молоко), несоблюдение надлежащих мер может привести к поломке клавиатуры.
Процедура: Ниже приведены рекомендации, которые помогут предотвратить поломку клавиатуры после того, как что-то попало на клавиши.
Если что-то пролилось на клавиатуру, немедленно выключите компьютер или, по крайней мере, отсоедините клавиатуру от компьютера. После этого переверните клавиатуру, чтобы вещество не проникло в цепи. Перевернув клавиатуру, потрясите ее над поверхностью, которую можно очистить позже. Пока все еще вверх дном, используйте ткань, чтобы начать чистить клавиши. После очистки клавиатуры оставьте ее в перевернутом виде как минимум на два дня, чтобы она высохла. После высыхания продолжайте очищать клавиатуру оставшимся веществом.
Если после очистки клавиши клавиатуры залипают, снимите клавиши и очистите область под клавишами и нижнюю часть клавиши.
Наконец, если клавиатура все еще работает, но остается грязной или липкой, перед тем, как выбросить клавиатуру, в крайнем случае попробуйте помыть клавиатуру в посудомоечной машине.
- Можно ли мыть клавиатуру компьютера в посудомоечной машине?
Если после выполнения всех вышеперечисленных действий клавиатура по-прежнему не работает, рекомендуем приобрести новую клавиатуру.
- Помощь и поддержка клавиатуры.
Очистка LCD/LED
Почему? Грязь, пыль и отпечатки пальцев могут затруднить чтение информации с экрана компьютера.
Процедура: В отличие от ЭЛТ-монитора, ЖК- или светодиодный монитор не является стеклянным и требует специальных процедур очистки.
При очистке ЖК-экрана или светодиодного экрана важно помнить, что нельзя распылять какие-либо жидкости непосредственно на экран. Аккуратно нажимайте во время очистки и не используйте бумажное полотенце, так как оно может поцарапать экран.
Для очистки ЖК-дисплея или светодиодного экрана используйте непрочную ткань из микрофибры, мягкую хлопчатобумажную ткань или тряпку Swiffer. Если сухая ткань не полностью очищает экран, можно нанести на ткань медицинский спирт и протереть экран влажной тканью. Медицинский спирт используется для очистки ЖК- и светодиодных мониторов перед отправкой с завода.
- Монитор, ЖК-дисплей и дисплей, помощь и поддержка.
Очистка ЭЛТ-монитора
Кончик
Этот раздел предназначен для ЭЛТ-мониторов. Если у вас монитор с плоским экраном, см. раздел о чистке LCD/LED.
Почему? Грязь, пыль и отпечатки пальцев могут затруднить чтение информации с экрана компьютера.
Процедура: Стеклянный экран монитора можно чистить обычным бытовым средством для мытья стекол. Отсоедините шнур питания монитора и распылите чистящее средство на безворсовую ткань, чтобы предотвратить попадание жидкости на какие-либо компоненты внутри монитора.
Пропылесосьте всю пыль, осевшую на верхней части монитора, и убедитесь, что книги или бумаги не закрывают вентиляционные отверстия. Засоренные вентиляционные отверстия монитора могут привести к его перегреву или даже возгоранию.Осторожность
Мы рекомендуем использовать только ткань, смоченную водой, при очистке нестеклянных мониторов или экранов с антибликовым покрытием. Использование обычных бытовых очистителей для стекол на специальных экранах, особенно очистителей с аммиаком, может удалить антибликовую защиту или другие специальные поверхности.
Другие хорошие чистящие растворы
- Полотенца из микрофибры
- Тряпки Swiffer
Сопутствующая информация
- Монитор, ЖК-дисплей и дисплей, помощь и поддержка.
Чистка материнской платы
Почему? Пыль и особенно частицы сигаретного дыма могут накапливаться и вызывать коррозию схем, вызывая различные проблемы, такие как зависание компьютера.
Осторожность
Находясь внутри компьютера, соблюдайте необходимые меры предосторожности от электростатического разряда и старайтесь не отсоединять кабели и другие соединения.
Процедура: Мы рекомендуем при очистке материнской платы от пыли, грязи или волос использовать сжатый воздух. При использовании сжатого воздуха держите его вертикально, чтобы химические вещества не вытекли из контейнера, что может привести к повреждению материнской платы или других компонентов. Кроме того, убедитесь, что при использовании сжатого воздуха вы всегда сдуваете пыль или грязь с материнской платы или из корпуса.
Еще одна хорошая альтернатива сжатому воздуху — портативный пылесос с батарейным питанием. Портативные пылесосы могут полностью удалить пыль, грязь и волосы с материнской платы и предотвратить их попадание в корпус.
- Помощь и поддержка материнской платы.
Предупреждение
Никогда не используйте пылесос с электроприводом, поскольку он может вызвать статическое электричество, которое может повредить компьютер.
При использовании пылесоса с батарейным питанием держите его на расстоянии нескольких дюймов от компонентов, чтобы предотвратить повреждение и засасывание чего-либо в пылесос (например, перемычек).Кончик
При очистке внутренней части корпуса также обратите внимание на вентиляторы и радиаторы. Пыль, грязь и волосы могут больше всего скапливаться вокруг этих компонентов.
Чистка мышей
Оптическая или лазерная мышь
Почему? Из-за грязной оптической или лазерной мыши курсор мыши может двигаться с трудом или хаотично.
Процедура: Используйте баллончик со сжатым воздухом, предназначенный для использования с электронным оборудованием, распыляя его вокруг оптического датчика в нижней части мыши. Обдув нижней части мыши воздухом удаляет грязь, пыль, волосы и другие препятствия, блокирующие оптический датчик.
Не используйте какие-либо чистящие химикаты и не протирайте тканью непосредственно оптический датчик, так как это может поцарапать или повредить оптический датчик.
Оптико-механическая мышь (шариковая мышь)
Почему? Грязная оптико-механическая мышь (мышь с шариком) может затруднить перемещение мыши и вызвать странное движение мыши.
Процедура: Чтобы очистить ролики оптико-механической мыши, необходимо сначала снять нижнюю крышку мыши. Для этого осмотрите нижнюю часть мыши, чтобы увидеть, в какую сторону вращать крышку. Как вы можете видеть на рисунке ниже, крышку мыши нужно двигать против часовой стрелки. Поместите два пальца на крышку мыши, нажмите и поверните ее в направлении стрелок.
После того, как крышка повернется примерно на дюйм, поверните мышь в нормальное положение, прикрывая нижнюю часть мыши одной рукой. Затем дно должно отпасть, включая шарик мыши. Если крышка не падает, попробуйте осторожно встряхнуть мышь.
После снятия нижней крышки и шарика внутри мыши должны быть видны три ролика. Используйте ватную палочку, палец или ноготь, чтобы удалить любые вещества с роликов.
Обычно в середине валика есть небольшая линия волос и грязи. Удалите как можно больше этого вещества.После того, как вы удалили как можно больше грязи и волос, установите шарик обратно в мышь и снова наденьте крышку.
Если с мышью по-прежнему возникают те же проблемы, повторите описанный выше процесс. Если после нескольких попыток с мышью все еще возникают те же проблемы, у вашей мыши есть другие проблемы с оборудованием, и ее следует заменить.
Примечание
Протирка коврика для мыши влажной тканью также может помочь улучшить движения компьютерной мыши.
Все типы мышей
Почему? Для поддержания мыши в чистоте и обеззараживании.
Процедура: Используйте ткань, смоченную спиртом или теплой водой, и протрите поверхность мыши и каждую из ее кнопок.
- Помощь и поддержка мыши и сенсорной панели.
Очистка принтера
Почему? Очистка внешних поверхностей принтера помогает поддерживать его внешний вид в хорошем состоянии, а если принтер используется несколькими людьми, очищает его от микробов.
Процедура: Прежде чем чистить принтер, обязательно выключите его. Смочите ткань водой или спиртом и протрите корпус и каждую кнопку или ручку на принтере. Как упоминалось ранее, никогда не распыляйте жидкость непосредственно на принтер.
Почему? Для обеспечения бесперебойной работы некоторых принтеров требуется внутренняя очистка.
- Справка и поддержка принтера.
Очистка сканера
Почему? Планшетные сканеры обычно загрязняются пылью, отпечатками пальцев и волосами. Если сканер загрязнен, изображения могут иметь искажения.
Процедура: Очистите поверхность планшетного сканера, распылив средство для мытья окон на бумажное полотенце или хлопчатобумажную ткань, и протрите стекло, пока оно не станет чистым. Как упоминалось ранее, никогда не распыляйте жидкость непосредственно на компонент.
Это же полотенце или хлопчатобумажная ткань можно использовать для очистки сканера снаружи.
- Помощь и поддержка сканера.
Прочие этапы очистки
Ниже приведен список разного компьютерного оборудования, которое сегодня редко используется, но хранится на этой странице людьми, работающими на старых компьютерах и нуждающихся в очистке этих устройств.
Очистка дисковода гибких дисков
Почему? Грязные головки чтения/записи на дисководе гибких дисков могут вызвать ошибки в процессе чтения или записи.
Процедуры: Дисковод гибких дисков очищается двумя разными способами. Первый способ очистки флоппи-дисковода — приобрести в местном розничном магазине комплект, предназначенный для очистки головок чтения/записи на флоппи-дисководе.
Второй способ очистки дисковода рекомендуется только опытным пользователям компьютеров. Откройте корпус дисковода гибких дисков и протрите головки чтения/записи безворсовой губкой, смоченной в чистом спирте или трихлорэтане. При выполнении этих шагов будьте предельно осторожны при очистке головок, чтобы убедиться, что вы не заблокируете их смещение, в результате чего флоппи-дисковод не будет работать.
Чтобы предотвратить проблемы с выравниванием головы, используйте похлопывающие движения, слегка накладывая тампон на голову и удаляя его, не выполняйте движения тампоном из стороны в сторону.- Помощь и поддержка дисковода гибких дисков.
Очистка пилота Palm
Почему? Грязные сенсорные экраны могут затруднить навигацию.
Процедура: Для очистки экрана PalmPilot используйте мягкую ткань, смоченную спиртом, и протрите экран и корпус PalmPilot. Не рекомендуется использовать средство для мытья стекол, так как со временем оно может повредить пластик.
Очистка SuperDisk и LS-120
Почему? Очистка SuperDisk и LS-120 предотвращает загрязнение головок накопителей.
Процедура: Приобретите комплект для очистки SuperDisk, доступный через Imation. Использование любого другого метода аннулирует гарантию на ваш диск.
Модуляция иммунных реакций размером и формой частиц
Введение
Клетки иммунной системы могут морфологически изменять свою плазматическую мембрану для поглощения и выведения чужеродных частиц, попадающих в организм, таких как бактерии и грибы при инфекции, и частиц из эндогенных источников, таких как тела апоптотических клеток (1).
Поглощение клетками внеклеточных частиц необходимо для многих клеточных функций и, среди прочего, играет важную роль в иммунной системе и ремоделировании тканей. Патогены или частицы, попадающие в организм, поглощаются иммунными фагоцитами путем взаимодействия с рецепторами на их поверхности. Этот процесс необходим для очистки организма от инфекционных микробов (например, бактерий, грибков), стареющих (раковых) клеток и неорганических частиц с помощью иммунных фагоцитов (1). Эндоцитоз — это общий термин для различных типов поглощения более мелких частиц, в то время как более крупные частицы поглощаются с помощью другого механизма, называемого фагоцитозом. В этом обзоре мы будем определять эндоцитоз как поглощение частиц размером <0,5 мкм, а фагоцитоз — как клеточное поглощение частиц размером более 0,5 мкм. Эндоцитарные везикулы могут иметь везикулярную, колбовидную или тубулярную морфологию (2). Наиболее понятная форма эндоцитоза опосредована клатрином, белком оболочки на цитоплазматической стороне мембраны, но существуют и другие белки оболочки (например, кавеолин), и эндоцитоз, по-видимому, не всегда зависит от белков оболочки (2). Из-за ограничений по размеру решетки клатрина клатрин предпочитает управлять поглощением мелких частиц (диаметром <0,1 мкм). Мембранные инвагинации, происходящие из складок мембраны и богатых актином расширений мембраны, называемых ламеллиподиями, образуют фагоцитарные чашечки, которые при запечатывании приводят к образованию крупных связанных с мембраной вакуолей размером в микрометр, называемых фагосомами (рецептор-опосредованный процесс) или макропиносомами (рецептор-независимый процесс). ) (2, 3). Фагоцитоз обычно опосредован актиновым цитоскелетом, который обеспечивает значительную силу для преодоления физических ограничений, необходимых для обертывания мембраны более крупных частиц (рис. 1). Фагоцитоз отличается от поглощения более мелких частиц эндоцитозом, так как он не зависит от белков оболочки, таких как клатрин и кавеолин, а эндоцитоз имеет меньшее участие актинового цитоскелета (1, 2). Для полимеризации актина фагоцитоз требует активности Rho-GTPases и киназ семейства фосфоинозитид-3-киназ (PI3K) (2), и это особенно необходимо для фагоцитоза более крупных частиц (> 5 мкм) (4, 5). В этом обзоре основное внимание уделяется модельным частицам микронного диапазона и грибковым патогенам, поскольку влияние морфологии на клеточное поглощение лучше всего понятно для этих фагоцитарных грузов.Рисунок 1 Поглощение антигенных частиц иммунными фагоцитами. (A) Дендритные клетки (ДК), макрофаги (МФ) и нейтрофилы (НЭ) иммунной системы сталкиваются с частицами с различными физическими свойствами (размером, формой, жесткостью, шероховатостью поверхности). NE может высвобождать внеклеточные ловушки (NET) для захвата крупных патогенов, таких как гифальные грибы. Макрофаги могут подвергаться перепрограммированию в провоспалительные (М1) или противовоспалительные (М2) фенотипы. (B) Частицы > 0,5 мкм поглощаются фагоцитозом; более мелкие частицы путем эндоцитоза, который может быть опосредован клатрином или кавеолином. F-актиновый цитоскелет важен для обертывания мембраны фагоцитарной чашечки. Слияние с лизосомами, содержащими литические ферменты, является последней стадией эндо/фагосомного созревания.
Основными иммунными фагоцитами являются макрофаги, нейтрофилы и дендритные клетки (ДК) (1). Нейтрофилы являются наиболее распространенными фагоцитами в организме, способными вызывать окислительные и неокислительные микробицидные реакции, такие как секреция антибактериальных белков и улавливание патогенов в нейтрофильных внеклеточных ловушках (НЭТ) высвобождаемой ДНК (рис. 1) (6). Макрофаги обнаруживаются во всех тканях и способны устранять инфекции путем фагоцитоза и запускать воспалительные реакции. Они более приспособлены для фагоцитоза меньших морфологических состояний грибов; более крупные гифы или споры иногда не могут быть поглощены макрофагами (7–9). Макрофаги могут поляризоваться в провоспалительные (М1) фенотипы, если они активируются провоспалительными стимулами, такими как интерферон (IFN)-γ, гранулоцитарно-макрофагальный колониестимулирующий фактор (GM-CSF) или микробными стимулами, такими как липополисахарид (LPS). 10). Альтернативно, макрофаги могут поляризоваться в противовоспалительные гомеостатические (М2) фенотипы, индуцированные противовоспалительными цитокинами интерлейкином (ИЛ)-4 или ИЛ-13 (10).
Сдвиг от М2 к М1-подобному фенотипу связан с большей микробицидной активностью макрофагов (10). Подобно нейтрофилам и макрофагам, дендритные клетки (ДК) также берут образцы крови и периферических тканей организма на патогены и апоптотические/некротические клетки (11). Однако, в отличие от нейтрофилов и макрофагов, ДК могут эффективно мигрировать в лимфатические узлы после фагоцитоза патогенов. В лимфатических узлах ДК могут представлять антигенные пептиды, полученные из проглоченных патогенных белков, на своей клеточной поверхности в главном комплексе гистосовместимости (MHC) класса II наивным CD4-положительным «хелперным» Т-клеткам (Th) и, в процессе, называемом перекрестным -представление MHC класса I наивным CD8-положительным «киллерным» Т-клеткам (11, 12). (Перекрестная) презентация антигена DCs необходима для активации наивных Т-клеток и вызывает адаптивный иммунный ответ против вторжения патогенов и рака.Поскольку удаление различных патогенов (грибков, бактерий, вирусов, гельминтов) и злокачественных клеток требует различных иммунных реакций, иммунная система выработала множество рецепторов, которые распознают различные патогены, называемых рецепторами распознавания патогенов (PRR).
Эти PRR распознают так называемые микробно-ассоциированные молекулярные паттерны (MAMP), которые представляют собой структуры, присутствующие на микробах, но не в клетках-хозяевах, и молекулярные паттерны, связанные с опасностью (DAMP), которые присутствуют на поврежденных, но не (или только в небольших количествах) на здоровых клетках-хозяевах (13). Многие PRR, но не все, также могут запускать эндоцитоз и/или фагоцитоз. Например, β-глюканы и хитины клеточной стенки грибов запускают как распознавание антигена, так и фагоцитоз, связываясь с лектинами C-типа dectin-1 и рецептором маннозы (CD206) на поверхности DCs и макрофагов. Основным классом PRR являются Toll-подобные рецепторы (TLR), которые связываются со многими различными MAMP, такими как TLR4, который связывается с LPS. Вовлечение различных рецепторов, таких как dectin-1 и TLR, позволяет иммунным фагоцитам различать различные морфологические состояния грибов (14, 15). Например, по сравнению с гифами набухшая форма конидий патогенного гриба Aspergillus fumigatus имеет повышенный уровень экспонированных на поверхности β-глюканов и вызывает более сильные воспалительные реакции (16).Вероятно, поскольку разные патогены имеют разный размер и задействуют разные фагоцитарные PRR, это может вызывать разные типы фагоцитоза. Например, большие гифы плесени Aspergillus fumigatus поглощаются фагоцитозом, опосредованным Fc-рецептором, тогда как более мелкие споры той же плесени поглощаются посредством маннозного рецептора (10). Фагоцитоз, опосредованный Fc-рецептором, запускает так называемую модель захвата с застежкой-молнией, характеризующуюся плотно прилегающим, подобным застежке-молнией, продвижением мембраны и разветвленного актинового цитоскелета по поверхности частицы; эффективность этого процесса зависит от жесткости частиц, размера, плотности антител на поверхности частиц (17–19) и расстояние между мембраной фагоцита и поверхностью частицы (ограничено размером комплекса Fc-рецептор-лиганд) (20). Менее жесткие апоптотические клетки также поглощаются моделью «молния» (3), но с меньшей эффективностью, чем более жесткие мишени, такие как бактерии и дрожжи (17).
Напротив, фагоцитоз с помощью рецептора комплемента 3 (CR3) происходит через хлопчатобумажных складок мембраны, организованных линейным актином, меньшими выпячиваниями мембраны и «погружением» фагосом в цитоплазму (обзор в (3)). Недавние данные показывают, что поглощение, опосредованное Fc и комплементом, может быть переплетено, и оба могут зависеть от закрепления актина 9.0649 через интегрин α M β 2 на переднем крае ложноножек на фагоцитарной чашечке (17, 21).Частицы, которые слишком велики для поглощения иммунными фагоцитами, приводят к остановке фагоцитарного процесса, и это называется фрустрированным фагоцитозом (22, 23). Такой неудовлетворенный фагоцитоз может привести к так называемой парадигме волокон, которая определяет основу вредного воздействия длинных (> 15 мкм), тонких, не поддающихся биологическому разложению волокон, таких как асбест в легких (22). Здесь альвеолярные макрофаги могут лишь частично поглощать волокно, что приводит к высвобождению литических ферментов и активных форм кислорода (АФК) во внеклеточную среду (22).
Это высвобождение этого содержимого может привести к воспалению и повреждению тканей и, в конечном итоге, вызвать хроническое воспаление, фиброз и онкогенез (22, 24).Фагоцитоз и эндоцитоз, таким образом, зависят от типа задействованного PRR и фагоцитарного рецептора, а также от геометрии и механических свойств частицы (23, 25–34). Как морфология частиц влияет на их поглощение, хорошо видно из экспериментальных (23, 25–33) и теоретических исследований (34–41). Цель этого обзора — сделать общие выводы о том, как геометрия и механические свойства влияют на эндоцитоз и фагоцитоз и последующее эндо/фагосомальное созревание и иммунные реакции, а также о том, как эти знания можно использовать при разработке вакцин на основе частиц. Поэтому мы сравнили как теоретические, так и экспериментальные исследования с модельными частицами и патогенами разного размера и формы. Однако выводы из этих сравнений следует интерпретировать с осторожностью, потому что способы поглощения могут сильно различаться среди типов клеток, и прямое сравнение между эндоцитарными и фагоцитарными грузами и методами не всегда оправдано.
Размер и форма частиц: теоретические исследования и моделирование
Теоретические подходы широко использовались для прогнозирования способов поглощения различных частиц разных размеров и форм (дополнительная таблица 1) (35–37, 41). Сфероидные частицы можно описать по соотношению сторон (AR), которое представляет собой отношение между осью эллипсоида (ось a, см. рис. 2) и осью сферы (ось b): сплюснутые дискообразные частицы (AR<1), сферы (AR=1) и вытянутые частицы (AR>1). Вытянутые частицы с высоким AR (AR>2) также называют эллипсоидными или стержнеобразными частицами. Монте-Карло (35) и крупнозернистая (39) при моделировании сравнивались три режима поглощения сплюснутых частиц и эллипсоидов: сначала кончик (мембрана сначала обращена к наибольшей кривизне вытянутых частиц или наименьшей кривизне поверхностей сплюснутых частиц), укладка (мембрана сначала обращена к наименьшей кривизне вытянутых частиц или наибольшей кривизне поверхностей) сплюснутых частиц) и наклонные (главные оси неперпендикулярны мембране) (рис.
2). На основании этих моделей было предсказано, что, по крайней мере, для эндоцитоза мелких частиц размером от 25 до 100 нм, сплюснутые частицы с низким AR (AR <0,5), вероятно, будут поглощаться в режиме захвата кончиком вперед, например, лицом к лицу. мембрана со стороной наименьшей кривизны (35, 39) (рис. 3). Точно так же вытянутые частицы с большей вероятностью будут поглощаться в режиме укладки, таким образом, они также обращены к мембране стороной с наименьшей кривизной (39). Другие компьютерные исследования также предсказали, что вытянутые частицы с большей вероятностью будут поглощаться 90 649 через 90 650 режим откладывания (35, 37) и что поглощение вытянутых частиц кончиком вперед будет медленнее, чем поглощение кончиком вперед сплюснутых частиц (34). . Однако заворачивание может сопровождаться переориентацией частицы. Например, в нескольких теоретических исследованиях было предсказано, что в то время как вытянутые частицы прикрепляются к мембране в режиме укладки, при более высоких силах адгезии это состояние становится нестабильным, и частица переориентируется в сторону глубоко завернутого механизма захвата кончиком вперед (42). , 43).Рисунок 2 Сферические, сплюснутые и вытянутые частицы. (A) Сфероидные частицы можно описать по их аспектному отношению (AR), которое представляет собой отношение между осью эллипсоида (ось a) и сферической осью (ось b) частицы: сплюснутые дискообразные частицы (AR< 1), сферы (AR=1) и вытянутые частицы (AR>1). Частицы могут поглощаться различными способами: сначала кончиком (мембрана сначала обращена к наибольшей кривизне вытянутых частиц или к поверхности с наименьшей кривизной сплюснутых частиц) или укладыванием (мембрана сначала обращена к наименьшей кривизне вытянутых частиц или к поверхности с наибольшей кривизной сплюснутых частиц). ). (B) Моделирование поглощения сферических, вытянутых, сплюснутых и соответствующих несферических частиц (кубических, стержневидных и дискообразных) обычно предсказывает, что сферические частицы поглощаются лучше, чем вытянутые и сплюснутые частицы. Обычно прогнозируется, что вытянутые частицы наиболее эффективно проглатываются в режиме укладки, тогда как сплюснутые частицы лучше всего проглатываются в режиме кончиком вперед.
Подробности; см. текст.Рис. 3 Эффективность поглощения, определяемая физическими свойствами частиц: теоретические предсказания и экспериментальные данные. Геометрия частиц при фагоцитозе. Размер, соотношение сторон (AR), угол начального контакта между основной осью и мембраной (см. Рисунок 2), жесткость частиц и плотность лиганда определяют эффективность эндо/фагоцитоза. Резюме теоретических предсказаний и экспериментальных результатов. Экспериментальные данные о том, как жесткость частиц влияет на поглощение, противоречивы.
Энергия поглощения зависит от свойств изгиба мембраны и силы взаимодействия с рецепторами, а общий эндоцитоз более эффективен (более низкая потребность в свободной энергии) с более сильной адгезией и большей площадью контакта (44). Моделирование опосредованного клатрином эндоцитоза частиц размером ~100 нм предсказало, что эффективность поглощения определяется жесткостью мембраны при изгибе, которая пропорциональна жесткости поверхности плазмы, поскольку это определяет образование связей рецептор-лиганд (37).
Компьютерное моделирование также показало, что мембрана может полностью обернуть частицы с 0,68Моделирование показывает, что для частиц разной формы и объема мембрана должна преодолевать разные пороги свободной энергии для обертывания частиц (38, 45).
Предполагается, что из-за повышенных затрат энергии на обертывание сильно изогнутых кончиков эллипсоидные частицы проглатываются менее эффективно, чем сферические частицы аналогичного размера (45). Из-за более низкой локальной кривизны более крупных частиц прогнозируется, что сплющенные частицы меньшего объема будут поглощаться менее эффективно, чем вытянутые частицы меньшего объема, но сплюснутые частицы большего объема поглощаются более эффективно, чем вытянутые частицы большего объема (38). Прогнозируется, что для эндоцитоза смоделированное клатрин-опосредованное поглощение будет максимально эффективным для более крупных сферических частиц с оптимальными размерами, близкими к пределу размера относительно жесткой решетки клатрина (более эффективное поглощение для 80 нм, чем для 40 нм или 160 нм). размера частиц (37)), и это также зависит от покрытия лигандом и жесткости моделируемых частиц (36, 37). Наконец, теоретическое исследование предсказало, что кривизна кончика частицы имеет значение, поскольку было предсказано, что частицы с высоким отношением длины к ширине и круглыми кончиками будут входить в через режим укладки , в то время как частицы с меньшим соотношением сторон и плоскими кончиками могут поглощаться более эффективно в режиме кончик-сначала (46). Это, кажется, противоречит другому исследованию компьютерного моделирования, в котором было предсказано, что, учитывая, что жесткость частиц остается неизменной, скорость поглощения и изгиб мембраны одинаковы для кубов и сфер, дисков и сплюснутых частиц, а также стержней и вытянутых частиц (рис. 2B). ) (39). В целом, к поглощению таких частиц разной формы, по-видимому, применимо одно общее правило: частицы большего объема могут поглощаться более эффективно, чем частицы меньшего объема, но с таким же AR (рис. 3), из-за меньшей кривизны мембраны и более высокой количество взаимодействий рецептор-лиганд (38).Что касается механических свойств частиц, компьютерное моделирование предсказывает, что поглощение эластичных (мягких) частиц менее эффективно, чем более жестких частиц, поскольку деформации частиц приводят к более высоким энергетическим барьерам для обертывания мембраны, а для поглощения более мягких частиц может потребоваться больше рецепторы для преодоления этого энергетического барьера (рис.
3) (39). Предполагается, что поглощение сплюснутых частиц будет наименее чувствительным к этой эластичности, если частицы расположены кончиком вперед, потому что они имеют большую площадь контакта с мембраной, способствующую привлечению рецепторов, облегчающих поглощение. Точно так же, когда вытянутая частица находится в положении лежа на клеточной мембране, ее поглощение может быть более эффективным из-за большей площади поверхности контакта с мембраной (39).). Это также может иметь последствия для фагоцитоза, поскольку, например, раковые клетки могут быть менее жесткими и более мягкими, чем нормальные клетки (47), и, возможно, их жесткость является определяющим фактором для фагоцитоза клетками иммунной системы.Наконец, предполагается, что асимметрия мембраны (т. е. различный состав листков мембраны) сильно влияет на фагоцитоз, поскольку это вызывает спонтанное искривление мембраны, которое может способствовать или препятствовать обертыванию мембраны и может формировать резервуар площади мембраны для обеспечения дополнительной мембраны для поглощение частиц (48).
Эти теоретические предсказания имеют несколько оговорок. Во-первых, в реальных биологических системах на взаимодействие между частицами и мембраной может сильно влиять «корона» белков на поверхности частиц, что приводит к несимметричному взаимодействию рецепторов поглощения (49). Кроме того, на клеточное поглощение могут влиять сахара на поверхности частиц (50), так называемый «гликокаликс», состоящий из гликопротеинов, гликолипидов и протеогликанов на поверхности клеток-мишеней (51) и патогенов (52). Во-вторых, плазматическая мембрана иммунных фагоцитов представляет собой сильно переполненную среду с различными рецепторами и многими другими мембранными белками, сгруппированными и сегрегированными в функциональных доменах, и моделирование показало, что мембранное распределение рецепторов может влиять на фагоцитоз (35, 36, 39)., 41). В-третьих, большинство подходов к моделированию исходят из основного предположения, что энергия деформации мембраны является узким местом для поглощения, и не учитывают роль цитоскелета и не включают регуляторную обратную связь.
В то время как было предсказано, что обертывание мембраны вокруг частицы сильно зависит от конкурирующих взаимодействий мембраны и цитоскелета (53, 54), моделирование не учитывает активное ремоделирование цитоскелета, которое может активно способствовать и даже может быть узким местом для , инвагинация частиц (см. ниже). В-четвертых, биологические мембраны содержат большое количество различных компонентов, которые не взаимодействуют с частицей, таких как мембранные белки и (глико)липиды, которые могут влиять на локальную кривизну мембраны и, согласно статистическому механическому моделированию и моделированию молекулярной динамики, сильно влияют на груз. поглощение, даже если общая спонтанная кривизна мембраны остается неизменной (55). В-пятых, количественные сравнения моделируемых и экспериментальных временных шкал, связанных с фагоцитарным процессом, проблематичны и могут быть сделаны только качественно. Из-за этих предостережений экспериментальные данные иногда противоречат теоретическим предсказаниям, как будет показано ниже.Размер и форма модельных частиц: экспериментальные данные
Экспериментальные исследования с синтетическими модельными частицами также показывают, что форма, размер и жесткость являются важными детерминантами фагоцитоза, но, вероятно, из-за высокой сложности клеток экспериментальные данные не всегда совпадают. с предсказаниями теоретического моделирования. Более того, как будет подробно описано ниже, экспериментальные данные между исследованиями не всегда согласуются друг с другом, вероятно, из-за различий в типе клеток, способах поглощения и свойствах фагоцитарных грузов. Тем не менее, из сравнения этих различных исследований можно сделать некоторые общие выводы о том, как размер и форма частиц влияют на эндоцитоз и фагоцитоз модельных частиц.
Модельные частицы могут поглощаться теми же PRR и механизмами поглощения, что и микробы. В кровотоке растворимые антитела и/или белки из системы комплемента могут откладываться на поверхности патогенов, что называется опсонизацией, и это запускает фагоцитоз с помощью Fc-рецепторов и рецептора комплемента 3 (CR3/интегрин α M β 2 /Mac-1) соответственно.
Точно так же опсонизация может опосредовать поглощение модельных частиц. Например, антитела также могут связываться с полианионной поверхностью латексных частиц, что усиливает их поглощение Fc-рецепторами, но связывание антител (в основном IgG1 и IgM) также может запускать классический путь комплемента, приводящий к отложению продуктов расщепления белка С3. распознается рецепторами комплемента клеток-хозяев (56–58). Мишени также могут быть распознаны классом PRR, называемых рецепторами-мусорщиками (SR), например, SCARB1 и MARCO, которые распознают окисленные липопротеины низкой плотности (LDL) на поверхности апоптотических клеток, но также связываются с полианионными структурами, такими как частицы латекса. 56). Наконец, интегрины могут опосредовать фагоцитоз частиц (56, 59).). Таким образом, частицы полистирола без покрытия могут подвергаться фагоцитозу через четырьмя способами: i) прямое связывание с рецепторами-мусорщиками, ii) опсонизация с помощью депонированных в организме хозяина IgG1 или IgM, приводящая к связыванию iC3b с последующим связыванием с рецептором комплемента 3 (57), iii) опсонизация антителами с последующим узнаванием Fc-рецепторами (FcR) (17) и iv) прямое связывание с интегринами (α V β 3 , α 5 β 1 и α V β 5 ) (59). Частицы, состоящие из других материалов, также поглощаются иммунными фагоцитами. Например, частицы кремнезема распознаются рецепторами-мусорщиками SCARA1, SCARB1 и CD36, тогда как кристаллы моноурата натрия (MSU) могут напрямую связываться с холестерином клеточных мембран, способствуя интернализации. Кристаллы MSU и холестерина также могут активировать пути комплемента посредством опсонизации кристаллов фактором комплемента iC3b (60). Пути поглощения частиц не всегда понятны, и, например, механизм поглощения кристаллов гидроксиапатита макрофагами до сих пор не ясен (60).Что касается размера частиц и фагоцитоза, имеющиеся данные позволяют предположить, что существует оптимальный размер сферических частиц диаметром около 3 мкм для наиболее эффективного фагоцитоза. Оба исследования in vivo и in vitro показали, что альвеолярные макрофаги крысы могут эффективно очищать частицы в диапазоне 1–5 мкм (рис. 3) (61–63) с оптимальным размером частиц ~2–3 мкм ( 63). Это согласуется с другим исследованием, посвященным поглощению более мелких (<1 мкм) частиц различной формы, изготовленных из гидрогеля на основе диакрилата полиэтиленгликоля (PEGDA), эндотелиальными клетками, где было обнаружено, что более крупные и более сплюснутые частицы попадают в организм.
более эффективно, чем более мелкие и вытянутые частицы эндотелиальными клетками (26).Существует верхний предел размера частиц, которые клетки могут поглощать, поскольку частицы, превышающие размер макрофагов, могут останавливать фагоцитоз (64), а более крупные частицы приводят к нарушению фагоцитоза (22). Для игольчатых частиц этот предел обычно составляет около 15 мкМ (22, 65), но это зависит от типа клеток: хотя длинные (> 20 мкм) полистироловые червеобразные частицы с соотношением размеров около 22,5 не могут быть полностью интернализованы. альвеолярными макрофагами крысы (27), карбонатом кальция длиной 20 мкм (CaCO 3 ) иглы (соотношение размеров >20) могут интернализоваться макрофагами THP-1 и первичными мышиными перитонеальными макрофагами (32). Что касается сферических частиц, то исследование, сравнивающее частицы поли(молочной-со-гликолевой кислоты) (PLGA) разного размера, которые перорально вводили мышам, показало, что размер частиц 10 мкм был наибольшим размером частиц, которые могли быть фагоцитированы кишечными фагоцитами (66), хотя это открытие также может быть объяснено меньшим проникновением более крупных частиц через слизистый барьер.
Вероятно, клетки не могут фагоцитировать очень большие частицы, потому что для этого требуется больше мембраны, а увеличение площади поверхности мембраны, которая может быть примерно равна площади поверхности мишени, может, таким образом, ограничивать размер частиц, которые могут быть поглощены (67). . Существует предел, до которого плазматическая мембрана может растягиваться без разрыва (68, 69).), около 3% для эритроцитов (68), и для того, чтобы удовлетворить потребность в прочной мембране для покрытия частиц-мишеней, существует два типа источников мембран: i) складки в плазматической мембране и ii) внутриклеточные везикулы и гранулы (70–74). Вполне вероятно, что для фагоцитоза крупных частиц требуется триггер для регуляции мобилизации этих мембранных резервуаров, а сама плазматическая мембрана может обеспечивать механизм механо-химической настройки, создавая натяжение мембраны во время поглощения (75). Действительно, натяжение мембраны может сниматься последовательным образом, когда сначала площадь поверхности плазматической мембраны увеличивается примерно на 20-40% из-за сглаживания складчатой мембраны, а затем дополнительная мембрана образуется за счет экзоцитоза вблизи фагоцитарной чашечки (76). Такая доставка внутриклеточных везикул также играет роль на более поздних стадиях фагоцитоза. Было показано, что внутренний источник мембраны, полученной из лизосом, имеет решающее значение для поддержания целостности мембраны фагосом, содержащих расширяющиеся гифы Candida albicans (74).Хотя поверхностная плотность лигандов для фагоцитарных рецепторов в целом улучшает поглощение, по-видимому, это также зависит от размера частиц. Для фагоцитоза более высокая опсонизация IgG только увеличивала поглощение полистироловых сфер размером от 0,5 до 2 мкм макрофагоподобными клетками RAW264.7, но не для сфер > 2 мкм (18). Когда сравнивали поглощение липосом разного размера альвеолярными крысиными макрофагами и клетками RAW264.7, самые большие протестированные липосомы размером 650 нм – 2 мкм поглощались более эффективно, чем липосомы меньшего размера (77, 78). Для клатрин-опосредованного эндоцитоза поглощение частиц, по-видимому, ограничено размером (~ 100 нм) клатриновой решетки, а более крупные частицы поглощаются менее эффективно (рис.
3). Плоские гексагональные или сферические частицы размером 70 нанометров поглощались с аналогичной эффективностью альвеолярными макрофагами мыши, но уже незначительное увеличение размера до 120 нм приводило к уменьшению поглощения частиц гексагональной формы (79).). Точно так же сферы кремнезема размером 150 нм поглощались клетками RAW264.7 более эффективно по сравнению со сферами размером 250, 500 и 850 нм (80), а круглые наночастицы кремнезема размером 70 нм поглощались более эффективно, чем частицы размером 300 нм и 1 мкм мышиными клетками XS52. эпидермальная клеточная линия Лангерганса (81). Таким образом, в целом фагоцитоз может быть наиболее эффективным для частиц размером около 3 мкм, тогда как эндоцитоз, опосредованный клатрином, оптимален для частиц размером около 100 нм.Теоретические предсказания о том, что сферы поглощаются более эффективно, чем частицы более сложной формы (35, 38, 39) подтверждаются экспериментальными данными (рис. 3). Как для эндоцитоза, так и для фагоцитоза был протестирован широкий спектр частиц, чтобы доказать, что форма, а не размер, больше влияет на клеточное поглощение, и поглощение сфер было более эффективным, чем поглощение любых других вытянутых форм (25, 29).
Точно так же, по сравнению со сферическими формами, удлиненные частицы (полученные из сфер PLGA размером 150 нм или 2 мкм) менее эффективно поглощались макрофагами J774.A1 (82). Клетки HeLa более эффективно эндоцитозировали сферические, а не палочковидные частицы золота нанометрового размера (83). Сферы микрометрового размера, изготовленные из микрокомпозитов с квантовыми точками CdTe и цистеином, фагоцитировались макрофагами RAW264.7 с более высокой эффективностью, чем прямоугольные диски и особенно игольчатые частицы (84). В другом исследовании сообщается, что хотя вытянутые частицы полистирола микрометрового размера лучше всего прикрепляются к клеткам RAW264.7, они менее эффективно фагоцитируются по сравнению со сферами (рис. 3) (30). Это же исследование показало, что не сферы, а сплюснутые дискообразные эллипсовидные частицы интернализуются с наибольшей эффективностью, но эта разница исчезает с увеличением размера частиц (30). По сравнению с другими формами и размерами полистироловые вытянутые эллипсовидные частицы с 2-3 мкм в самом длинном измерении также больше прикреплялись к мышиным J774 и крысиным альвеолярным макрофагоподобным клеткам NR8383, но не давали преимуществ в фактическом поглощении (25, 31). Аналогичное предпочтение прикрепления вытянутых частиц было показано для цилиндров кремнезема размером в нанометры по сравнению со сферами, хотя чрезвычайно длинные частицы (черви) хуже прикреплялись к клеткам RAW264.7 (85). Наконец, вытянутые частицы PLGA неэффективно фагоцитировались мышиными макрофагами J774, но фагоцитировались, как только форма частиц изменялась на более сферическую (диаметр 3 мкм) под давлением и температурой (86).Имеющиеся данные свидетельствуют о том, что для фагоцитоза частиц разной формы требуются разные сигнальные каскады. Эксперименты с амебой Dictyostelium показали, что скоординированная активность малых ГТФаз Rac и Ras в фагоцитарной чашечке под действием многодоменного белка RGBARG (RCC1, RhoGEF, BAR и белок, содержащий RasGAP) является ключевым фактором для захвата частиц и микробов. разной формы (87). Авторы предполагают, что RGBARG способствует выпячиванию цитоскелета на периферии фагоцитарной чашечки за счет усиления активации Rac в этой области, в то время как он подавляет Ras в более центральных областях формирующейся фагоцитарной чашечки (87).
Хотя Dictyostelium без RGBARG показал улучшенный фагоцитоз более крупных модельных частиц и дрожжей, пространственная регуляция Ras с помощью RGBARG оказалась важной для фагоцитоза удлиненных грузов (87).Что касается способа поглощения при фагоцитозе, экспериментальные данные, по-видимому, противоречат большинству теоретических предсказаний (рис. 3). Было показано, что фагоцитоз с первым кончиком клетками NR8383 и J774 более эффективен для уплощенных вытянутых частиц микронного размера (, т. 22.5) (27), показывая, что поглощение стимулируется взаимодействием мембран с областями с высокой положительной кривизной. То же самое наблюдалось при поглощении мышиными альвеолярными макрофагами цилиндров размером 2 × 10 мкм, сделанных из нескольких сфер кремнезема, склеенных вместе с агарозой (33). Интересно, что было обнаружено, что мембрана фагоцитарной чашечки перемещается по длине ортогонально расположенных цилиндров, показывая, что фагоциты активно искали кончики цилиндров с высокой кривизной перед поглощением (33).
Было обнаружено, что сплюснутые частицы полистирола размером в микрометры неэффективно фагоцитируются в режиме «наконечник-сначала» клеточными линиями макрофагов крысы и мыши (25). Аналогичное предпочтение для поглощения via высокая кривизна (кончиком вперед для вытянутых частиц или укладыванием вниз для сплюснутых частиц) по сравнению с малой кривизной (укладка вниз для вытянутых и кончиком вперед для сплюснутых частиц) контакт с мембраной наблюдался для прямоугольных дисков, состоящих из квантовой точки CdTe-цистина. микрокомпозиты клетками RAW264.7 (84). Таким образом, все эти данные показывают, что фагоцитоз более эффективен при контакте с более высокими изогнутыми областями мембраны частиц, и это, по-видимому, противоречит большинству теоретических предсказаний (рис. 3) (35, 38).Теоретические предсказания о том, что более жесткие частицы, как правило, лучше усваиваются клетками, также не всегда верны (39). Обзор доступных экспериментальных данных по эластичности частиц (39) показал, что иногда менее жесткие/более мягкие частицы интернализуются с большей эффективностью, чем более жесткие частицы.
Например, было обнаружено, что фагоцитоз более мягких послойных (LBL) капсул клетками HeLa и SUM159 более эффективен, чем более жестких аналогов (39). Однако сообщалось и об обратном, например, опсонизированные антителами деформируемые поли-AAm-co-AAc микрочастицы (DAAM-частицы) с относительно высокой жесткостью 7 кПа были фагоцитированы мышиными макрофагоподобными клетками J774 с в 6 раз более высокой эффективность, чем у частиц 1 кПа (19). Это противоречие, вероятно, связано с различными способами поглощения для разных частиц и типов клеток. Например, клатриновые оболочки относительно жесткие и имеют четко определенную форму, а в некоторых случаях менее жесткие частицы могут быть легче проглатываемы, поскольку они могут быть деформированы для лучшего размещения в покрытых клатрином везикулах.В частности, частицы кремнезема могут быть токсичными для клеток, хотя, похоже, нет единого мнения о зависимости этой токсичности от размера. В исследовании, сравнивающем поглощение кварцевых сфер размером 150–850 нм клетками RAW264.
7, более крупные частицы вызывали более высокую токсичность, о чем свидетельствует более высокий разрыв мембраны, АФК и продукция фактора некроза опухоли (ФНО)-α (80). Однако это контрастирует с данными, полученными в эпидермальной клеточной линии Лангерганса мышей XS52, где наночастицы аморфного кремнезема размером 70 нм были более цитотоксичными по сравнению с частицами размером 300 нм и 1 мкм, что оценивалось по высвобождению ацетатдегидрогеназы (ЛДГ) (81). Более того, кристаллические частицы кремнезема размером 0,3 мкм вызывали большую продукцию АФК и ФНО-α в клетках RAW264.7, чем более крупные частицы размером 4,1 мкм (88). Также было доказано, что в нефагоцитирующих клетках HeLa сферические частицы кремнезема размером 70 нм более цитотоксичны, чем частицы размером 200 и 500 нм, и это было связано с их более высокой тенденцией к лизосомальной локализации (89).). Токсичность кремнезема, вероятно, вызвана активацией воспаления, как будет обсуждаться ниже.Размер и форма патогена: экспериментальные данные
Выводы, полученные на модельных частицах, не всегда могут быть непосредственно применимы к реальным патогенам, поскольку патогены могут задействовать разные фагоцитарные рецепторы, что приводит к различным способам поглощения.
Более того, иногда патогены могут реконструировать свою форму и экспрессировать факторы вирулентности, которые могут влиять на процесс фагоцитоза (90–93). В этом разделе мы предоставляем обзор того, как форма и размер патогенов влияют на фагоцитоз. Фагоцитоз патогенов разной формы и размера лучше всего понятен для грибов, которым, таким образом, уделяется основное внимание в этом разделе.Различные штаммы патогенных грибов C. albicans могут иметь различную морфологию, от сферических дрожжей до удлиненных клеток гиф (рис. 4А) (7, 8, 13, 93–97). Как было предсказано путем моделирования сферических частиц по сравнению с удлиненными частицами (см. выше), живые C. albicans со сферической морфологией поглощались более эффективно, чем гифы C. albicans и гифы размером более 20 мкм (но не менее 20 мкм). значительно препятствовал поглощению макрофагами J774.1 (7). Однако фагоцитоз живых мутантных штаммов дрожжей C. albicans , который не может образовывать гифы (и, следовательно, имеет более сферическую форму), был значительно медленнее, чем штаммы, которые образовывали гифы (7).
Эффективность фагоцитоза также зависела от ориентации патогена, потому что клетки гиф C. albicans , которые были расположены по направлению к фагоциту в ориентации кончик-сначала, поглощались быстрее, чем те, которые были поглощены под углом или где клетка-клетка. контакт находился в положении лежа (рис. 4А) (7). Также для бактерий фагоцитоз нитчатых 9Бактерии 0649 Escherichia coli макрофагами J774A.1 требуют доступа к одному из концевых полюсов бактериальных филаментов (кончиком вперед), в то время как метод захвата лежащим вниз (с самой длинной осью E. coli , ориентированной параллельно макрофагу). поверхности) не удалось, и не наблюдалось накопления актина и образования фагоцитарной чашечки (98). Как обсуждалось выше, это противоречит предыдущим теоретическим предсказаниям поглощения вытянутых эллипсоидов, но согласуется с экспериментальными данными с модельными частицами (рис. 3).Рисунок 4 Морфология грибов влияет на фагоцитоз. (A) Эффективность поглощения дрожжевых и гифальных форм Candida albicans и Exserohilum rostratum макрофагами.
(B) Paracoccidioides brasiliensis может образовывать звездообразные формы мать-дочь. Cryptococcus gattii образует крупные титановые клетки, а Coccidioides spp. образует шарики, размер которых превышает размеры фагоцитирующих клеток. Подробности; см. текст.В отличие от удлиненных стержней или филаментных форм бактерий, которые захватываются предпочтительно кончиком вперед, фагоцитоз сильно изогнутых спиралевидных видов бактерий, например Campylobacter и Helicobacter , может быть остановлен из-за их форма (27, 99). Моделирование продемонстрировало ингибирование поглощения таких вытянутых частиц в горизонтальной ориентации из-за высоких энергетических затрат, необходимых для изгиба плазматической мембраны вокруг двух разных изгибов (9).9). Таким образом, сочетание положительной и отрицательной кривизны в спиралевидных частицах может препятствовать обертыванию мембраны вокруг скрученной поверхности и тем самым предотвращать фагоцитоз.
Гифы C. albicans поглощаются быстрее, если фагоцитоз живых грибов инициируется на дрожжевом конце, чем на конце гиф (7), возможно, потому, что круглая морфология дрожжевого конца более допустима для усвоения. Аналогичное наблюдение было сделано для поглощения как живых, так и убитых нагреванием веретенообразных спор гриба 9.0649 Exserohilum rostratum (~20 × 60 мкм) (9). Они слишком велики для поглощения, но ~84% макрофагов пытались поглощать на полюсах (напоминает поглощение кончиком вперед) и только ~16% располагались в средней части споры (режим поглощения укладыванием) (9). Однако это дифференциальное расположение макрофагов может быть связано не с формой спор, а скорее с различиями в составе клеточной стенки на полюсах и в середине микробов (9).
Некоторые Штаммы C. albicans могут изменять свою форму после фагоцитоза, потому что C. albicans может избежать своего разрушения и уклониться от иммунного ответа, образуя зародышевые трубки и дефис внутри запечатанной фагосомы, что приводит к перфорации фагосомной мембраны (рис.
4А) (95). , 100). Образованию гиф способствуют условия внутри фагосом, такие как оксид азота (101–103), H 2 O 2 (104, 105) и щелочной рН (106). Candida albicans активно подщелачивает просвет фагосом, используя аминокислоты в качестве источника углерода и выделяя аммиак (94).Грибок Blastomyces dermatitidis может использовать метод «троянского коня», при котором он сначала эффективно фагоцитируется в форме мелких спор (2–5 мкм) альвеолярными макрофагами, а затем выживает внутриклеточно, увеличивая свой размер в 10 раз и распространяясь из клетка-хозяин в виде более крупной дрожжевой формы (10–30 мкм) (107). Эта крупная форма дрожжей также может поглощаться фагоцитами, но менее эффективно по сравнению с более мелкой споровой формой (107). Не только грибы, но и некоторые бактерии могут принимать нитевидную морфологию, чтобы избежать фагоцитоза (108-110), но как это влияет на созревание фагосом, не до конца понятно.
Как уже упоминалось выше, споры (~ 20 × 60 мкм) или гифы E.
rostratum слишком велики для фагоцитоза макрофагами RAW264.7 и первичными макрофагами костного мозга мыши (рис. 4A) (9). Точно так же грибковый патоген Cryptococcus gattii способен образовывать титановые криптококковые клетки диаметром 50–100 мкм, что больше, чем у большинства иммунных фагоцитов, и препятствует фагоцитозу (13, 111), тогда как клетки меньшего размера (5–10 мкм) Криптококк неоформанс эффективно фагоцитируются (112, 113). Другие грибы, такие как Coccidioides immitis и Coccidioides posadasii , также образуют гигантские шарики размером около 120 мкм, чтобы избежать поглощения фагоцитами хозяина. Попадая в организм хозяина, патогенные грибы Paracoccidioides brasiliensis и Paracoccidioides lutzii могут препятствовать фагоцитозу, образуя так называемые астероидные тельца, представляющие собой круглые колонии с радиальной симметрией материнских клеток (~30 мкм 9 ).1739 2 ), окруженных дочерними клетками, достигающими площади от 75 мкм 2 до более 150 мкм 2 в диаметре (рис. 4B) (114). Нокдаун регулятора актина Cdc42p у P. brasiliensis вызвал уменьшение размера этих астероидных тел и способствовал поглощению макрофагами костного мозга мышей и более эффективному выведению из кровотока после внутривенной инъекции мышам, демонстрируя, что этот патоген увеличивает свою активность. размера, чтобы предотвратить клиренс иммунной системы хозяина (115).Данные свидетельствуют о том, что нейтрофилы могут ощущать размер микробов. Фагоцитоз живых мелких мутантных штаммов C. albicans , запертых в дрожжах, запускает продукцию АФК и рекрутирование эластазы в фаголизосомы, тогда как поглощение более крупных форм гиф C. albicans или больших агрегатов Mycobacterium bovis приводит к транслокации эластазы в ядра нейтрофилов (6). Последняя ядерная транслокация эластазы вызывает протеолитическое расщепление гистонов и деконденсацию хроматина, что приводит к высвобождению NET, нацеленных на патогены (рис. 1А) (6). Наконец, почкующиеся дрожжи могут остановить фагоцитоз, когда фагоцитарная чашечка достигает отрицательной кривизны на шейке, разделяющей материнскую и дочернюю клетки, как это показано для фагоцитоза живых дрожжей амебой 9.
0649 Dictyostelium (116) и макрофагами RAW264.7 для фиксированных параформальдегидом и убитых нагреванием дрожжей (117). Это согласуется с выводами теоретического моделирования о том, что частицы в форме песочных часов могут останавливать фагоцитоз (рис. 5А) (34).Рисунок 5 Патоген и форма частиц влияют на фагосомальный F-актин. (A) Теоретическое моделирование опосредованного рецептором поглощения частиц песочных часов и экспериментальные данные почкующихся дрожжей. Локальная отрицательная кривизна на шее может замедлить поглощение. Кольца F-актина присутствуют во время поглощения дрожжами (117, 118) и вытянутыми частицами (64). (B) F-актин в фагосомах, содержащих грибы, которые изменяют морфологию внутри клеток-хозяев. F-актин образует волнообразные динамические кольца и манжеты вокруг гиф во время интернализации. На более поздних стадиях гифы покрывают трубки динамического F-актина (93, 97, 119). Кончики гиф, выступающие из мембран, также рекрутируют F-актин.
Подробности; см. текст.Фагоцитоз и цитоскелет
Фагоцитоз зависит от актинового цитоскелета, который способствует выпячиванию мембраны для поглощения, формируя чашеобразную складку мембраны вокруг частицы (3, 17, 21) (рис. 1 и 4В). В основании фагоцитарной чашечки F-актин очищается, тем самым обеспечивая инвагинацию мембраны в процессе ремоделирования фосфоинозитидами (1, 2). По краю фагоцитарной чашечки богатые F-актином псевдоподии продолжают распространяться вокруг частицы до тех пор, пока они полностью не охватывают частицу и не сливаются, тем самым запечатывая частицу в фагосому (1, 2). Цитоскелет F-actin воздействует на частицы, подтягивая их к центру клетки, и регулирует события слияния мембран (118, 120-122). Более того, сократительная активность F-actin необходима для закрытия фагоцитарной чашечки (17, 21). Роль F-actin в фагоцитарном процессе широко рассматривалась в других работах (3, 17, 123-127), и мы ограничим наше обсуждение ролью и организацией F-actin для поглощения фагоцитарных грузов различной формы и размера.
Организация F-актина на поверхности фагосом вполне может зависеть от формы проглоченной частицы. Микроскопия со сверхвысоким разрешением показала, что F-актин выстраивается в концентрические кольца или параллельные волокна на поверхности фагосом, содержащих (сферические) зимозановые дрожжевые частицы (рис. 5А) (118). До сих пор лишь в ограниченном числе исследований рассматривался вопрос о том, как F-актиновый скелет организован на субдифракционном уровне на фагосомах, содержащих частицы/патогены неправильной формы (128), но поскольку многие белки и липиды участвуют в закреплении цитоскелета (интегрины, фосфоинозитиды, белки ERM) чувствительны к искривлению, можно ожидать, что фагосомы, несущие частицы неправильной формы, будут иметь отчетливые участки прикрепления к цитоскелету (129)., 130). Более того, из-за персистентной длины F-актиновых филаментов можно ожидать, что F-актиновые филаменты, связанные с фагосомами, содержащими удлиненные частицы, будут выстраиваться вдоль длинной оси таких частиц.
В соответствии с этим экспериментов in vitro с полимеризацией актина, инкапсулированного в искусственных липосомах, преобразовали морфологию липосом из сферической в трубчатую (131). Представление о том, что образование F-актина зависит от формы частиц, подтверждается открытием, что F-актин формирует кольцо только на переднем крае фагоцитарной чаши в альвеолярных макрофагах, фагоцитирующих сферы или уплощенные вытянутые частицы (~ 14 × 4 × 1 мкм). ) при условии, что они были фагоцитированы ортогонально их главной оси (кончиком вперед) (рис. 5А). Напротив, даже несмотря на то, что F-актин собирался, если первоначальный контакт с частицами происходил ортогонально их малой оси (укладка), он не мог образовывать чашеобразные расширения мембраны или кольцо F-актина, что останавливало поглощение (64). ). Точно так же поглощение нитевидных Бактерии E. coli макрофагами J774A.1 оказались безуспешными, и не наблюдалось накопления актина и образования фагоцитарной чашечки, когда бактерии располагались своей длинной осью параллельно поверхности макрофага (укладка вниз) (98).Исследование с использованием DAAM-частиц с жесткостью, сравнимой с биологическими объектами, показало, что во время фагоцитарного поглощения мышиными макрофагоподобными клетками J774A.1 псевдоподы фагоцитов оказывают кольцевидное сжатие на частицы DAAM (19). Интересно, что эти кольца не были круглыми и содержали вмятины и ямки, указывающие на неравномерность давления по периметру кольца (19). Вмятины могут быть вызваны локализованным давлением подосомоподобных пятен F-актина на фагоцитарных чашечках (132). Во время поглощения DAAM-частиц в основании фагоцитарной чашечки также наблюдались ямки, вызванные силами сжатия, и эти ямки наблюдались даже во время закрытия фагоцитарной чашечки (19). Кажется маловероятным, что эти сжимающие силы вызываются F-актиновым цитоскелетом, т.к. F-актин очищается от основания чашечки на этой стадии фагоцитоза (133).
F-актиновый цитоскелет не только опосредует фагоцитоз, но и играет роль позже, во время созревания фагосомы, и грибковые патогены иногда экспрессируют факторы вирулентности, препятствующие этому.
Например, внутриклеточный патоген Cryptococcus neoformans может блокировать закисление своих фагосом в процессе, управляемом патогеном, и нарушать целостность фагосомной мембраны, чтобы получить доступ к цитозолю (134). За этой пермеабилизацией фагосом следует временное появление F-актина на поверхности фагосомы во время слияния фагосомы с плазматической мембраной, что приводит к нелитическому высвобождению C. neoformans из клетки (вомоцитоз) (135). В дополнение к факторам вирулентности размер, форма и/или свойства поверхности C. neoformans также могут способствовать этим актиновым вспышкам, поскольку они наблюдались не только в фагосомах с живым патогеном, но также и с инактивированным нагреванием патогеном и они были более стойкими, чем латексные шарики (135). Вопрос в том, выполняет ли этот F-актин защитную функцию против разрыва фагосомальной мембраны. Такие вспышки F-актина также наблюдались на фагоцитированных латексных гранулах и термоинактивированных C. neoformans , но эти вспышки происходили с меньшей частотой, и такие частицы не были способны к нелитическому изгнанию (135). Более того, удаление F-актина из фагосомы может быть необходимым шагом перед тем, как фагосома сможет слиться с эндосомами и лизосомами (117, 135), хотя это спорно, поскольку было показано, что связывающий F-актин белок эзрин стабилизирует фагосомальный F-актин, стимулирующий рекрутирование лизосомального мембранного белка LAMP2 (136).Имеются указания на то, что рекрутирование F-актина на более поздних стадиях созревания фагосом зависит от формы проглоченной частицы. Шейка почкующихся дрожжей создает отрицательную кривизну между материнской и дочерней клетками, которая останавливает F-актин в этой области во время фагоцитарного захвата макрофагами RAW264.7, что приводит к постоянным вспышкам F-актина в этой области (рис. 5B) (117). F-актин присутствовал только на ранних стадиях фагоцитарного захвата 9.0649 мутантов C. albicans , которые не могли образовывать гифы, тогда как он также присутствовал после фагоцитоза образующих гифы дикого типа C.
albicans . Фактически, на фагосомах, содержащих гифальную форму C. albicans , наблюдались различные топологии F-актина: манжеты F-актина в точках входа (рукавоподобные расширенные фагоцитарные чашечки, также наблюдаемые в (97)), трубочки F-актина, выстилающего полностью интернализованные гифы (в отличие от дрожжевой формы, не имеющей F-актина), и F-актина на кончиках гиф (93) (рис. 5Б). Однако спорно, насколько эти различия в локализации F-актина обусловлены геометрией патогена и насколько это зависит от состава клеточной стенки; F-actin в меньшей степени рекрутировался в фагосомы, содержащие мутантов C. albicans с дисфункциональным O-маннозилированием (и разрушенными клеточными стенками), хотя этот мутант образовывал гифы, сравнимые с грибами дикого типа (93).Кроме того, C. albicans могут экспрессировать факторы вирулентности, которые изменяют образование F-актина в фагосомах. Использование ингибитора PI3K класса I LY2
, было показано, что активность PI3K не нужна для более поздней трубчатой полимеризации F-актина вокруг гиф C.
albicans (97). Поскольку известно, что PI3K класса I играет существенную роль в кортикальной динамике F-актина (2, 4, 137), это предполагает, что C. albicans может запускать сигнальные каскады, чтобы уклоняться от защитных реакций хозяина, либо создавая F-актин барьер, независимый от киназ класса I PI3K, который предотвращает слияние с лизосомами, или за счет использования F-актина для восстановления разрыва мембраны, вызванного быстро растущими гифами. Интересно, что роль PI3K в эндоцитозе (138), по-видимому, зависит от формы частиц, поскольку широкий ингибитор PI3K вортманнин может блокировать поглощение кремнеземных червей размером в нанометры RAW264.7 и альвеолярными макрофагами, но влияет на поглощение сфер в меньшей степени. (85).Хотя изучено гораздо меньше, не только F-актиновый цитоскелет, но и микротрубочки могут влиять на фагоцитоз. Во-первых, микротрубочки контролируют распространение и миграцию макрофагов RAW264.7, тем самым связывая это с фагоцитарной способностью (139).
Участие микротрубочек зависит от фагоцитарного груза, так как фагоцитоз сферических неопсонизированных и опсонизированных комплементом частиц кремнезема размером 3 мкм клеточной линией альвеолярных макрофагов мыши MH-S, но не частиц кремнезема, опсонизированных антителами, зависит от микротрубочек. цитоскелет (140). Во-вторых, после фагоцитоза проглоченные частицы перемещаются от клеточной периферии к центру организации микротрубочек (MTOC), расположенному в центре клетки (139)., 141). Этот ретроградный внутриклеточный транспорт, вероятно, обеспечивается моторными белками семейства динеинов (142). Таким образом, микротрубочки ответственны за внутриклеточный транспорт фагосом (143, 144) и это облегчает слияние фагосом с эндоцитарными органеллами (145). При опосредованном Fc-рецептором фагоцитозе макрофагами RAW264.7 микротрубочки также переориентируют MTOC в направлении фагосомного груза, и это необходимо для позиционирования аппарата Гольджи рядом с фагосомой для последующей презентации антигена Т-клеткам (141).Актин и микротрубочки также важны для клатрин-опосредованного эндоцитоза, и это зависит от формы частиц. Например, было показано, что низкомолекулярные ингибиторы F-актина или ингибиторы сборки микротрубочек в макрофагах вызывают пониженное поглощение наночастиц более сложной формы, таких как черви и цилиндры, но не более чувствительных сферических частиц. к ингибиторам клатрина (85).
Поскольку ожидается, что форма частиц будет определять силы сопротивления и сопротивления, с которыми фагосома сталкивается внутри клетки, логично предположить, что частицы будут переориентироваться внутри клетки, чтобы свести к минимуму силы, необходимые для внутриклеточного транспорта. В соответствии с этим сообщалось, что начальный контакт не является единственным параметром, определяющим направление фагоцитоза, и (по крайней мере, сферические) частицы могут вращаться после захвата (146).
Фагоцитоз и транспортировка органелл
Большинство исследований клеточного поглощения частиц различной формы и размера сосредоточено только на механизме поглощения, а не на созревании фагоцитарных/эндоцитарных органелл в клетках-хозяевах.
После полного поглощения частиц плазматической мембраной как эндосомы, так и фагосомы подвергаются процессу, называемому эндо/фагосомным созреванием. Это созревание управляется событиями слияния мембран, где компартменты ранней эндосомной и лизосомальной природы сливаются с эндо/фагосомами, и событиями деления, характеризующимися отпочкованием везикул от эндо/фагосом (1, 147-150). Цитозольные компоненты, такие как Rab-белки, их эффекторы, SNARE, SNARE-взаимодействующие белки, BAR-белки и фосфоинозитидкиназы, ремоделируют мембрану эндо/фагосом и тем самым также управляют созреванием (1, 147, 148). После поглощения микробного патогена фагоцит пытается убить проглоченный патоген, генерируя большое количество активных форм кислорода (АФК) с помощью НАДФН-оксидазы NOX2 в просвете фагосомы (151). Во время эндо/фагосомного созревания эндо/фагосома обогащается липазами, нуклеазами и протеазами (126, 152). Среди протеаз есть катепсины, которые играют роль в процессинге антигена и препроцессинге цепи шаперона МНС класса II для презентации антигена Т-клеткам (152). Образовавшаяся в результате лизосома (эндоцитоз) или фаголизосома (фагоцитоз) представляет собой компартмент деградации, характеризующийся низким рН в результате импорта протонов вакуолярной (v-)АТФазой из цитоплазмы в просвет фагосомы, что позволяет расщеплять проглоченные вещества. возбудитель (1, 94, 108, 137).Представление о том, что перемещение мембран приводит к асимметрии в созревании фагосом, хорошо известно. Эта асимметрия может возникать уже на ранней стадии фагоцитарного процесса в зарождающейся фагоцитарной чашечке. Например, было замечено, что слияние мембран во время фагоцитоза крупных сферических частиц приводило к смещению первоначальных компонентов мембраны в основании фагосомальной чашечки, которая, таким образом, становилась более зрелой, чем периферические области, и эта гетерогенность была более очевидной для более крупных частиц. 153). В соответствии с этим, четкий пространственный градиент фосфоинозитидных липидов и белков p85 и SHIP1 наблюдался на ранних стадиях фагоцитоза вытянутых полистирольных частиц эллипсоидной формы размером ~10 мкм (154).
В макрофагоподобных клетках RAW264.7 поглощение PFA-инактивированных филаментозных Legionella pneumophila , Salmonella typherium и C. albicans развивались через долгоживущих фагоцитарных чашечек и эндосом, уже слившихся с этими зарождающимися фагосомами и высвободивших свое содержимое (23, 155). Однако, несмотря на приобретение поздних фагосомальных маркеров (например, Rab7, LAMP1), просвет формирующихся фагосом не закисляется до запечатывания, и низкомолекулярные соединения и лизосомальные гидролазы могут просачиваться во внеклеточную среду (23, 155). Это показывает, что морфология частиц влияет на ремоделирование и закрытие фагоцитарной чаши, закисление фагосом и целостность фагосомной мембраны.Также после запечатывания фагоцитарной чашечки можно ожидать, что асимметрия созревания присутствует на фагосомах, несущих сильно изогнутые частицы, поскольку многие фагосомные белки (включая белки домена BAR, оболочки везикул) и липиды (фосфоинозитиды, фосфатидные кислоты) реагирует на искривление мембраны (129, 130, 156, 157).
Поскольку эти белки и липиды регулируют слияние и деление внутриклеточных мембран, это повышает вероятность того, что события органелларного переноса, управляющие созреванием фагосом, могут преимущественно происходить в сильно изогнутых участках мембраны фагосом, содержащих частицы неправильной формы. Это может способствовать патогенности C. albicans , так как было обнаружено, что фагосомы, содержащие гифальную форму патогена, созревают медленнее (93) и имеют длительное присутствие малой ГТФазы Rab14, которая способствует рекрутированию LAMP1 и катепсина B, ставя под угрозу выживаемость грибов и ускользание от иммунитета (119) . В этом исследовании было обнаружено, что пролонгированное удержание Rab14 на фагосомах пропорционально длине гиф, что подтверждает гипотезу о том, что форма частиц является ключевым параметром для созревания фагосом.Активация воспаления
Некоторые MAMP и DAMP могут восприниматься важным классом PRR: нуклеотидсвязывающим доменом и белками, богатыми лейцином, повторами (NLRP) (158).
В цитозоле NLRP могут собираться в большой макромолекулярный комплекс, называемый инфламмасомой, который запускает активность каспазы-1. Каспаза-1 вызывает выработку воспалительных цитокинов IL-1β и IL-18, что приводит к воспалению и гибели воспалительных клеток (пироптозу) (рис. 6А). При различных патологических состояниях могут быть обнаружены кристаллы квасцов (например, в вакцинных адъювантах), уратов (при заболевании подагрой), холестерина (атеросклеротические поражения), пирофосфата кальция (псевдоподагра), асбеста (асбестоз, мезотелиома) и кремнезема (силикоз). поглощаются фагоцитами и вызывают активацию воспаления (158–160), и это является основным объяснением цитотоксичности частиц кристаллического кремнезема (см. выше). Активация инфламмасом может быть вызвана дестабилизацией мембран лизосом, приводящей к высвобождению катепсина В в цитозоль и/или оттоку ионов калия через К + каналов (рис. 6A, B), которые способствуют активации воспаления и продукции IL-1β и IL-18, но также могут усиливать апоптоз из-за высвобождения цитохрома C из митохондрий (158, 161–164).Рисунок 6 Размер и морфология частиц: активация воспаления. (A) Частицы с остроконечной морфологией могут разрушать эндо/фагосомные мембраны, что приводит к K + и утечке катепсина в цитозоль (так называемый сигнал 2). Митохондриальная дестабилизация вызывает высвобождение цитохрома С, что приводит к цитотоксичности. Эти стимулы активируют сборку инфламмасомы из каспазы-1, ASC и NLRP3, которая расщепляет про-ИЛ-1β и про-ИЛ-18 на зрелые ИЛ-1β и ИЛ-18. Активация воспалительного процесса может вызвать гибель клеток (пироптоз). (B) Форма частиц является агонистом в вакцинации на основе частиц. Введение шиповидных частиц TiO 2 , инициирующих механический разрыв фагосом (сигнал 2) у мышей вместе с бактериальным лигандом TLR (сигнал 1). Передача сигналов TLR активирует перемещение NF-κB в ядро, что приводит к транскрипции и трансляции про-IL-1β и про-IL-18, а также к повышению активности маркеров активации CD40, CD80 и CD86.
Сигнал 2 инициирует опосредованное инфламмасомами протеолитическое превращение в зрелые IL-1β и IL-18. Таким образом, мыши, получающие комбинацию шиповидных частиц и лиганда TLR, индуцируют сильный ответ Th2.Активация инфламмасомы частицами зависит от свойств их поверхности. Частицы с реактивным химическим составом поверхности и/или частицы с шероховатой топологией поверхности (например, диоксид кремния, сажа, серебро, полистирол) могут дестабилизировать лизосомальные и фагосомальные мембраны, тем самым вызывая активацию воспалительных процессов, как было подробно рассмотрено в других работах (158, 165–168). . В целом, для таких частиц с шероховатой и/или реакционноспособной поверхностью наночастицы меньшего размера более эффективны для активации воспаления, поскольку они имеют большую площадь поверхности на единицу массы, что также рассматривалось в другом месте (рис. 7) (166, 169).). Однако сообщалось об обратной зависимости от размера длинных частиц с экстремальным AR. Такие игольчатые частицы также могут приводить к активации воспаления, даже если они состоят из биологически инертного материала, и это является отличительной чертой парадигмы волокон, объясняющей токсичность материалов, подобных длинным волокнам (асбест, углеродные наночастицы) (22).
Для таких частиц с экстремальной числовой апертурой более крупные частицы, как правило, в большей степени способны активировать инфламмасомы, что хорошо известно для игольчатых частиц рутила титана, полиэтиленоксида, золота, углерода и других материалов (рис. 7) (161, 166). Активация инфламмасомы игольчатыми частицами зависит от продукции АФК и катепсина В (170), и хотя точный механизм и вклад дестабилизации мембраны в этот процесс менее ясны (170, 171), это, вероятно, связано с нарушением фагоцитоза. и утечка АФК и литических ферментов во внеклеточную среду (парадигма волокон) (22). Подобно игольчатым частицам, частицы неправильной формы с высокой кривизной также могут индуцировать инфламмасомы, и здесь более крупные частицы также более эффективны, чем более мелкие (161, 164, 166). Например, частицы CoCrMo неправильной формы размером 6 мкм были более эффективны для продукции IL-1β первичными моноцитами человека и моноцитами человека THP-1, чем гладкие частицы аналогичного размера и частицы неправильной формы размером 1 мкм (163).Рисунок 7 Краткая информация о геометрии частиц при активации воспаления (выработка IL-1β) и цитотоксичности. Влияние размера частиц на активацию воспаления и цитотоксичность является спорным. Подробности; см. текст.
Инфламмасомы также могут быть вызваны поглощением определенных патогенов. Здесь инфламмасома может быть активирована после дестабилизации эндосомальных или фагосомальных мембран факторами вирулентности, но также некоторые топологические сигналы, такие как гифы грибов или некоторые бактериальные наноструктуры, такие как жгутики, пили и фимбрии, могут активировать инфламмасому (74, 167, 168). Для возбудителя C. albicans , это играет роль в его уходе от фагоцитов путем литического изгнания после пироптоза (воспалительно-опосредованной гибели клеток) (96). Утверждается, что гифы C. albicans могут запускать активацию инфламмасомы, которая, в свою очередь, запускает цитотоксичность фагоцитов (94), и что таким образом патоген может ускользнуть от фагоцитов.
Вопрос о том, является ли филаментация единственным триггером такого пироптоза, обсуждается, но все согласны с тем, что только разрыв фагосомы вызывает не гибель клеток, а скорее комбинацию активации NLRP3, физической деструкции клеток, изменений в морфологии патогенов (9).6) и ремоделирование клеточной стенки грибов (172).Размер частиц и морфология в иммунной системе
PRR могут не только опосредовать поглощение, но также передавать внутриклеточные сигналы, а IL-1β и IL-18 имеют решающее значение для иммунной системы; поэтому неудивительно, что размер и форма частиц могут модулировать иммунную активацию, как показано как на культивируемых иммунных клетках, так и на животных моделях (рис. 8). Например, по сравнению со сферическими частицами, состоящими из нуклеиновых кислот и катионных пептидов, эндоцитоз культивируемыми спленоцитами мыши волокнистых частиц размером 10–100 нм из того же материала приводил к большей экспрессии маркера активации CD86 и воспалительного цитокина IL-6, тогда как экспрессия костимулирующего рецептора CD40 и продукция воспалительного цитокина IFN-γ были снижены (173).
В другом исследовании фагоцитоз игольчатых частиц карбоната кальция размером 15–20 мкм макрофагальной клеточной линией THP-1 приводил к большей секреции TNF-α и IL-8, тем самым вызывая провоспалительную реакцию (32). Наконец, у мышей, которым вводили частицы размером 7–8 мкм со сложной геометрией, рекрутирование нейтрофилов к месту инъекции было более быстрым, чем при использовании гладких частиц, а фагоцитоз, активация инфламмасомы и секреция IL-1β увеличивались (174). Сравнение этих 9Таким образом, системы 0649 in vitro и in vivo последовательно показывают, что как эндоцитарные, так и фагоцитарные грузы с высокой числовой апертурой вызывают более сильные иммунные ответы, чем более сферические частицы.Рисунок 8 Размер и морфология частиц: иммунный ответ. (A) Резюме иммуномодулирующих эффектов физических свойств частиц. Аг, антиген. (B) Краткое изложение влияния физических свойств частиц на CD4 + Дифференцировка Т-клеток (Th2 и Th3) и поляризация макрофагов (M1 и M2).
Подробности; см. текст.Размер частиц также является важным фактором, определяющим иммунную активацию. Кристаллы гидроксиапатита размером менее 1–2 мкм вызывали большее высвобождение TNF-α культивируемыми первичными макрофагами человека по сравнению с частицами размером 6 и 14 мкм (175), вероятно, из-за более эффективной активации инфламмасомы, как обсуждалось выше (166). Однако размер частиц также влияет на процессинг антигенов для активации Т-клеток. Антигенные белки, конъюгированные с частицами PLGA нанометрового размера, более эффективно вызывали перекрестную презентацию MHC-I по сравнению с частицами микрометрового размера в культивируемых DC (рис. 8A) (рассмотрено в (176)). Здесь было обнаружено, что частицы разного размера по-разному процессируются антигенпрезентирующими ДК: белки, связанные с частицами нанометрового диапазона, подвергались процессингу протеасомой для кросс-презентации в культивируемых ДК, что указывает на то, что они ускользают из просвета эндосомы в цитозоль (177).
Напротив, белки, связанные с частицами микрометрового размера, подвергались процессингу эндолизосомными протеазами (177), что указывает на то, что эти белки не проникали в цитозоль.Размер эндоцитарных частиц также влияет на иммунный ответ, определяя скорость созревания эндосом. Эндосомы, содержащие более мелкие частицы серебра размером 200 нм, приобретают лизосомальные маркеры, такие как LAMP1, раньше, чем эндосомы, несущие более крупные частицы размером 500 нм, в культивируемых мышиных макрофагах костного мозга (178). Хотя MHC-II рекрутировался в эндосомы, содержащие оба типа частиц, презентация антигена происходила более эффективно с более крупными частицами, и это способствовало более эффективному CD4 9.Активация Т-клеток 1739 + (рис. 8А) (178). В этом случае эффективное привлечение лизосом к эндосомам, содержащим более мелкие частицы, может привести к полному разрушению антигенного белка и тем самым снизить эффективность презентации MHC-II (179). Действительно, другие исследования показали, что не только поглощение, но и внутриклеточный процессинг определяется размером частиц: частицы размером 20–40 нм, состоящие из амфифильной поли(гамма-глутаминовой кислоты), не только более эффективно перевариваются культивируемым мышиным костным мозгом.
–производные ДК и макрофагоподобные клетки RAW264.7, чем более крупные частицы размером 200–1000 нм (179, 180), но также подвергались более быстрой лизосомной деградации и с меньшей вероятностью располагались в более ранних эндосомальных компартментах (178, 179).Поскольку форма и размер частиц могут влиять на иммунную систему, они могут влиять на эффективность вакцин на основе частиц (обзор (181)). Во-первых, размер частицы является важным параметром для определения того, где частица оказывается в организме. Крупные частицы (>1 мкм) обычно накапливаются в мелких капиллярах, особенно в легких, и могут вызывать тромбоз (181). Частично из-за поглощения макрофагами и моноцитами, которые имеют оптимальный размер частиц для эндоцитоза около 100 нм, как обсуждалось выше, частицы с этим диапазоном размеров обычно легче выводятся из кровотока. Например, в одном исследовании сообщалось, что липосомальные частицы (бисфосфонаты), несущие иммуноактивирующие молекулы размером ~80 нм, дольше оставались в кровотоке на животных моделях, чем частицы размером 100 нм (рис.
8А) (78). Однако то же исследование показало более сильный иммунный ответ на более крупные частицы, поскольку большие липосомы (~200 нм) были более склонны вызывать выработку цитокинов (ИЛ-1β, ФНО-α, ИФН-γ, ИЛ-6, ИЛ-8, и IL-10), чем небольшие липосомы (~ 80 нм) in vitro и in vivo (78). Наконец, размер важен для эффективности вакцины, потому что большие частицы могут просто содержать больше антигена, чем более крупные частицы. Частицы PLGA размером 40, 100 и 200 нм вводили мышам подкожно, и, хотя их поглощение лимфатическими ДК показало в два раза более высокое поглощение для более мелких частиц 40 нм, ДК, несущие частицы 200 нм, показали трехкратное увеличение. более высокий уровень относительного содержания антигена (180).Основываясь на различной передаче сигналов MAMP и DAMP, DC могут различать различные патогены и другие иммунные проблемы. Это позволяет DC управлять CD4 + Т-клеток в сторону разных подмножеств: Th2 или Th3 (182). Клетки Th2 секретируют провоспалительные цитокины, такие как IFN-γ и TNF-α, для активации экспрессии MHC и антимикробных ответов в фагоцитах.
Клетки Th3 секретируют больше противовоспалительных цитокинов, включая L-4, IL-5, IL-10 и IL-13, для активации ответов против внеклеточных патогенов посредством переключения класса и продукции антител B-клетками. Более того, IL-10, продуцируемый клетками Th3, ингибирует функцию и развитие клеток Th2 и макрофагов (182). Из-за этих различных цитокинов Th2-клетки способствуют иммунным реакциям, которые способствуют элиминации микробов и вирусов (10), в то время как Th3-реакции в большей степени направлены на борьбу с крупными паразитами, такими как гельминты и черви (182). В дополнение к распознаванию различных патогенов на основе передачи сигналов MAMP и DAMP, как обсуждалось выше, иммунные фагоциты могут различать различные типы патогенов на основе формы и размера, и это играет роль в дифференцировке Т-клеток (рис. 8B) (182). Хотя очень маленькие частицы золота (2 нм; размер белка) неэффективно активируют ДК, поскольку они диффундируют в клетки и попадают в немембранные компартменты, частицы размером 12 нм могут проникать в ДК через некоторую форму рецептор-опосредованного эндоцитоза и способствуют дифференцировке Th2 DCs (181). Более того, частицы полистирола, конъюгированные с OVA, размером ~50 нм с большей вероятностью инициируют активацию Th2 у мышей, чем частицы микрометрового размера (183, 184), тогда как частицы микрометрового размера более склонны вызывать Th3 ответы (185, 186). В соответствии с этими выводами, круглые конидии меньшего размера Aspergillus fumigatus или дрожжевая форма C. albicans искажают иммунные ответы моноцитов в сторону Th2, в то время как более крупные формы гиф этих патогенов вызывают больше ответов Th3 (рис. 8B) (рис. 8B) (рис. 8B). 10). Однако это также зависит от формы и топологии частиц, особенно от того, активирована ли инфламмасома, как обсуждалось выше, потому что, например, наношипы на поверхности TiO 2 частиц вызывали Th2-ответы у мышей, несмотря на большой микрометровый размер этих частиц (рис. 8В) (164).Перспективы на будущее
Очевидный вывод в этом обзоре заключается в том, что прогнозы, полученные на основе моделирования и теоретического моделирования, не всегда согласуются с экспериментальными данными, особенно в отношении режима поглощения удлиненных сплюснутых или вытянутых частиц.
Как обсуждалось выше, большинство теоретических исследований предсказывают, что такие продолговатые частицы поглощаются путем максимизации площади контакта между плазматической мембраной и частицей (режим укладки для вытянутых частиц; режим кончиком вперед для сплюснутых частиц) (34–41). Однако, как также обсуждалось выше, это, по-видимому, прямо противоречит экспериментальным данным с модельными частицами и грибковыми патогенами, образующими дефис, и нитчатыми бактериями (7, 9)., 10, 23, 25–33, 93, 94, 96–98, 119, 181). Причиной этого несоответствия может быть то, что большинство симуляций и теоретического моделирования рассматривают только/в основном плазматическую мембрану и, следовательно, основаны на предположении, что энергия, необходимая для ремоделирования мембраны, ограничивает скорость поглощения. Однако, как объясняется в этом обзоре, различные формы эндоцитоза и фагоцитоза гораздо сложнее, и это предположение может оказаться неверным. В будущем более сложные модели должны включать эту сложность и, например, учитывать региональную передачу сигналов с помощью рецепторов захвата, локальную активацию Rho-GTPases и поляризацию и активное ремоделирование актинового цитоскелета.Как обсуждалось в этом обзоре, многие белки и липиды, участвующие в передаче сигналов и закреплении цитоскелета, чувствительны к искривлению мембраны (129, 130, 156, 157, 187-189). Следовательно, области на частицах с высокой положительной или отрицательной кривизной вполне могут приводить к фагосомным доменам белково-липидных взаимодействий и передачи сигналов. Эти домены могут быть временными, учитывая высокодинамичный характер эндоцитоза и фагоцитоза. Чтобы облегчить изучение этого, некоторые подходы сосредоточены на создании стабильной кривизны мембраны, индуцированной топографией поверхности, с которой соприкасаются клетки (рис. 9).) (188–192). Методы, основанные на нанотопографии, такие как электронно-лучевая (электронно-лучевая) литография, могут использоваться для создания наноструктур размером от 50 до 100 нм, что позволяет создавать мембраны с большой кривизной (188–190). Эти подходы могут быть использованы для изучения широкого круга вопросов, связанных с индуцированным искривлением распределением клатрин-зависимых эндоцитарных белков (187), полимеризацией F-актина (188, 189) и деформациями ядерной мембраны, влияющими на распределение хроматина и экспрессию генов (190).
). Используя этот подход, было обнаружено, что нанотопографии размером более 30 нм воспринимаются макрофагами, а печатные узоры высотой более 71 нм вызывают более высокую эндоцитарную активность (19).3). Кроме того, микропечать позволяет создавать смежные структуры с разными размерами и топологией для сравнения их друг с другом с помощью конфокальной или электронной микроскопии (рис. 9В) (187, 190). Например, одно исследование с использованием такого градиента топологий показало, что родственные клатрину белки предпочитают положительную кривизну с радиусом <200 нм (187). Кроме того, массив нанометровых стержнеобразных структур с разным диаметром кончиков был использован, чтобы показать, что только кривизна в диапазоне ~100–400 нм может запускать рекрутирование белка BAR FBP17 и комплекса Arp2/3, необходимого для Сборка F-актина (188). Интересной модификацией этого метода является разработка импринтированных светочувствительных полимерных структур на основе азобензола, способных динамически изменять свою форму с помощью света, тем самым оказывая силы и влияя на кривизну мембран клеток, взаимодействующих с этими структурами (рис. 9).С) (189). Используя этот подход, было показано, что индуцированное светом удлинение вертикальных столбиков, вызывающее более высокую кривизну клеточной мембраны на кончиках столбиков, способствует локальному накоплению комплекса актинового зародыша Arp2/3 и F-действующей сборки (189). ). Этот метод может вдохновить на новые подходы к динамическому управлению кривизной мембраны и анализу реакции белков в режиме реального времени.Рисунок 9 Топологии наноструктур для изучения влияния кривизны на мембрану. (A) Изготовление наноструктур с использованием электронно-лучевой (E-beam) литографии. Столбы, стержни и C-образные шаблоны могут быть разработаны для создания библиотеки положительных и отрицательных кривизн и плоских поверхностей. Библиотеки кривизны, такие как нано-полоски, можно использовать для картирования белков, чувствительных к кривизне, при клатрин-опосредованном эндоцитозе. Отношение конца к центру выше для маркеров, локализованных в основном в областях высокой кривизны.
Сигнатуру кривизны можно использовать для картирования зависимого от кривизны рекрутирования белка во времени. (B) Клетки можно культивировать на структурированных субстратах для наноразмерных манипуляций с кривизной мембраны. Меньший диаметр столба приводит к большей кривизне. Положительная кривизна с радиусом <200 нм предпочтительнее для компонентов клатрин-опосредованного эндоцитоза и F-актина. (C) Азополимеры могут менять форму от вертикального столба до вытянутого нано-стержня под действием света и могут использоваться для динамического управления кривизной мембраны. Растяжение мембраны на концах нанополос запускает нуклеацию F-актина.Другие достижения в области материаловедения и микроскопии также приведут к разработке более совершенных инструментов для изучения того, как размер и форма частиц влияют на различные формы эндоцитоза и фагоцитоза. Современные синтетические методы позволяют создавать функциональные коллоидные строительные блоки и определять общие принципы самосборки (194).
Форма, активность и пятнистость частиц могут быть созданы путем сочетания классической золь-гель химии с недавно разработанной методологией микроэмульгирования (195). Эти подходы позволят спроектировать и изготовить биосовместимые частицы, которые будут служить моделью фагоцитарного груза с заданной кривизной и симметрией. Конструкторские грузы можно использовать для разрешения переориентации частиц внутри клеток, неоднородностей фагосомной мембраны, участков мембранного переноса и расположения цитоскелета. Стохастическая оптическая реконструктивная микроскопия со сверхвысоким разрешением (STORM), микроскопия истощения стимулированного излучения (STED) (128) и микроскопия резонанса Фёрстера с передачей энергии и флуоресценцией при жизни (FRET-FLIM) (19)6, 197) было бы полезно для определения неоднородностей мембран, наличия маркеров созревания и перестроек цитоскелета на поверхности коллоидов неправильной формы, поглощенных фагоцитами. Сочетание световой листовой флуоресцентной микроскопии с атомно-силовой микроскопией (АСМ) позволяет одновременно визуализировать фагоцитоз с измерением силы (198) и, следовательно, позволит ответить на вопрос, как размер и форма частиц влияют на силы, действующие на клетку. Новые алгоритмы машинного обучения обработки изображений и методы цифровой голографической визуализации (128, 199) набирают обороты для визуализации положения и ориентации коллоидов в 3D из 2D-изображений. Фагосомы можно выделить из клеток с помощью центрифугирования в градиенте плотности или с помощью магнитных частиц, позволяющих проводить протеомный анализ. Такой протеомный анализ различных морфотипов патогенных грибов, взаимодействующих с иммунными клетками хозяина, может пролить больше света на ключевые чувствительные к искривлению белки, на которые полагаются фагоциты для взаимодействия с захватчиками (14).В дополнение к этим экспериментам с клетками, эксперименты с искусственными мембранами могут быть полезными и восполнить пробел в моделировании и теории. Например, частицы с различной кривизной потенциально могут быть инкапсулированы в искусственные мембраны для определения организации мембраны (9).0649, т.е. , инкапсулированием флуоресцентных аналогов липидов, таких как фосфоинозитиды, вокруг коллоидов, например, центрифугированием коллоидов через липид/масляную суспензию поверх водного буфера (200)).
Эти искусственные липидные мембраны можно использовать для изучения потенциального локального связывания рекомбинантно очищенных белков, которые связывают определенные виды липидов (например, домены гомологии плекстрина) или чувствительные к кривизне мембраны (например, белки домена BAR).Текущие исследования в основном используют либо искусственные коллоиды, либо убитые микробные клетки (с помощью нагревания или химических веществ) в качестве моделей фагоцитоза, тогда как иммунная система сталкивается с широким спектром патогенов в различных формах, которые активно перемещаются (например, с помощью жгутиков) и могут реконструировать свои форма (13, 201). Как подвижность патогенов внутри фагосом влияет на их деградацию и способны ли иммунные клетки каким-то образом ощущать движение проглоченных патогенов, до конца не изучено. Как обсуждалось выше, различные патогены используют форму, чтобы избежать уничтожения иммунной системой, и многие патогенные грибы могут менять свою морфологию, например, между маленькими круглыми клетками и обширными удлиненными гифами (рис.
5В) (обзоры (13, 201)). Более того, бактерии обычного размера (например, Legionella pneumophila , Streptococcus pneumoniae , уропатогенные E. coli ) могут подвергаться филаментации, которая замедляет фагоцитоз из-за более высокой АР (23, 98, 110). Уменьшение размера также используется некоторыми бактериями, например, Moraxella catarrhalis , Neisseria meningitidis , Salmonella typhimurium и Streptococcus pneumoniae превращаются из длинных цепочек в короткие, что приводит к уменьшению площади поверхности для отложение комплемента и уменьшение зависимого от комплемента киллинга фагоцитами (110). Эти динамические изменения формы в морфологии частиц и движение частиц в фагосомах можно изучать с помощью частиц, которые могут менять свою форму, например, под действием света (189).), а также микро- и нанодвигатели с химическим приводом, соединенные с частицами для создания самодвижущихся движений в жидкостях (202, 203) и с помощью моделирования (204). Особенно интересным классом модельных фагоцитарных грузов являются самодвижущиеся коллоиды — синтетические частицы, оснащенные наномотором с химическим приводом. Двигатель, как правило, платиновый, катализирует разложение подходящего топлива, создавая локальный химический градиент, который приводит в движение частицу с помощью механизмов форетического или пузырькового движения (19).4, 205–211). Двигатели на основе гематита или диоксида титана работают аналогичным образом, однако их работой можно управлять с помощью света, что позволяет использовать удобный выключатель (212).H 2 O 2 , продуцируемый NADPH-оксидазой NOX2 в фагосомах нейтрофилов и дендритных клеток, может использоваться в качестве топлива для питания активных коллоидных частиц и создания сил в живых клетках (213, 214). Более конкретно, H 2 O 2 -топливные частицы могут быть разработаны для использования H 2 O 2 продуцируется NOX2 в просвете фагосомы для запуска движения частиц, разрыва мембраны и выхода фагоцитов (аналогично грибковым патогенам).
Более того, деформации мембраны асимметричными силами, воздействующими на мембрану фагосомы H 2 O 2 -подпитываемыми активными коллоидами, могут также приводить к переносу поляризованной мембраны. Альтернативным подходом было бы создание магнитных частиц (, например, , с ядрами Fe 2 O 3 ), чтобы обеспечить возможность внешней манипуляции с использованием магнитных полей (19).4, 205–211, 215).Биосовместимые частицы с размерами в диапазоне нанометров представляют интерес в качестве носителей для доставки лекарств или вакцинации, поэтому особый интерес представляет то, как их размер, форма, жесткость и состав поверхности влияют на поглощение клетками (рассмотрено в (216)). Многочисленные исследования пытаются использовать частицы в организме для доставки лекарств, вакцин, агентов визуализации или ДНК или РНК для генной терапии (обзор (66, 164, 176, 181, 183, 184, 217)). Теоретическое моделирование и эксперименты, посвященные тому, как форма и размер частиц влияют на клеточное поглощение и (иммунный ответ), могут помочь в разработке таких клинических агентов на основе частиц (35–41).
Заключительные замечания
— Теоретическое моделирование в сочетании с экспериментами по форме, AR и размеру патогена может вдохновить на создание частиц для вакцинации с высоким поглощением антигена или длительным пребыванием в кровотоке.
— Будущие теоретические исследования должны включать локальную передачу сигналов рецепторов и цитоскелета.
— Размер и форма патогенов влияют на активацию антигенпрезентирующих клеток и могут искажать ответы Т-клеток на ответы Th2 и Th3.
— Шероховатые и остроконечные частицы, а также частицы с высоким AR могут вызывать активацию воспаления и гибель клеток.
— Патогены изменяют свою форму, чтобы ускользнуть от иммунной системы, и это потенциально может быть использовано в вакцинах.
— Молекулярные наномоторы можно использовать для изучения того, как подвижность патогенов и изменение формы влияют на фагоцитоз и иммунные реакции.
— Методы нанесения рисунков, напечатанных на подложках, основанные на нанотопографии, могут использоваться для детального понимания того, как кривизна влияет на фагоцитоз и различные формы эндоцитоза.
Вклад автора
MB написал рукопись при поддержке MK, SS, ST и GB. Все авторы внесли свой вклад в статью и одобрили представленную версию.
Финансирование
ST, SS и GB финансируются за счет гранта для молодых исследователей от Human Frontier Science Program (HFSP; RGY0080/2018). Великобритания получила финансирование от Европейского исследовательского совета (ERC) в рамках исследовательской и инновационной программы Horizon 2020 Европейского Союза (грантовое соглашение № 862137) и Нидерландской организации научных исследований (грант Vidi NWO-ALW VIDI 864.14.001). ST также выражает благодарность Департаменту атомной энергии правительства Индии за поддержку в рамках проекта №. 12-R&D-TFR-5.04-0800 и 12-D&D-TFR-5.10-1100, Фонд Саймонса (грант № 287975) и Общество Макса Планка через Партнерскую группу Макса Планка.
Конфликт интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Дополнительный материал
Дополнительный материал к этой статье можно найти в Интернете по адресу: https://www.frontiersin.org/articles/10.3389/fimmu.2020.607945/full#supplementary-material
Ссылки
1. Flannagan RS, Jaumouille V, Grinstein S. Клеточная биология фагоцитоза. Annu Rev Pathol (2012) 7:61–98. doi: 10.1146/annurev-pathol-011811-132445
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
2. Богданович М., Гринштейн С. Роль фосфолипидов в эндоцитозе, фагоцитозе и макропиноцитозе. Physiol Rev (2013) 93(1):69–106. doi: 10.1152/physrev.00002.2012
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
3. Суонсон Дж.А. Формирование чашечек в фагосомы и макропиносомы. Nat Rev Mol Cell Biol (2008) 9(8):639–49. doi: 10.1038/nrm2447
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
4. Cox D, Tseng CC, Bjekic G, Greenberg S. Потребность в фосфатидилинозитол-3-киназе при расширении ложноножек.
J Biol Chem (1999) 274(3):1240–7. doi: 10.1074/jbc.274.3.1240PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
5. Schlam D, Bagshaw RD, Freeman SA, Collins RF, Pawson T, Fairn GD, et al. Фосфоинозитид-3-киназа обеспечивает фагоцитоз крупных частиц путем прекращения сборки актина через белки, активирующие GTPase Rac/Cdc42. Nat Commun (2015) 6(1):8623. doi: 10.1038/ncomms9623
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
6. Бранцк Н., Любоемска А., Хардисон С.Е., Ван К., Гутьеррес М.Г., Браун Г.Д. и др. Нейтрофилы чувствуют размер микробов и избирательно высвобождают нейтрофильные внеклеточные ловушки в ответ на крупные патогены. Nat Immunol (2014) 15(11):1017–25. doi: 10.1038/ni.2987
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
7. Льюис Л.Э., Бейн Дж.М., Лоус С., Гиллеспи С., Рудкин Ф.М., Гоу Н.А. и др. Стадийно-специфическая оценка фагоцитоза Candida albicans макрофагами идентифицирует состав клеточной стенки и морфогенез как ключевые детерминанты.
PloS Pathog (2012) 8(3):e1002578. doi: 10.1371/journal.ppat.1002578PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
8. Дагган С., Эссиг Ф., Хюннигер К., Мохтари З., Бауэр Л., Ленерт Т. и соавт. Активация нейтрофилов Candida glabrata, но не Candida albicans, способствует поглощению грибков моноцитами. (2015) 17(9):1259–76. doi: 10.1111/cmi.12443
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
9. Reedy JL, Negoro PE, Feliu M, Lord AK, Khan NS, Lukason DP, et al. Рецептор углеводного лектина Dectin-1 опосредует иммунный ответ на Exserohilum rostratum. Заразить иммунитет (2017) 85(3):e00903-16. doi: 10.1128/IAI.00903-16
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
10. Heung LJ. Моноциты и ответ хозяина на грибковые патогены. Front Cell Infect Microbiol (2020) 10:34. doi: 10.3389/fcimb.2020.00034
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
11.
Вьяс Дж. М., Ван дер Вин А. Г., Плоех Х. Л. Известные неизвестные процессы процессинга и презентации антигена. Nat Rev Immunol (2008) 8(8):607–18. дои: 10.1038/nri2368Резюме PubMed | Полный текст перекрестной ссылки | Google Scholar
12. Гротцке Дж. Э., Сенгупта Д., Лу К., Крессуэлл П. Непрекращающаяся сага о механизме (механизмах) ограниченной перекрестной презентации MHC класса I. Curr Opin Immunol (2017) 46:89–96. doi: 10.1016/j.coi.2017.03.015
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
13. Эрвиг Л.П., Гоу Н.А. Взаимодействие грибковых возбудителей с фагоцитами. Nat Rev Microbiol (2016) 14(3):163–76. doi: 10.1038/nrmicro.2015.21
Резюме PubMed | Полный текст перекрестной ссылки | Google Scholar
14. Крюгер Т., Луо Т., Шмидт Х., Шопова И., Книмейер О. Проблемы и стратегии протеомного анализа взаимодействия патогенных грибов человека с иммунными клетками хозяина. Протеомы (2015) 3(4):467–95.
doi: 10.3390/proteomes3040467PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
15. Сукхитасри В., Ниша Н., Бисвас Л., Анил Кумар В., Бисвас Р. Врожденное иммунное распознавание компонентов микробной клеточной стенки и микробные стратегии уклонения от такого распознавания. Microbiol Res (2013) 168(7):396–406. doi: 10.1016/j.micres.2013.02.005
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
16. Steele C, Rapaka RR, Metz A, Pop SM, Williams DL, Gordon S, et al. Дектин-1 рецептора бета-глюкана распознает специфическую морфологию Aspergillus fumigatus. PloS Pathog (2005) 1(4):e42. doi: 10.1371/journal.ppat.0010042
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
17. Фриман С.А., Гринштейн С. Фагоцитоз: механосенсорика, силы тяги и молекулярная муфта. Curr Biol (2020) 30(1):R24–r26. doi: 10.1016/j.cub.2019.11.047
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
18.
Пачеко П., Уайт Д., Сульчек Т. Влияние размера микрочастиц и плотности Fc на фагоцитоз макрофагов. PloS One (2013) 8(4):e60989. doi: 10.1371/journal.pone.0060989PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
19. Vorselen D, Wang Y, de Jesus MM, Shah PK, Footer MJ, Huse M, et al. Микроскопия силы тяги микрочастиц выявляет паттерны проявления субклеточной силы во взаимодействиях иммунных клеток и мишеней. Нац коммуна (2020) 11(1):20. doi: 10.1038/s41467-019-13804-z
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
20. Бакалар М.Х., Джоффе А.М., Шмид Э.М., Сон С., Подольский М., Флетчер Д.А. Зависимая от размера сегрегация контролирует фагоцитоз макрофагов мишеней, опсонизированных антителами. Cell (2018) 174(1):131–42.e13. doi: 10.1016/j.cell.2018.05.059
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
21. Jaumouille V, Cartagena-Rivera AX, Waterman CM.
Связывание бета2-интегринов с актином с помощью механочувствительного молекулярного сцепления приводит к фагоцитозу, опосредованному комплементарными рецепторами. Nat Cell Biol (2019) 21(11):1357–69. doi: 10.1038/s41556-019-0414-2PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
22. Дональдсон К., Мерфи Ф.А., Даффин Р., Польша, Калифорния. Асбест, углеродные нанотрубки и мезотелий плевры: обзор гипотезы о роли задержки длинных волокон в париетальной плевре, воспалении и мезотелиоме. Часть Fibre Toxicol (2010) 7(1):5. doi: 10.1186/1743-8977-7-5
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
23. Prashar A, Bhatia S, Gigliozzi D, Martin T, Duncan C, Guyard C, et al. Нитевидная морфология бактерий задерживает время морфогенеза фагосом в макрофагах. J Cell Biol (2013) 203(6):1081–97. doi: 10.1083/jcb.201304095
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
24.
Huang SX, Jaurand MC, Kamp DW, Whysner J, Hei TK. Роль мутагенности в индуцированной асбестовым волокном канцерогенности и других заболеваниях. J Toxicol Environ Health B Crit Rev (2011) 14(1-4):179–245. doi: 10.1080/104.2011.556051
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
25. Champion JA, Walker A, Mitragotri S. Роль размера частиц в фагоцитозе полимерных микросфер. Фарм Рез (2008) 25(8):1815–21. doi: 10.1007/s11095-008-9562-y
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
26. Агарвал Р., Сингх В., Джерни П., Ши Л., Шринивасан С.В., Рой К. Клетки млекопитающих предпочтительно усваивают гидрогелевые нанодиски, а не наностержни, и используют специфические для формы механизмы поглощения. Proc Natl Acad Sci USA (2013) 110(43):17247–52. doi: 10.1073/pnas.1305000110
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
27.
Champion JA, Mitragotri S. Ингибирование фагоцитоза полимерных частиц, индуцированное формой. Фарм Рез (2009) 26(1):244–9. doi: 10.1007/s11095-008-9626-zPubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
28. Lu Z, Qiao Y, Zheng XT, Chan-Parka MB, Li CM. Влияние формы частиц на фагоцитоз композитов квантовых точек CdTe и цистина. Med Chem Commun (2010) 1:84–6. doi: 10.1039/c0md00008f
Полный текст CrossRef | Google Scholar
29. Paul D, Achouri S, Yoon YZ, Herre J, Bryant CE, Cicuta P. Динамика фагоцитоза зависит от формы мишени. Biophys J (2013) 105(5):1143–50. doi: 10.1016/j.bpj.2013.07.036
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
30. Sharma G, Valenta DT, Altman Y, Harvey S, Xie H, Mitragotri S, et al. Форма полимерных частиц независимо влияет на связывание и интернализацию макрофагами. J Control Release (2010) 147(3):408–12. doi: 10.1016/j.jconrel.2010.
07.116PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
31. Доши Н., Митраготри С. Макрофаги распознают размер и форму своих целей. PLoS One (2010) 5(4):e10051. doi: 10.1371/journal.pone.0010051
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
32. Tabei Y, Sugino S, Eguchi K, Tajika M, Abe H, Nakajima Y, et al. Влияние формы частиц карбоната кальция на фагоцитоз и провоспалительный ответ в дифференцированных макрофагах THP-1. Biochem Biophys Res Commun (2017) 490(2):499–505. doi: 10.1016/j.bbrc.2017.06.069
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
33. Чека С., Хиттингер М., Лер С.М., Шнайдер-Даум Н., Шнайдер М. Поглощение цилиндрических микрочастиц макрофагами исследовано с помощью коррелятивной микроскопии. Eur J Pharm Biopharm (2015) 95(Pt A):151–5. doi: 10.1016/j.ejpb.2015.03.010
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
34.
Ричардс Д.М., Эндрес Р.Г. Зависимость от формы мишени в простой модели рецептор-опосредованного эндоцитоза и фагоцитоза. Proc Natl Acad Sci USA (2016) 113(22):6113–8. doi: 10.1073/pnas.1521974113PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
35. Дэн Х., Датта П., Лю Дж. Режимы входа эллипсоидальных наночастиц на мембрану во время клатрин-опосредованного эндоцитоза. Мягкая материя (2019) 15 (25): 5128–37. doi: 10.1039/C9SM00751B
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
36. Дэн Х., Датта П., Лю Дж. Стохастическое моделирование интернализации и выведения наночастиц посредством рецептор-опосредованного трансцитоза. Nanoscale (2019) 11(23):11227–35. doi: 10.1039/C9NR02710F
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
37. Deng H, Dutta P, Liu J. Стохастическое моделирование интернализации наночастиц через клатрин-опосредованный эндоцитоз, зависимый от рецептора трансферрина.
Biochim Biophys Acta Gen Subj (2018) 1862(9):2104–11. doi: 10.1016/j.bbagen.2018.06.018PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
38. Чен Л., Сяо С., Чжу Х., Ван Л., Лян Х. Кинетика интернализации наночастиц мембранами в зависимости от формы. Мягкая материя (2016) 12 (9): 2632–41. doi: 10.1039/C5SM01869B
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
39. Shen Z, Ye H. Эффективность мембранного покрытия эластичных наночастиц во время эндоцитоза: размер и форма имеют значение. ACS Nano (2019) 13(1):215–28. doi: 10.1021/acsnano.8b05340
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
40. Xiong K, Zhao J, Yang D, Cheng Q, Wang J, Ji H. Совместная упаковка наночастиц различных размеров и форм липидными мембранами. Soft Matter (2017) 13(26):4644–52. doi: 10.1039/C7SM00345E
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
41.
Бахрами А.Х., Раатц М., Агудо-Каналехо Дж., Мишель Р., Кертис Э.М., Холл К.К. и др. Обертывание наночастиц мембранами. Adv Colloid Interface Sci (2014) 208:214–24. doi: 10.1016/j.cis.2014.02.012PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
42. Агудо-Каналехо Дж. Поглощение эллипсоидальных наночастиц мембранами: полное описание ориентационных изменений. J Phys Condens Matter (2020) 32(29):2
. doi: 10.1088/1361-648X/ab8034PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
43. Бахрами А.Х. Ориентационные изменения и нарушение интернализации эллипсоидальных наночастиц мембранами везикул. Мягкая материя (2013) 9 (36): 8642–6. doi: 10.1039/c3sm50885d
Полный текст CrossRef | Google Scholar
44. Агудо-Каналехо Дж., Липовски Р. Адгезивные наночастицы как локальные зонды кривизны мембраны. Nano Lett (2015) 15 (10): 7168–73. doi: 10.1021/acs.nanolett.5b03475
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
45.
Дасгупта С., Аут Т., Гомппер Г. Обертывание эллипсоидальных наночастиц жидкими мембранами. Мягкая материя (2013) 9 (22): 5473–82. doi: 10.1039/C3SM50351HПолный текст CrossRef | Google Scholar
46. Дасгупта С., Аут Т., Гомппер Г. Форма и ориентация имеют значение для поглощения несферических частиц клетками. Nano Lett (2014) 14 (2): 687–93. doi: 10.1021/nl403949h
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
47. Alibert C, Goud B, Manneville JB. Действительно ли раковые клетки мягче нормальных клеток? Biol Cell (2017) 109(5):167–89. doi: 10.1111/boc.201600078
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
48. Агудо-Каналехо Дж. Поглощение частиц сильно асимметричными мембранами с площадными резервуарами. Мягкая материя (2020) 17: 298–307. doi: 10.1039/C9SM02367D
Полный текст CrossRef | Google Scholar
49. Ge C, Tian J, Zhao Y, Chen C, Zhou R, Chai Z.
К пониманию короны наночастиц-белков. Arch Toxicol (2015) 89(4):519–39. doi: 10.1007/s00204-015-1458-0PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
50. Wu L, Zhang Y, Li Z, Yang G, Kochovski Z, Chen G, et al. «Сладкое» архитектурно-зависимое поглощение макрофагами имитирующих гликокаликс наночастиц на основе биоразлагаемых алифатических полиэфиров. J Am Chem Soc (2017) 139(41):14684–92. doi: 10.1021/jacs.7b07768
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
51. Kuo JC-H, Gandhi JG, Zia RN Paszek MJ. Физическая биология гликокаликса раковых клеток. Nat Phys (2018) 14(7):658–69. doi: 10.1038/s41567-018-0186-9
Полный текст CrossRef | Google Scholar
52. Мекл Л. Новая роль гликокаликса млекопитающих в организации функциональной мембраны и регуляции иммунной системы. Front Cell Dev Biol (2020) 8:253. doi: 10.3389/fcell.2020.00253
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
53.
Хосраванизаде А., Сенс П., Мохаммад-Рафи Ф. Обертывание нанопроволоки липидной мембраной на подложке. Soft Matter (2019) 15(37):7490–500. doi: 10.1039/C9SM00618DPubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
54. Хашеми С.М., Сенс П., Мохаммад-Рафи Ф. Регуляция перехода оболочки мембраны цилиндрической мишени за счет адгезии цитоскелета. JR Soc Interface (2014) 11(100):20140769. doi: 10.1098/rsif.2014.0769
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
55. Курк Т., Вирнсбергер П., Добникар Дж., Френкель Д., Шарич А. Управление транспортировкой грузов в многокомпонентных мембранах. Nano Lett (2018) 18(9):5350–6. doi: 10.1021/acs.nanolett.8b00786
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
56. Патель Б., Гупта Н., Ахсан Ф. Инженерия частиц для увеличения или уменьшения поглощения частиц альвеолярными макрофагами и влияния на терапевтический результат.
Eur J Pharm Biopharm (2015) 89:163–74. doi: 10.1016/j.ejpb.2014.12.001PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
57. Dunkelberger JR, Song WC. Комплемент и его роль во врожденных и адаптивных иммунных реакциях. Cell Res (2010) 20(1):34–50. doi: 10.1038/cr.2009.139
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
58. Jaumouillé V, Grinstein S. Молекулярные механизмы образования фагосом. Микробиолог Спектр (2016) 4(3):507–26. doi: 10.1128/9781555819194.ch37
Полный текст CrossRef | Google Scholar
59. Завери Т.Д., Льюис Дж.С., Долгова Н.В., Клэр-Зальцлер М.Дж., Кеселовский Б.Г. Интегрин-направленная модуляция ответов макрофагов на биоматериалы. Биоматериалы (2014) 35(11):3504–15. doi: 10.1016/j.biomaterials.2014.01.007
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
60. Накаяма М. Распознавание кристаллов и наночастиц макрофагами.
Фронт Иммунол (2018) 9:103. doi: 10.3389/fimmu.2018.00103PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
61. Эдвардс Д.А., Хейнс Дж., Капонетти Г., Хркач Дж., Бен-Джебриа А., Эскью М.Л. и соавт. Крупные пористые частицы для доставки лекарств в легкие. Наука (1997) 276 (5320): 1868–71. doi: 10.1126/science.276.5320.1868
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
62. Макино К., Ямамото Н., Хигучи К., Харада Н., Ошима Х., Терада Х. Фагоцитарное поглощение полистироловых микросфер альвеолярными макрофагами: влияние размера и свойств поверхности микросфер. Colloids Surf B: Biointerfaces (2003) 27(1):33–9. doi: 10.1016/S0927-7765(02)00042-5
Полный текст CrossRef | Google Scholar
63. Hirota K, Hasegawa T, Hinata H, Ito F, Inagawa H, Kochi C, et al. Оптимальные условия для эффективного фагоцитоза микросфер PLGA, нагруженных рифампицином, альвеолярными макрофагами. J Control Release (2007) 119(1):69–76.
doi: 10.1016/j.jconrel.2007.01.013PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
64. Чемпион Дж. А., Митраготри С. Роль геометрии мишени в фагоцитозе. Proc Natl Acad Sci USA (2006) 103(13):4930–4. doi: 10.1073/pnas.0600997103
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
65. Польша CA, Duffin R, Kinloch I, Maynard A, Wallace WAH, Seaton A, et al. Углеродные нанотрубки, введенные в брюшную полость мышей, показали асбестоподобную патогенность в пилотном исследовании. Nat Nanotechnol (2008) 3(7):423–8. doi: 10.1038/nnano.2008.111
Резюме PubMed | Полный текст перекрестной ссылки | Google Scholar
66. Tabata Y, Inoue Y, Ikada Y. Влияние размера на системные и слизистые иммунные реакции, вызванные пероральным введением биоразлагаемых микросфер. Вакцина (1996) 14(17-18):1677–85. doi: 10.1016/S0264-410X(96)00149-1
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
67.
Герант М., Генрих В., Дембо М. Механика фагоцитоза нейтрофилов: поведение коркового напряжения. J Cell Sci (2005) 118 (Pt 9): 1789–97. doi: 10.1242/jcs.02275PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
68. Эванс Э.А., Во Р., Мельник Л. Модуль упругой сжимаемости мембраны эритроцитов. Biophys J (1976) 16(6):585–95. doi: 10.1016/S0006-3495(76)85713-X
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
69. Циммерберг Дж., Козлов М.М. Как белки вызывают искривление клеточной мембраны. Nat Rev Mol Cell Biol (2006) 7(1):9–19. doi: 10.1038/nrm1784
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
70. Petty HR, Hafeman DG, McConnell HM. Исчезновение складок поверхности макрофагов после антителозависимого фагоцитоза. J Cell Biol (1981) 89(2):223–9. doi: 10.1083/jcb.89.2.223
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
71.
Hirsch JG, Cohn ZA. Дегрануляция полиморфноядерных лейкоцитов после фагоцитоза микроорганизмов. J Exp Med (1960) 112(6):1005–14. doi: 10.1084/jem.112.6.1005PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
72. Сузаки Э., Кобаяши Х., Кодама Ю., Масуджима Т., Теракава С. Динамика скорости экзоцитоза, связанная с фагоцитозом нейтрофилов. Cell Motil Cytoskeleton (1997) 38(3):215–28. doi: 10.1002/(SICI)1097-0169(1997)38:3<215::AID-CM1>3.0.CO;2-4
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
73. Jaumouillé V, Waterman CM. Физические ограничения и силы, участвующие в фагоцитозе. Фронт Иммунол (2020) 11:1097. doi: 10.3389/fimmu.2020.01097
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
74. Westman J, Walpole GFW, Kasper L, Xue BY, Elshafee O, Hube B, et al. Слияние лизосом поддерживает целостность фагосом во время грибковой инфекции. Микроб-хозяин клетки (2020) 28(6):798–812.
e6. doi: 10.1016/j.chom.2020.09.004PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
75. Hochmuth FM, Shao JY, Dai J, Sheetz MP. Деформация и затекание мембраны в нити, извлеченные из конусов роста нейронов. Biophys J (1996) 70(1):358–69. doi: 10.1016/S0006-3495(96)79577-2
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
76. Мастерс Т.А., Понтес Б., Вяснофф В., Ли И., Готье Н.К. Натяжение плазматической мембраны управляет перемещением мембраны, ремоделированием цитоскелета и биохимической передачей сигналов во время фагоцитоза. Proc Natl Acad Sci США (2013) 110(29):11875–80. doi: 10.1073/pnas.1301766110
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
77. Chono S, Tanino T, Seki T, Morimoto K. Влияние размера частиц на доставку лекарственного средства в альвеолярные макрофаги крыс после легочного введения ципрофлоксацина, включенного в липосомы. J Drug Target (2006) 14(8):557–66.
doi: 10.1080/10611860600834375PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
78. Epstein-Barash H, Gutman D, Markovsky E, Mishan-Eisenberg G, Koroukhov N, Szebeni J, et al. Физико-химические параметры, влияющие на биоактивность липосомальных бисфосфонатов для терапии рестеноза: интернализация, ингибирование клеток, активация цитокинов и комплемента, механизм гибели клеток. J Control Release (2010) 146(2):182–95. doi: 10.1016/j.jconrel.2010.03.011
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
79. Линь С.Ю., Хсу В.Х., Ло Дж.М., Цай Х.К., Сюэ Г.Х. Наноносители с новой геометрией смягчают фагоцитоз при доставке лекарств. J Control Release (2011) 154(1):84–92. doi: 10.1016/j.jconrel.2011.04.023
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
80. Leclerc L, Rima W, Boudard D, Pourchez J, Forest V, Bin V, et al. Размер субмикрометровых и нанометровых частиц влияет на клеточное поглощение и биологическую активность макрофагов in vitro.
Inhal Toxicol (2012) 24(9):580–8. doi: 10.3109/08958378.2012.699984PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
81. Nabeshi H, Yoshikawa T, Matsuyama K, Nakazato Y, Arimori A, Isobe M, et al. Зависящие от размера цитотоксические эффекты наночастиц аморфного кремнезема на клетки Лангерганса. Pharmazie (2010) 65(3):199–201.
Резюме PubMed | Google Scholar
82. Mathaes R, Winter G, Besheer A, Engert J. Влияние геометрии частиц и пегилирования на фагоцитоз частиц-носителей. Int J Pharm (2014) 465(1-2):159–64. doi: 10.1016/j.ijpharm.2014.02.037
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
83. Читрани Б.Д., Газани А.А., Чан В.К. Определение зависимости от размера и формы поглощения наночастиц золота клетками млекопитающих. Nano Lett (2006) 6 (4): 662–8. doi: 10.1021/nl052396o
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
84.
Lu Z, Qiao Y, Zheng XT, Chan-Parka MB, Li CM. Влияние формы частиц на фагоцитоз композитов квантовых точек CdTe и цистина. Med Chem Commun (2010) 1:84–6. doi: 10.1039/c0md00008fПолный текст CrossRef | Google Scholar
85. Herd H, Daum N, Jones AT, Huwer H, Ghandehari H, Lehr CM. Геометрия наночастиц и ориентация поверхности влияют на способ клеточного поглощения. ACS Nano (2013) 7 (3): 1961–73. doi: 10.1021/nn304439f
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
86. Yoo JW, Mitragotri S. Полимерные частицы, которые меняют форму в ответ на раздражитель. Proc Natl Acad Sci USA (2010) 107(25):11205–10. doi: 10.1073/pnas.1000346107
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
87. Buckley CM, Pots H, Gueho A, Vines JH, Munn CJ, Phillips BA, et al. Координированная активность Ras и Rac формирует чашечки макропиноцитов и обеспечивает фагоцитоз геометрически разнообразных бактерий.
Карр Биол (2020) 30(15):2912–26.e5. doi: 10.1016/j.cub.2020.05.049PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
88. Mischler SE, Cauda EG, Di Giuseppe M, McWilliams LJ, St Croix C, Sun M, et al. Дифференциальная активация макрофагов RAW 264.7 сегрегированным по размеру кристаллическим кремнеземом. J Occup Med Toxicol (2016) 11:57. doi: 10.1186/s12995-016-0145-2
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
89. Al-Rawi M, Diabate S, Weiss C. Поглощение и внутриклеточная локализация субмикронных и наноразмерных частиц SiO(2) в клетках HeLa. Arch Toxicol (2011) 85(7):813–26. doi: 10.1007/s00204-010-0642-5
Резюме PubMed | Полный текст перекрестной ссылки | Google Scholar
90. Höfs S, Mogavero S, Hube B. Взаимодействие Candida albicans с клетками-хозяевами: факторы вирулентности, защита хозяина, стратегии побега и микробиота. J Microbiol (2016) 54(3):149–69.
doi: 10.1007/s12275-016-5514-0PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
91. Дуглас Л.М., Конопка Дж.Б. Организация плазматической мембраны способствует вирулентности грибкового патогена человека Candida albicans. J Microbiol (Сеул, Корея) (2016) 54(3):178–91. doi: 10.1007/s12275-016-5621-y
CrossRef Полный текст | Google Scholar
92. Ray SC, Rappleye CA. Полет под радаром: Histoplasma capsulatum избегает распознавания врожденным иммунитетом. Semin Cell Dev Biol (2019) 89:91–8. doi: 10.1016/j.semcdb.2018.03.009
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
93. Bain JM, Louw J, Lewis LE, Okai B, Walls CA, Ballou ER, et al. Образование гиф Candida albicans и маскирование бета-глюкана маннаном ингибируют созревание фагосом макрофагов. mBio (2014) 5(6):e01874. doi: 10.1128/mBio.01874-14
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
94.
Вылкова С., Лоренц М.С. Нейтрализация фагосом грибковым патогеном Candida albicans вызывает пироптоз макрофагов. Infect Immun (2017) 85(2):e00832-16. doi: 10.1128/IAI.00832-16PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
95. Корнитцер Д. Регуляция морфогенеза гиф Candida albicans с помощью эндогенных сигналов. J Fungi (Базель) (2019) 5(1):21. doi: 10.3390/jof5010021
Полный текст CrossRef | Google Scholar
96. Веллингтон М., Косельный К., Суттервала Ф.С., Крысан Д.Дж. Candida albicans запускает NLRP3-опосредованный пироптоз в макрофагах. Эукариотическая клетка (2014) 13(2):329–40. doi: 10.1128/EC.00336-13
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
97. Heinsbroek SE, Kamen LA, Taylor PR, Brown GD, Swanson J, Gordon S. Привлечение актина и фосфоинозитида к полностью сформированным фагосомам Candida albicans в макрофагах мыши. J Innate Immun (2009) 1(3):244–53.
doi: 10.1159/000173694PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
98. Möller J, Luehmann T, Hall H, Vogel V. Гонка к полюсу: как форма с высоким соотношением сторон и гетерогенная среда ограничивают фагоцитоз нитчатых бактерий Escherichia coli макрофагами. Nano Lett (2012) 12(6):2901–5. doi: 10.1021/nl3004896
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
99. Толлис С., Дарт А.Е., Циркотис Г., Эндрес Р.Г. Механизм застежки-молнии при фагоцитозе: энергетические потребности и вариабельность формы фагоцитарной чашечки. BMC Syst Biol (2010) 4:149. doi: 10.1186/1752-0509-4-149
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
100. McKenzie CG, Koser U, Lewis LE, Bain JM, Mora-Montes HM, Barker RN, et al. Вклад компонентов клеточной стенки Candida albicans в распознавание и выход из мышиных макрофагов. Заразить иммунитет (2010) 78(4):1650–8. doi: 10.
1128/IAI.00001-10PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
101. Хроматка Б.С., Ноубл С.М., Джонсон А.Д. Транскрипционный ответ Candida albicans на оксид азота и роль гена YHB1 в нитрозативном стрессе и вирулентности. Mol Biol Cell (2005) 16(10):4814–26. doi: 10.1091/mbc.e05-05-0435
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
102. Koch B, Barugahare AA, Lo TL, Huang C, Schittenhelm RB, Powell DR, et al. Метаболическая контрольная точка для переключателя развития дрожжей на гифы, регулируемого сигнализацией эндогенного оксида азота. Cell Rep (2018) 25(8):2244–58.e7. doi: 10.1016/j.celrep.2018.10.080
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
103. Harcus D, Nantel A, Marcil A, Rigby T, Whiteway M. Профилирование транскрипции передачи сигналов циклического AMP у Candida albicans. Mol Biol Cell (2004) 15(10):4490–9. doi: 10.1091/mbc.e04-02-0144
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
104.
Nasution O, Srinivasa K, Kim M, Kim YJ, Kim W, Jeong W, et al. Перекись водорода индуцирует дифференцировку гиф у Candida albicans. Eukaryot Cell (2008) 7(11):2008–11. doi: 10.1128/EC.00105-08PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
105. Rossi DCP, Gleason JE, Sanchez H, Schatzman SS, Culbertson EM, Johnson CJ, et al. Candida albicans FRE8 кодирует представителя семейства NADPH-оксидаз, который вызывает всплеск АФК во время морфогенеза грибов. PloS Pathog (2017) 13(12):e1006763. doi: 10.1371/journal.ppat.1006763
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
106. Davis D. Адаптация Candida albicans к pH окружающей среды и ее связь с патогенезом. Curr Genet (2003) 44(1):1–7. doi: 10.1007/s00294-003-0415-2
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
107. Стеркель А.К., Меттельман Р., Вутрих М., Кляйн Б.С. Недооцененный внутриклеточный образ жизни Blastomyces dermatitidis.
J Immunol (2015) 194(4):1796–805. doi: 10.4049/jimmunol.1303089PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
108. Науфер А., Иполито В.Э.Б., Ганесан С., Прашар А., Заремберг В., Ботельо Р.Дж. и соавт. рН эндофагосом контролирует ассоциацию их мембран с уровнями Vps34 и PtdIns(3)P. J Cell Biol (2018) 217(1):329–46. doi: 10.1083/jcb.201702179
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
109. Джастис С.С., Хунстад Д.А., Цегельски Л., Халтгрен С.Дж. Морфологическая пластичность как стратегия выживания бактерий. Nat Rev Microbiol (2008) 6(2):162–8. doi: 10.1038/nrmicro1820
Резюме PubMed | Полный текст перекрестной ссылки | Google Scholar
110. Ян Д.К., Блэр К.М., Салама Н.Р. Оставаться в форме: влияние формы клеток на выживание бактерий в различных средах. Microbiol Mol Biol Rev (2016) 80(1):187–203. doi: 10.1128/MMBR.00031-15
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
111.
Johnston SA, May RC. Взаимодействия криптококка с макрофагами: уклонение от фагосомы и манипулирование ею грибковым патогеном. Cell Microbiol (2013) 15(3):403–11. doi: 10.1111/cmi.12067PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
112. Возняк К.Л., Левитц С.М. Cryptococcus neoformans проникает по эндолизосомному пути дендритных клеток и уничтожается лизосомальными компонентами. Infect Immun (2008) 76(10):4764–71. doi: 10.1128/IAI.00660-08
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
113. Хьюстон С.М., Ли С.С., Стек Д., Тимм-Макканн М., Джонс Г.Дж., Ислам А. и др. Cryptococcus gattii уничтожается дендритными клетками, но уклоняется от адаптивного иммунитета, не индуцируя созревание дендритных клеток. J Immunol (2013) 191(1):249–61. doi: 10.4049/jimmunol.1202707
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
114. Эрнандес-Чавес М.Дж., Перес-Гарсия Л.
А., Нино-Вега Г.А., Мора-Монтес Х.М. Грибковые стратегии уклонения от распознавания иммунитетом хозяина. J Fungi (2017) 3(4):55. doi: 10.3390/jof3040051Полный текст CrossRef | Google Scholar
115. Almeida AJ, Cunha C, Carmona JA, Sampaio-Marques B, Carvalho A, Malavazi I, et al. Cdc42p контролирует форму дрожжевых клеток и вирулентность Paracoccidioides brasiliensis. Fungal Genet Biol (2009) 46(12):919–26. doi: 10.1016/j.fgb.2009.08.004
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
116. Clarke M, Engel U, Giorgione J, Muller-Taubenberger A, Prassler J, Veltman D, et al. Распознавание кривизны и генерация силы при фагоцитозе. BMC Biol (2010) 8:154. doi: 10.1186/1741-7007-8-154
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
117. Либл Д., Гриффитс Г. Временная сборка F-актина фагосомами задерживает слияние фагосом с лизосомами в макрофагах, перегруженных грузом. J Cell Sci (2009) 122 (Pt 16): 2935–45.
doi: 10.1242/jcs.048355PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
118. Баранов М.В., Ревело Н.Х., Дингжан И., Мараспини Р., Тер Беест М., Хонигманн А. и др. SWAP70 организует актиновый цитоскелет и необходим для фагоцитоза. Cell Rep (2016) 17(6):1518–31. doi: 10.1016/j.celrep.2016.10.021
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
119. Okai B, Lyall N, Gow NA, Bain JM, Erwig LP. Rab14 регулирует созревание фагосом макрофагов, содержащих грибковый патоген Candida albicans, и исход взаимодействия хозяин-патоген. Infect Immun (2015) 83(4):1523–35. doi: 10.1128/IAI.02917-14
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
120. Гринберг С., Эль Хури Дж., Ди Вирджилио Ф., Каплан Э.М., Сильверстайн С.К. Са(2+)-независимая сборка и разборка F-актина во время опосредованного рецептором Fc фагоцитоза в макрофагах мыши. J Cell Biol (1991) 113(4):757–67.
doi: 10.1083/jcb.113.4.757PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
121. Kjeken R, Egeberg M, Habermann A, Kuehnel M, Peyron P, Floetenmeyer M, et al. Слияние фагосом, ранних и поздних эндосом: роль актина в слиянии поздних, но не ранних эндоцитарных органелл. Mol Biol Cell (2004) 15(1):345–58. doi: 10.1091/mbc.e03-05-0334
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
122. Yam PT, Theriot JA. Повторяющиеся циклы быстрой сборки и разборки актина на фагосомах эпителиальных клеток. Mol Biol Cell (2004) 15(12):5647–58. doi: 10.1091/mbc.e04-06-0509
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
123. Баргер С.Р., Готье Н.К., Крендель М. Сжимание еды: функции миозина в фагоцитозе. Trends Cell Biol (2020) 30(2):157–67. doi: 10.1016/j.tcb.2019.11.002
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
124. Bain J, Gow NA, Erwig LP.
Новое понимание взаимодействия хозяина и грибкового патогена, полученное с помощью визуализации живых клеток. Semin Immunopathol (2015) 37(2):131–9. doi: 10.1007/s00281-014-0463-3PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
125. Фельдман М.Б., Вьяс Дж.М., Мансур М.К. Требуется деревня: фагоциты играют центральную роль в грибковом иммунитете. Semin Cell Dev Biol (2019) 89:16–23. doi: 10.1016/j.semcdb.2018.04.008
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
126. Левин Р., Гринштейн С., Кантон Дж. Жизненный цикл фагосом: формирование, созревание и разрешение. Immunol Rev (2016) 273(1):156–79. doi: 10.1111/imr.12439
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
127. Фриман С.А., Гринштейн С. Фагоцитоз: рецепторы, интеграция сигналов и цитоскелет. Immunol Rev (2014) 262(1):193–215. doi: 10.1111/imr.12212
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
128.
Баранов М.В., Олеа Р.А., ван ден Богарт Г. В погоне за поглощением: микроскопия сверхвысокого разрешения при эндоцитозе и фагоцитозе. Trends Cell Biol (2019) 29(9):727–39. doi: 10.1016/j.tcb.2019.05.006PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
129. McMahon HT, Boucrot E. Кривизна мембраны с первого взгляда. J Cell Sci (2015) 128(6):1065–70. doi: 10.1242/jcs.114454
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
130. Suetsugu S, Kurisu S, Takenawa T. Динамическое формирование клеточных мембран фосфолипидами и мембрано-деформирующими белками. Physiol Rev (2014) 94(4):1219–48. doi: 10.1152/physrev.00040.2013
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
131. Палмер А.Ф., Вингерт П., Никелс Дж. Атомно-силовая микроскопия и светорассеяние небольших однослойных актинсодержащих липосом. Biophys J (2003) 85(2):1233–47. doi: 10.1016/S0006-3495(03)74559-7
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
132.
Островски П.П., Фриман С.А., Фэрн Г., Гринштейн С. Динамические подосомоподобные структуры в зарождающихся фагосомах координируются фосфоинозитидами. Dev Cell (2019) 50(4):397–410.e3. doi: 10.1016/j.devcel.2019.05.028PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
133. Marion S, Mazzolini J, Herit F, Bourdoncle P, Kambou-Pene N, Hailfinger S, et al. Сигнальный белок NF-κB Bcl10 регулирует динамику актина, контролируя AP1 и везикулы, несущие OCRL. Dev Cell (2012) 23(5):954–67. doi: 10.1016/j.devcel.2012.09.021
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
134. De Leon-Rodriguez CM, Fu MS, Corbali MO, Cordero RJB, Casadevall A. Капсула Cryptococcus neoformans модулирует рН фагосом за счет своих кислотно-щелочных свойств. mSphere (2018) 3(5):e00437-18. doi: 10.1128/mSphere.00437-18
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
135. Johnston SA, May RC.
Грибковый патоген человека Cryptococcus neoformans ускользает от макрофагов с помощью механизма опорожнения фагосомы, который ингибируется опосредованной комплексом Arp2/3 полимеризацией актина. PloS Pathog (2010) 6(8):e1001041. doi: 10.1371/journal.ppat.1001041PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
136. Marion S, Hoffmann E, Holzer D, Le Clainche C, Martin M, Sachse M, et al. Эзрин способствует сборке актина на мембране фагосомы и регулирует слияние фаго-лизосом. Traffic (2011) 12(4):421–37. doi: 10.1111/j.1600-0854.2011.01158.x
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
137. Kasper L, Seider K, Hube B. Внутриклеточное выживание Candida glabrata в макрофагах: уклонение от иммунитета и персистенция. FEMS Yeast Res (2015) 15(5):fov042. doi: 10.1093/femsyr/fov042
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
138. Джин С., Кигер А.А.
Краткий обзор классов фосфоинозитид-3-киназ. J Cell Sci (2014) 127 (Pt 5): 923–8. doi: 10.1242/jcs.0PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
139. Бинкер М.Г., Чжао Д.Ю., Панг С.Дж., Харрисон Р.Э. Цитоплазматический линкерный белок-170 усиливает распространение и фагоцитоз в активированных макрофагах за счет стабилизации микротрубочек. J Immunol (2007) 179(6):3780–91. doi: 10.4049/jimmunol.179.6.3780
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
140. Гилберти Р.М., Кнехт Д.А. Макрофаги фагоцитируют неопсонизированные частицы кремнезема, используя уникальный путь, зависящий от микротрубочек. Mol Biol Cell (2015) 26(3):518–29. doi: 10.1091/mbc.E14-08-1301
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
141. Eng EW, Bettio A, Ibrahim J, Harrison RE. Переориентация MTOC происходит во время FcgammaR-опосредованного фагоцитоза в макрофагах.
Mol Biol Cell (2007) 18(7):2389–99. doi: 10.1091/mbc.e06-12-1128PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
142. Bonifacino JS, Neefjes J. Перемещение и позиционирование эндолизосомной системы. Curr Opin Cell Biol (2017) 47:1–8. doi: 10.1016/j.ceb.2017.01.008
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
143. Блокер А., Гриффитс Г., Оливо Дж. К., Хайман А. А., Северин Ф. Ф. Роль динамики микротрубочек в движении фагосом. J Cell Sci (1998) 111(3):303.
Резюме PubMed | Google Scholar
144. Blocker A, Severin FF, Burkhardt JK, Bingham JB, Yu H, Olivo J-C, et al. Молекулярные требования для двунаправленного движения фагосом вдоль микротрубочек. J Cell Biol (1997) 137(1):113–29. doi: 10.1083/jcb.137.1.113
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
145. Hart PD, Young MR, Jordan MM, Perkins WJ, Geisow MJ. Химические ингибиторы слияния фагосомы и лизосомы в культивируемых макрофагах также ингибируют скачкообразные движения лизосом.
Комбинированное микроскопическое и компьютерное исследование. J Exp Med (1983) 158(2):477–92. doi: 10.1084/jem.158.2.477PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
146. Sanchez L, Patton P, Anthony SM, Yi Y, Yu Y. Отслеживание вращения отдельных частиц во время поглощения макрофагами. Мягкая материя (2015) 11 (26): 5346–52. doi: 10.1039/C5SM00893J
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
147. Huotari J, Helenius A. Созревание эндосом. EMBO J (2011) 30(17):3481–500. doi: 10.1038/emboj.2011.286
Резюме PubMed | Полный текст перекрестной ссылки | Google Scholar
148. Fairn GD, Grinstein S. Как зарождающиеся фагосомы созревают, чтобы стать фаголизосомами. Trends Immunol (2012) 33(8):397–405. doi: 10.1016/j.it.2012.03.003
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
149. Dingjan I, Paardekooper LM, Verboogen DRJ, von Mollard GF, Ter Beest M, van den Bogaart G.
Опосредованное VAMP8 рекрутирование NOX2 в эндосомы необходимо для высвобождения антигена. Eur J Cell Biol (2017) 96(7):705–14. doi: 10.1016/j.ejcb.2017.06.007PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
150. Dingjan I, Linders PT. Окисленный фагосомальный комплекс NOX2 пополняется за счет лизосом. J Cell Sci (2017) 130(7):1285–98. doi: 10.1242/jcs.196931
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
151. Вулкано М., Дуси С., Лиссандрини Д., Бадолато Р., Мацци П., Рибольди Э. и др. Опосредованная рецептором Toll регуляция НАДФН-оксидазы в дендритных клетках человека. J Immunol (2004) 173(9):5749–56. doi: 10.4049/jimmunol.173.9.5749
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
152. Турк В., Стока В., Васильева О., Ренко М., Сун Т., Турк Б. и др. Цистеиновые катепсины: от структуры, функции и регуляции к новым рубежам. Biochim Biophys Acta (2012) 1824(1):68–88.
doi: 10.1016/j.bbapap.2011.10.002PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
153. Lee WL, Mason D, Schreiber AD, Grinstein S. Количественный анализ ремоделирования мембраны фагоцитарной чашки. Mol Biol Cell (2007) 18(8):2883–92. doi: 10.1091/mbc.e06-05-0450
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
154. Камен Л.А., Левинсон Дж., Суонсон Дж.А. Дифференциальная ассоциация фосфатидилинозитол-3-киназы, SHIP-1 и PTEN с формированием фагосом. Mol Biol Cell (2007) 18(7):2463–72. doi: 10.1091/mbc.e07-01-0061
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
155. Maxson ME, Naj X, O’Meara TR, Plumb JD, Cowen LE, Grinstein S. Диффузионный барьер на основе интегрина разделяет мембранные домены, обеспечивая образование микробостатических фрустрированных фагосом. eLife (2018) 7:e34798. doi: 10.7554/eLife.34798
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
156.
Kassas N, Tanguy E, Thahouly T, Fouillen L, Heintz D, Chasserot-Golaz S, et al. Сравнительная характеристика сенсоров фосфатидной кислоты и их локализация при фрустрированном фагоцитозе. J Biol Chem (2017) 292(10):4266–79. doi: 10.1074/jbc.M116.742346PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
157. Кэннон К.С., Вудс Б.Л., Гладфельтер А.С. Нерешенная проблема того, как клетки воспринимают кривизну микронного масштаба. Trends Biochem Sci (2017) 42(12):961–76. doi: 10.1016/j.tibs.2017.10.001
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
158. Sun B, Wang X, Ji Z, Li R, Xia T. Активация воспаления NLRP3, вызванная сконструированными наноматериалами. Малый (2013) 9 (9-10): 1595–607. doi: 10.1002/smll.201201962
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
159. Саян М., Моссман Б.Т. Инфламмасома NLRP3 при воспалении и заболеваниях легких, связанных с патогенными частицами и волокнами.
Часть Fibre Toxicol (2016) 13(1):51. doi: 10.1186/s12989-016-0162-4PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
160. Desai J, Foresto-Neto O, Honarpisheh M, Steiger S, Nakazawa D, Popper B, et al. Частицы разного размера и формы вызывают некроптоз нейтрофилов с последующим высвобождением нейтрофильного внеклеточного хроматина, подобного ловушке. Научный представитель (2017) 7(1):15003. doi: 10.1038/s41598-017-15106-0
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
161. Лебре Ф., Шридхаран Р., Сокинс М.Дж., Келли Д.Дж., О’Брайен Ф.Дж., Лавель Е.К. Форма и размер частиц гидроксиапатита определяют воспалительные реакции после имплантации. Научный представитель (2017) 7(1):2922. doi: 10.1038/s41598-017-03086-0
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
162. Rabolli V, Lison D, Huaux F. Сложный каскад клеточных событий, регулирующих активацию воспаления и процессинг IL-1beta в ответ на вдыхаемые частицы.
Часть Fibre Toxicol (2016) 13(1):40. doi: 10.1186/s12989-016-0150-8PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
163. Caicedo MS, Samelko L, McAllister K, Jacobs JJ, Hallab NJ, et al. Увеличение как размера частиц сплава CoCrMo, так и неровностей поверхности вызывает повышенную активацию воспаления макрофагов in vitro потенциально за счет механизмов дестабилизации лизосом. J Orthop Res (2013) 31(10):1633–42. doi: 10.1002/jor.22411
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
164. Wang J, Chen HJ, Hang T, Yu Y, Liu G, He G, et al. Физическая активация врожденного иммунитета шиповидными частицами. Nat Nanotechnol (2018) 13(11):1078–86. doi: 10.1038/s41565-018-0274-0
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
165. Chen L, Liu J, Zhang Y, Zhang G, Kang Y, Chen A, et al. Токсичность наночастиц кремнезема для иммунной системы. Наномед (Лондон) (2018) 13 (15): 1939–62.
doi: 10.2217/nnm-2018-0076Полный текст CrossRef | Академия Google
166. Rabolli V, Lison D, Huaux F. Сложный каскад клеточных событий, регулирующих активацию воспаления и процессинг IL-1β в ответ на вдыхаемые частицы. Часть Fibre Toxicol (2016) 13(1):40–0. doi: 10.1186/s12989-016-0150-8
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
167. де Риверо Ваккари Дж.П., Дитрих В.Д., Кин Р.В. Активация и регуляция клеточных инфламмасом: пробелы в наших знаниях о поражении центральной нервной системы. J Cereb Blood Flow Metab (2014) 34(3):369–75. doi: 10.1038/jcbfm.2013.227
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
168. Джонсон Б.М., Джанкинс Р.Д., Тинг Дж.П. Всплеск воспаления. Нац. Нанотехнология (2018) 13(11):980–1. doi: 10.1038/s41565-018-0292-y
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
169. Кусака Т., Накаяма М., Накамура К.
, Ишимия М., Фурусава Э., Огасавара К. Влияние размера частиц кремнезема на воспалительные реакции макрофагов. PloS One (2014) 9(3):e. doi: 10.1371/journal.pone.00
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
170. Palomäki J, Välimäki E, Sund J, Vippola M, Clausen PA, Jensen KA, et al. Длинные игольчатые углеродные нанотрубки и асбест активируют инфламмасому NLRP3 с помощью сходного механизма. ACS Nano (2011) 5(9):6861–70. doi: 10.1021/nn200595c
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
171. Hornung V, Bauernfeind F, Halle A, Samstad EO, Kono H, Rock KL, et al. Кристаллы кремнезема и соли алюминия активируют инфламмасому NALP3 посредством дестабилизации фагосом. Nat Immunol (2008) 9(8):847–56. doi: 10.1038/ni.1631
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
172. O’Meara TR, Duah K, Guo CX, Maxson ME, Gaudet RG, Koselny K, et al. Высокопроизводительный скрининг идентифицирует гены, необходимые для индукции Candida albicans пироптоза макрофагов.
mBio (2018) 9(4):e01581-18. doi: 10.1128/mBio.01581-18PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
173. Mammadov R, Cinar G, Gunduz N, Goktas M, Kayhan H, Tohumeken S, et al. Вирусоподобные наноструктуры для настройки иммунного ответа. Научный представитель (2015) 5:16728. doi: 10.1038/srep16728
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
174. Vaine CA, Patel MK, Zhu J, Lee E, Finberg RW, Hayward RC, et al. Настройка активации врожденного иммунитета путем текстурирования поверхности полимерных микрочастиц: роль формы в активации воспаления. J Immunol (2013) 190(7):3525–32. doi: 10.4049/jimmunol.1200492
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
175. Nadra I, Boccaccini AR, Philippidis P, Whelan LC, McCarthy GM, Haskard DO, et al. Влияние размера частиц на индуцированную кристаллами гидроксиапатита секрецию фактора некроза опухоли альфа макрофагами.
Атеросклероз (2008) 196(1):98–105. doi: 10.1016/j.atherosclerosis.2007.02.005PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
176. Silva AL, Soema PC, Slutter B, Ossendorp F, Jiskoot W. Системы доставки частиц PLGA для субъединичных вакцин: связь свойств частиц с иммуногенностью. Hum Vaccin Immunother (2016) 12(4):1056–69. doi: 10.1080/21645515.2015.1117714
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
177. Мант А., Чиннери Ф., Эллиотт Т., Уильямс А.П. На путь перекрестной презентации влияет размер частиц фагоцитированного антигена. Иммунология (2012) 136(2):163–75. doi: 10.1111/j.1365-2567.2012.03558.x
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
178. Брюэр Дж. М., Поллок К. Г., Тетли Л., Рассел Д. Г. Размер везикул влияет на перенос, процессинг и представление антигенов в липидных везикулах. J Immunol (2004) 173(10):6143–50. дои: 10.
4049/jиммунол.173.10.6143Резюме PubMed | Полный текст перекрестной ссылки | Google Scholar
179. Seydoux E, Rothen-Rutishauser B, Nita IM, Balog S, Gazdhar A, Stumbles PA, et al. Зависимое от размера накопление частиц в лизосомах модулирует функцию дендритных клеток посредством нарушения деградации антигена. Int J Nanomed (2014) 9:3885–902. doi: 10.2147/IJN.S64353
Полный текст CrossRef | Google Scholar
180. Шима Ф., Уто Т., Акаги Т., Баба М., Акаши М. Влияние размера амфифильных наночастиц поли(гамма-глутаминовой кислоты) на клеточное поглощение и созревание дендритных клеток in vivo. Acta Biomater (2013) 9(11):8894–901. doi: 10.1016/j.actbio.2013.06.010
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
181. Бенн Н., ван Дуйн Дж., Куйпер Дж., Джискот В., Слаттер Б. Управление иммунными реакциями: как размер, форма и жесткость влияют на иммуногенность вакцин в виде частиц. J Control Release (2016) 234:124–34.
doi: 10.1016/j.jconrel.2016.05.033PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
182. Рамирес-Ортис З.Г., Меанс Т.К. Роль дендритных клеток во врожденном распознавании патогенных грибов (A. fumigatus, C. neoformans и C. albicans). Вирулентность (2012) 3(7):635–46. doi: 10.4161/viru.22295
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
183. Mottram PL, Leong D, Crimeen-Irwin B, Gloster S, Xiang SD, Meanger J, et al. Иммунитет типа 1 и 2 после вакцинации зависит от размера наночастиц: состав модельной вакцины против респираторно-синцитиального вируса. Мол Фарм (2007) 4(1):73–84. doi: 10.1021/mp060096p
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
184. Fifis T, Gamvrellis A, Crimeen-Irwin B, Pietersz GA, Li J, Mottram PL, et al. Иммуногенность, зависящая от размера: терапевтические и защитные свойства нановакцин против опухолей. J Immunol (2004) 173(5):3148–54.
doi: 10.4049/jimmunol.173.5.3148PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
185. Канчан В., Панда А.К. Взаимодействия полилактидных частиц, нагруженных антигеном, с макрофагами и их корреляция с иммунным ответом. Биоматериалы (2007) 28(35):5344–57. doi: 10.1016/j.biomaterials.2007.08.015
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
186. Li X, Sloat BR, Yanasarn N, Cui Z. Взаимосвязь между размером наночастиц и их адъювантной активностью: данные исследования с улучшенным экспериментальным планом. Eur J Pharm Biopharm (2011) 78(1):107–16. doi: 10.1016/j.ejpb.2010.12.017
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
187. Zhao W, Hanson L, Lou HY, Akamatsu M, Chowdary PD, Santoro F, et al. Наномасштабные манипуляции с кривизной мембраны для исследования эндоцитоза в живых клетках. Нанотехнологии (2017) 12(8):750–6. doi: 10.1038/nnano.2017.98
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
188.
Lou HY, Zhao W, Li X, Duan L, Powers A, Akamatsu M, et al. Кривизна мембраны лежит в основе реорганизации актина в ответ на наноразмерную топографию поверхности. Proc Natl Acad Sci USA (2019) 116(46):23143–51. doi: 10.1073/pnas.16116PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
189. De Martino S, Zhang W, Klausen L, Lou HY, Li X, Alfonso FS, et al. Динамическое управление кривизной клеточной мембраны путем изменения формы азополимера под действием света. Nano Lett (2020) 20 (1): 577–84. doi: 10.1021/acs.nanolett.9b04307
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
190. Li X, Matino L, Zhang W, Klausen L, McGuire AF, Lubrano C, et al. Платформа наноструктур для манипулирования кривизной мембраны живых клеток. Nat Protoc (2019) 14(6):1772–802. doi: 10.1038/s41596-019-0161-7
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
191. Лу Х.И., Чжао В.
, Цзэн Ю., Цуй Б. Роль кривизны мембраны во внутриклеточной передаче сигналов, индуцированной наноразмерной топографией. Acc Chem Res (2018) 51(5):1046–53. doi: 10.1021/acs.accounts.7b00594PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
192. Сингх А.В., Патил Р., Томбре Д.К., Гейд В.Н. Микронанопаттернирование как инструмент для изучения роли физико-химических свойств во взаимодействии клеток с поверхностью. J BioMed Mater Res A (2013) 101(10):3019–32. doi: 10.1002/jbm.a.34586
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
193. Йим Э.К., Леонг К.В. Значение синтетических наноструктур в определении клеточного ответа. Наномедицина (2005) 1(1):10–21. doi: 10.1016/j.nano.2004.11.008
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
194. Palacci J, Sacanna S, Steinberg AP, Pine DJ, Chaikin PM. Живые кристаллы активируемых светом коллоидных серферов. Наука (2013) 339(6122):936–40.
doi: 10.1126/science.1230020PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
195. Юссеф М., Хюкель Т., Йи Г.Р., Саканна С. Коллоиды, изменяющие форму, посредством стимулированного осушения. Nat Commun (2016) 7:12216. doi: 10.1038/ncomms12216
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
196. Verboogen DRJ, Baranov MV, Ter Beest M, van den Bogaart G. Визуализация внутриклеточного трафика SNARE с помощью микроскопии флуоресцентной визуализации. J Vis Exp (2017) (130): e56745. doi: 10.3791/56745
CrossRef Полный текст | Google Scholar
197. Verboogen DRJ, Gonzalez Mancha N, Ter Beest M, van den Bogaart G. Микроскопия флуоресцентной визуализации в течение всей жизни показывает перенаправление SNARE-трафика, вызывающего активацию дендритных клеток. eLife (2017) 6:e23525. doi: 10.7554/eLife.23525
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
198.
Nelsen E, Hobson CM, Kern ME, Hsiao JP, O’Brien III ET, Watanabe T, et al. Комбинированная система атомно-силового микроскопа и объемного светового листа для корреляционных силовых и флуоресцентных механобиологических исследований. Научный представитель (2020) 10(1):8133. doi: 10.1038/s41598-020-65205-8PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
199. Озкан А., Маклеод Э. Безлинзовые изображения и датчики. Annu Rev BioMed Eng (2016) 18:77–102. doi: 10.1146/annurev-bioeng-0
-010849PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
200. Noireaux V, Libchaber A. Везикулярный биореактор как шаг к сборке искусственных клеток. Proc Natl Acad Sci USA (2004) 101 (51): 17669–74. doi: 10.1073/pnas.0408236101
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
201. Тревихано-Контадор Н., Руэда С., Сарагоса О. Морфогенетические изменения грибов внутри хозяина-млекопитающего.
Semin Cell Dev Biol (2016) 57:100–9. doi: 10.1016/j.semcdb.2016.04.008PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
202. Симмхен Дж., Катури Дж., Успал В.Е., Попеску М.Н., Тасинкевич М., Санчес С. Топографические пути направляют химических микропловцов. Nat Commun (2016) 7:10598. doi: 10.1038/ncomms10598
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
203. Санчес С., Солер Л., Катури Дж. Химические микро- и наномоторы. Angew Chem Int Ed Engl (2015) 54(5):1414–44. doi: 10.1002/anie.201406096
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
204. Chen P, Xu Z, Zhu G, Dai X, Yan LT. Клеточное поглощение активных частиц. Phys Rev Lett (2020) 124(19)):198102. doi: 10.1103/PhysRevLett.124.198102
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
205. Howse JR, Jones RA, Ryan AJ, Gough T, Vafabakhsh R, Golestanian R, et al.
Самодвижущиеся коллоидные частицы: от направленного движения к беспорядочному блужданию. Phys Rev Lett (2007) 99(4):048102. doi: 10.1103/PhysRevLett.99.048102PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
206. Гао В., Пей А., Ван Дж. Микромоторы с водяным приводом. ACS Nano (2012) 6(9):8432–8. doi: 10.1021/nn303309z
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
207. Wu Y, Wu Z, Lin X, He Q, Li J. Автономное движение управляемых собранных капсульных двигателей Janus. ACS Nano (2012) 6(12):10910–6. doi: 10.1021/nn304335x
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
208. Ван В., Чанг Т.Ю., Велегол Д., Маллук Т.Э. Понимание эффективности автономных нано- и микродвигателей. J Am Chem Soc (2013) 135(28):10557–65. doi: 10.1021/ja405135f
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
209. Ван С., Ву Н.
Выбор механизмов плавания коллоидных частиц: движение пузырьков по сравнению с самодиффузиофорезом. Ленгмюр (2014) 30(12):3477–86. doi: 10.1021/la500182fPubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
210. Бертье Л. Неравновесная стеклообразная динамика самодвижущихся жестких дисков. Phys Rev Lett (2014) 112(22):220602. doi: 10.1103/PhysRevLett.112.220602
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
211. Голестанян Р., Ливерпуль Т.Б., Айдари А. Движение молекулярной машины за счет асимметричного распределения продуктов реакции. Phys Rev Lett (2005) 94(22):220801. doi: 10.1103/PhysRevLett.94.220801
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
212. Palacci J, Sacanna S, Kim SH, Yi GR, Pine DJ, Chaikin PM. Светоактивируемые самодвижущиеся коллоиды. Philos Trans A Math Phys Eng Sci (2014) 372 (2029): 20130372. doi: 10.1098/rsta.2013.0372
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
213.
Paardekooper LM, Dingjan I, Linders PTA, Staal AHJ, Cristescu SM, Verberk W, et al. Дендритные клетки человека, полученные из моноцитов, продуцируют миллимолярные концентрации АФК в фагосомах в секунду. Фронт Иммунол (2019) 10:1216. doi: 10.3389/fimmu.2019.01216PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
214. Бедард К., Краузе К.Х. Семейство NOX НАДФН-оксидаз, генерирующих АФК: физиология и патофизиология. Physiol Rev (2007) 87(1):245–313. doi: 10.1152/physrev.00044.2005
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
215. Мандал П., Патил Г., Какоти Х., Гош А. Магнитно-активная материя на основе спирального движения. Acc Chem Res (2018) 51(11):2689–98. doi: 10.1021/acs.accounts.8b00315
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
216. Francia V, Montizaan D. Взаимодействие на клеточной мембране и пути интернализации наноразмерных материалов для наномедицины.
Beilstein J Nanotechnol (2020) 11:338–53. doi: 10.3762/bjnano.11.25PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
217. Getts DR, Shea LD, Miller SD, King NJ. Использование наночастиц для иммуномодуляции. Trends Immunol (2015) 36(7):419–27. doi: 10.1016/j.it.2015.05.007
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
Цирконий — информация об элементе, свойства и применение
Перейти к основному содержанию
У вас не включен JavaScript. Пожалуйста, включите JavaScript, чтобы получить доступ ко всем функциям сайта.
Перейти к ниобию >
Группа 4 Температура плавления 1854°С, 3369°F, 2127 К Период 5 Температура кипения 4406°С, 7963°F, 4679 К Блок г Плотность (г см −3 ) 6,52 Атомный номер 40 Относительная атомная масса 91,224 Состояние при 20°С Твердый Ключевые изотопы 90 Zr, 92 Zr, 94 Zr Электронная конфигурация [Кр] 4d 2 5s 2 Номер КАС 7440-67-7 ChemSpider ID 22431 ChemSpider — бесплатная база данных химической структуры. Древние египтяне использовали драгоценные камни циркона в украшениях. Для древних египтян жук-скарабей (представленный здесь) был символом возрождения и созидания, передающим идеи трансформации, обновления и воскресения.
Твердый металл серебристого цвета, очень устойчивый к коррозии.
Цирконий не поглощает нейтроны, что делает его идеальным материалом для использования на атомных электростанциях. Таким образом используется более 90% циркония. Ядерные реакторы могут иметь более 100 000 метров труб из циркониевого сплава. Вместе с ниобием цирконий обладает сверхпроводящими свойствами при низких температурах и используется для изготовления сверхпроводящих магнитов.
Металлический цирконий защищен тонким слоем оксида, что делает его исключительно устойчивым к коррозии кислотами, щелочами и морской водой. По этой причине он широко используется в химической промышленности.
Оксид циркония(IV) используется в сверхпрочной керамике. Он используется для изготовления тиглей, выдерживающих тепловой удар, футеровки печей, литейного кирпича, абразивов, а также в стекольной и керамической промышленности. Он настолько прочен, что из него можно делать даже ножницы и ножи. Он также используется в косметике, антиперспирантах, пищевой упаковке и для изготовления микроволновых фильтров.
Циркон — природный полудрагоценный камень, встречающийся в различных цветах. Самые желательные имеют золотистый оттенок. Элемент был впервые обнаружен в этой форме, что привело к его названию. Кубический цирконий (оксид циркония) — синтетический драгоценный камень. Бесцветные камни при огранке напоминают бриллианты.
Циркон, смешанный с ванадием или празеодимом, дает синий и желтый пигменты для глазури керамики.
Цирконий не играет известной биологической роли. Обладает низкой токсичностью.
Цирконий встречается примерно в 30 видах минералов, основными из которых являются циркон и бадделеит.
Ежегодно добывается более 1,5 млн тонн циркона, в основном в Австралии и Южной Африке. Большая часть бадделеита добывается в Бразилии.Металлический цирконий получают в промышленных масштабах, сначала превращая циркон в хлорид циркония, а затем восстанавливая хлорид магнием.
Элементы и история периодической таблицы
Драгоценные камни, содержащие цирконий, в древние времена были известны как циркон. В 1789 году немецкий химик Мартин Клапрот проанализировал циркон и выделил цирконий в форме его «земного» циркония, который представляет собой оксид ZrO 2 .
Клапроту не удалось выделить сам чистый металл, и Хамфри Дэви также потерпел неудачу, когда попытался провести электролиз в 1808 году. Только в 1824 году элемент был выделен, когда шведский химик Йонс Берцелиус нагрел гексафтороцирконат калия (K 2 ZrF 6 ) с металлическим калием и получил некоторое количество циркония в виде черного порошка.
Полностью чистый цирконий был получен только в 1925 году голландскими химиками Антоном Эдуардом ван Аркелем и Яном Хендриком де Буром путем разложения тетраиодида циркония (ZrI 4 ).
В настоящее время металл производится в больших количествах путем нагревания тетрахлорида циркония (ZrCl 4 ) с магнием.Атомный радиус, несвязанный (Å) 2,23 Ковалентный радиус (Å) 1,64 Сродство к электрону (кДж моль −1 ) 41. 103Электроотрицательность
(шкала Полинга)1,33 Энергии ионизации
(кДж моль −1 )1 ст
640.074
2 и
1264
3 -й
2218.
24 -й
3313,31
5 -й
7752.404
Общие степени окисления 4 Изотопы Изотоп Атомная масса Естественное изобилие (%) Период полураспада Режим распада 90 Зр 89. 90551,45 — — 91 Зр 90.906 11.22 — — 92 Зр 91,905 17. 15— — 94 Зр 93,906 17.38 > 10 17 г β-β- 96 Зр 95,908 2,8 2,3 х 10 19 г β-β- > 1,7 х 18 лет β- Относительный риск поставок 5. 7Содержание земной коры (ppm) 132 Скорость переработки (%) Взаимозаменяемость Неизвестный Концентрация производства (%) 39 Распределение резерва (%) 40 Топ-3 производителя - 1) Австралия
- 2) ЮАР
- 3) Китай
Топ 3 держателя резерва - 1) Австралия
- 2) ЮАР
- 3) Украина
Политическая стабильность топ-производителя 74,5 Политическая стабильность главного держателя резерва 74,5 Удельная теплоемкость
(Дж кг -1 К -1 )278 Модуль Юнга (ГПа) Неизвестный Модуль сдвига (ГПа) Неизвестный Объемный модуль (ГПа) Неизвестный Давление пара Температура (К) 400 600 800 1000 1200 1400 1600 1800 2000 г. 2200 2400 Давление (Па) — — — — — 1,05
x 10 -106. 17
x 10 -88,68
x 10 -60,00045 0,011 0,155 Слушайте циркониевый подкаст Стенограмма:
(Promo)
Вы слушаете Химию в ее стихии, представленную вам Chemistry World , журнал Королевского химического общества.
(Конец промо)
Крис Смит
Привет и добро пожаловать в наш тур по необычным, захватывающим и смертоносным аспектам элементов, из которых состоит мир вокруг нас. Мы начинаем наше путешествие по Периодической таблице с химического вещества, которое иногда маскируется под алмаз, но в равной степени чувствует себя как дома в ядре ядерного реактора или даже на металлургическом заводе. Чтобы рассказать историю этого таинственного вещества, которое также известно как цирконий, химик и отмеченный наградами автор Джон Эмсли расскажет вам об этом.
Джон Эмсли
Цирконий. Носите его сверкающим на пальце или невидимым в кадре, он держит ключ к ядерной энергии, и у него имя, похожее на драгоценный камень. Это цирконий.
Название цирконий происходит от арабского слова zargun , обозначающего золотистый драгоценный камень, известный с библейских времен под названием циркон. Сегодня искусственные драгоценные камни изготавливаются из оксида циркония, известного как кубический цирконий, и они сверкают ярче, чем алмаз, хотя и не такие твердые.
Что отличает их от настоящего алмаза, так это их более высокая плотность 6,0 г см -3 по сравнению с алмазным 3,52.Цирконий широко распространен в звездах S-типа, в которых более тяжелые элементы образуются в результате захвата нейтронов. Следы есть и на Солнце. Было обнаружено, что горная порода, доставленная с Луны, имеет удивительно высокое содержание циркония. Цирконы здесь, на Земле, показали, что жизнь могла зародиться намного раньше, чем предполагалось. Они были обнаружены в Австралии в 2000 году, им было 4,4 миллиарда лет, а соотношение изотопов кислорода в них составляло O 16 /O 9.1739 18 показали, что они могли образоваться только тогда, когда на поверхности Земли была жидкая вода, а это произошло почти на 500 миллионов лет раньше, чем предполагалось ранее.
В Средние века бесцветные драгоценные камни циркона считались низшим видом алмаза, но это оказалось неверным, когда немецкий химик Мартин Клапрот (1743-1817) проанализировал один из них в 1789 году и обнаружил цирконий.
Клапрот не смог выделить сам металл. Это было достигнуто в 1824 году шведским химиком Йонсом Якобом Берцелиусом, но от него или его химических соединений было мало пользы, и поэтому он томился в течение столетия или более.Сегодня этот элемент широко используется в качестве циркона, оксида циркония и самого металла. Цирконий можно найти в керамике, литейном оборудовании, стекле, химикатах и металлических сплавах.
Песок циркониевый применяется для термостойкой футеровки печей, гигантских ковшей для расплавленного металла, изготовления литейных форм. Циркон, смешанный с ванадием или празеодимом, дает синий и желтый пигменты для полировки керамики и плитки.
Оксид циркония используется для изготовления термостойких тиглей, керамики и абразивов. Сделанный из него раскаленный тигель можно опустить в холодную воду, и он не треснет. Оксид циркония содержится в сверхпрочной керамике, которая прочнее и острее, чем даже закаленная сталь, и используется для изготовления ножей, ножниц и клюшек для гольфа.
Производство чистого оксида циркония составляет почти 25 000 тонн в год, и он также идет на различные химические вещества, которые в конечном итоге используются в качестве косметики, антиперспирантов, упаковки для пищевых продуктов и даже поддельных драгоценных камней. Бумажная и упаковочная промышленность считает, что соединения циркония являются хорошими поверхностными покрытиями, поскольку они обладают отличной водостойкостью и прочностью. Не менее важна их низкая токсичность.Металлический цирконий имеет окисленную поверхность, которая является одновременно твердой и невосприимчивой к химическому воздействию, что делает его идеальным не только для химических заводов, но и для имплантатов тела, таких как протезы тазобедренного сустава. Цирконий-алюминиевый сплав используется для производства велосипедных рам высшего класса, потому что он сочетает в себе прочность и легкость.
Металлический цирконий обладал некоторыми скрытыми активами, которые внезапно сделали его известным в конце 1940-х годов; было обнаружено, что это идеальный металл для внутренней части ядерных реакторов и атомных подводных лодок.
Он не подвергается коррозии при высоких температурах и не поглощает нейтроны с образованием радиоактивных изотопов. Даже сегодня атомная промышленность покупает почти весь производимый металл, а некоторые ядерные реакторы имеют более 100 километров циркониевых труб. Цирконий используется для изготовления оболочек тепловыделяющих элементов из оксида урана. В добытом цирконии содержится 1-3% гафния, очень похожего по химическому составу, и, хотя эти два элемента трудно разделить, это необходимо сделать для металла, используемого в атомной промышленности, поскольку гафний очень сильно поглощает нейтроны.Наконец, у нас есть два циркониевых материала с экстремальными свойствами, один из которых проявляет себя при очень низких температурах, а другой — при нагревании до высоких температур. Первый представляет собой сплав циркония-ниобия, который становится сверхпроводящим при температуре ниже 35 Кельвинов (- 238 o C), другими словами, он будет проводить электричество без потери энергии.
Второй — вольфрамат циркония (ZrW 2 O 8 ), который на самом деле сжимается при нагревании, по крайней мере, до достижения 700 o C при разложении на два оксида металла.Крис Смит
Джон Эмсли раскрывает секрет элемента номер 40, циркония. И вы можете узнать больше о любимых элементах Джона в серии, которую он написал для RSC’s Education in Chemistry, которая доступна онлайн по адресу rsc.org/education. В следующий раз на «Химии в ее стихии» жизнь — это газ с Марком Пеплоу.
Марк Пеплоу
Эти скромные цианобактерии даже не осознавали, что они делали, когда два с половиной миллиарда лет назад они начали создавать свои собственные запасы богатых энергией химических веществ, комбинируя воду и углекислый газ. Приведенные в действие солнечным светом, они провели следующие два миллиарда лет, терраформируя всю нашу планету с помощью отхода своего фотосинтеза, довольно токсичного газа, называемого кислородом.
Крис Смит
Так что присоединяйтесь к нам на следующей неделе, чтобы подышать свежим воздухом и узнать историю о кислороде. Я Крис Смит, спасибо за внимание, увидимся в следующий раз.
(Promo)
Химия в ее стихии представлена вам Королевским химическим обществом и произведена thenakedscientists.com. Дополнительную информацию и другие эпизоды химии в ее стихии можно найти на нашем веб-сайте chemistryworld.org/elements.
(Конец промо)
Нажмите здесь, чтобы посмотреть видео о цирконии
Learn Chemistry: ваш единственный путь к сотням бесплатных учебных ресурсов по химии.
Изображения и видео Visual Elements
© Murray Robertson 1998-2017.
Data
W.M. Haynes, ed., CRC Handbook of Chemistry and Physics , CRC Press/Taylor and Francis, Boca Raton, FL, 95th Edition, Internet Version 2015, по состоянию на декабрь 2014 г.Таблица 906 & Chemical Constants, Kaye & Laby Online, 16-е издание, 1995 г. Версия 1.0 (2005 г.), по состоянию на декабрь 2014 г.
Дж. С. Курси, Д. Дж. Шваб, Дж. Дж. Цай и Р. А. Драгосет, Атомные веса и изотопные композиции (версия 4.1) , 2015 г., Национальный институт стандартов и технологий, Гейтерсберг, Мэриленд, по состоянию на ноябрь 2016 г.
TL Cottrell, The Strengths of Chemical Bonds , Butterworth, London, 1954.
Использование и свойства
John Emsley, Строительные блоки природы: руководство по элементам от А до Я , Oxford University Press, New York, 2nd, New York, 2nd.
 В отличие от актиновых филаментов и микротрубочек промежуточные филаменты не принимают непосредственного участия в клеточных движениях. Вместо этого они, по-видимому, играют в основном структурную роль, обеспечивая механическую прочность клеток и тканей.
В отличие от актиновых филаментов и микротрубочек промежуточные филаменты не принимают непосредственного участия в клеточных движениях. Вместо этого они, по-видимому, играют в основном структурную роль, обеспечивая механическую прочность клеток и тканей.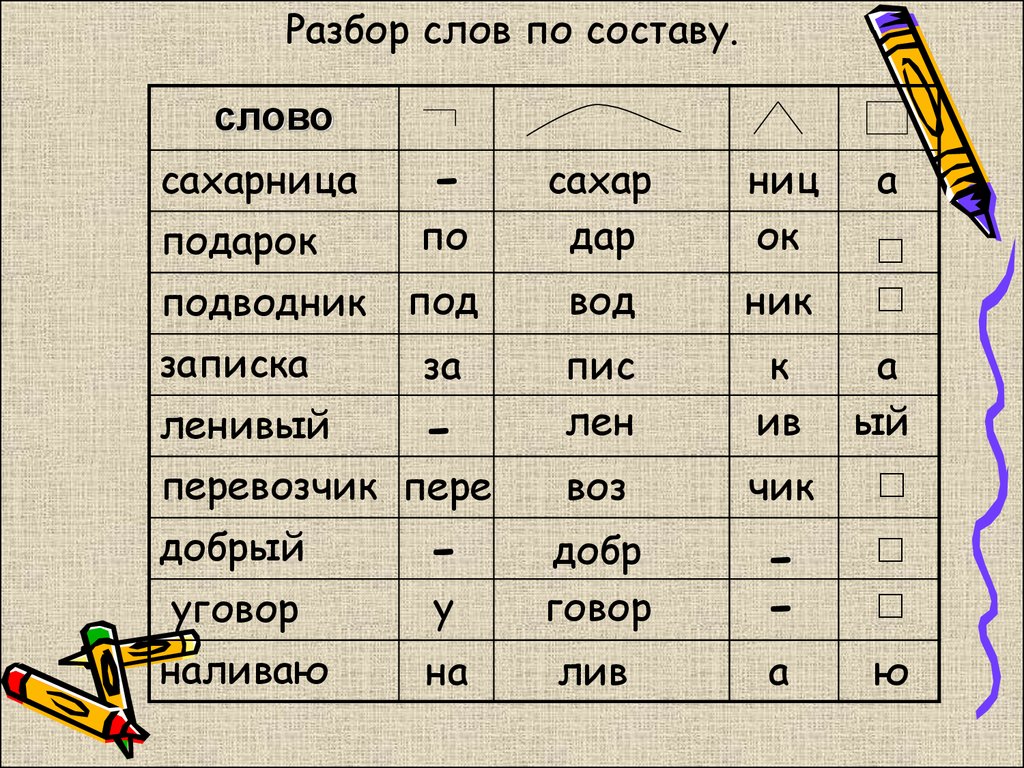 Некоторые кератины типа I и II (называемые твердые кератины ) используются для изготовления таких структур, как волосы, ногти и рога. Другие кератины I и II типов ( мягкие кератины ) в изобилии присутствуют в цитоплазме эпителиальных клеток, при этом разные кератины экспрессируются в различных типах дифференцированных клеток.
Некоторые кератины типа I и II (называемые твердые кератины ) используются для изготовления таких структур, как волосы, ногти и рога. Другие кератины I и II типов ( мягкие кератины ) в изобилии присутствуют в цитоплазме эпителиальных клеток, при этом разные кератины экспрессируются в различных типах дифференцированных клеток. ). Эти белки образуют основные промежуточные филаменты многих типов зрелых нейронов. Их особенно много в аксонах двигательных нейронов, и считается, что они играют решающую роль в поддержании этих длинных тонких отростков, длина которых может достигать более метра. Другой белок типа IV (α-интернексин) экспрессируется на более ранней стадии развития нейронов, до экспрессии белков нейрофиламентов. Единственный белок промежуточных филаментов VI типа (нестин) экспрессируется еще раньше в процессе развития нейронов, в стволовых клетках центральной нервной системы.
). Эти белки образуют основные промежуточные филаменты многих типов зрелых нейронов. Их особенно много в аксонах двигательных нейронов, и считается, что они играют решающую роль в поддержании этих длинных тонких отростков, длина которых может достигать более метра. Другой белок типа IV (α-интернексин) экспрессируется на более ранней стадии развития нейронов, до экспрессии белков нейрофиламентов. Единственный белок промежуточных филаментов VI типа (нестин) экспрессируется еще раньше в процессе развития нейронов, в стволовых клетках центральной нервной системы. Все белки промежуточных филаментов имеют центральный α-спиральный стержневой домен примерно из 310 аминокислот (350 аминокислот в ядерных ламинах). Этот центральный стержневой домен окружен амино- и карбоксиконцевыми доменами, которые варьируют среди различных белков промежуточных филаментов по размеру, последовательности и вторичной структуре. Как обсуждается далее, α-спиральный стержневой домен играет центральную роль в сборке филаментов, в то время как вариабельные головные и хвостовые домены, по-видимому, определяют специфические функции различных белков промежуточных филаментов.
Все белки промежуточных филаментов имеют центральный α-спиральный стержневой домен примерно из 310 аминокислот (350 аминокислот в ядерных ламинах). Этот центральный стержневой домен окружен амино- и карбоксиконцевыми доменами, которые варьируют среди различных белков промежуточных филаментов по размеру, последовательности и вторичной структуре. Как обсуждается далее, α-спиральный стержневой домен играет центральную роль в сборке филаментов, в то время как вариабельные головные и хвостовые домены, по-видимому, определяют специфические функции различных белков промежуточных филаментов. Затем димеры объединяются в шахматном порядке антипараллельно с образованием тетрамеров, которые могут собираться конец к концу, образуя протофиламенты. Последний промежуточный филамент содержит примерно восемь протофиламентов, намотанных друг вокруг друга в виде веревкообразной структуры. Поскольку они собраны из антипараллельных тетрамеров, оба конца промежуточных филаментов эквивалентны. Следовательно, в отличие от актиновых филаментов и микротрубочек промежуточные филаменты неполярны; у них нет отчетливых плюсовых и минусовых концов.
Затем димеры объединяются в шахматном порядке антипараллельно с образованием тетрамеров, которые могут собираться конец к концу, образуя протофиламенты. Последний промежуточный филамент содержит примерно восемь протофиламентов, намотанных друг вокруг друга в виде веревкообразной структуры. Поскольку они собраны из антипараллельных тетрамеров, оба конца промежуточных филаментов эквивалентны. Следовательно, в отличие от актиновых филаментов и микротрубочек промежуточные филаменты неполярны; у них нет отчетливых плюсовых и минусовых концов. Напротив, белки типа III могут собираться в филаменты, содержащие только один полипептид (например, виментин) или состоящие из двух разных белков типа III (например, виментин плюс десмин). Однако белки типа III не образуют сополимеров с кератинами. Среди белков типа IV α-интернексин может сам собираться в филаменты, тогда как три белка нейрофиламентов сополимеризуются с образованием гетерополимеров.
Напротив, белки типа III могут собираться в филаменты, содержащие только один полипептид (например, виментин) или состоящие из двух разных белков типа III (например, виментин плюс десмин). Однако белки типа III не образуют сополимеров с кератинами. Среди белков типа IV α-интернексин может сам собираться в филаменты, тогда как три белка нейрофиламентов сополимеризуются с образованием гетерополимеров.
 ..)
..)