Глицин — описание ингредиента, инструкция по применению, показания и противопоказания
16 Апреля 2020
2 Июня 2020
3 минуты
16086
ProWellness
Оглавление
- Описание глицина
- Состав и форма выпуска глицина
- Фармакологические свойства
- Действие на организм
- Противопоказания и побочные эффекты
- Способы применения и дозировки
Отказ от ответсвенности
Обращаем ваше внимание, что вся информация, размещённая на сайте
Prowellness предоставлена исключительно в ознакомительных целях и не является персональной программой, прямой рекомендацией к действию или врачебными советами. Не используйте данные материалы для диагностики, лечения или проведения любых медицинских манипуляций. Перед применением любой методики или употреблением любого продукта проконсультируйтесь с врачом. Данный сайт не является специализированным медицинским порталом и не заменяет профессиональной консультации специалиста.
Описание глицина
Глицин – это нейромедиаторная аминокислота, которая используется в медицине как ноотропное лекарственное средство. Синоним – аминоуксусная кислота. Она присутствует в составе многочисленных белков. Из нее в клетках живых организмов синтезируются производные пурина.
Препараты глицина используются в терапии психических и невролгических заболеваний. В головном и спинном мозге есть глициновые рецепторы. Аминокислота прикрепляется к ним и оказывает «замедляющее» воздействие на нейроны, сокращает выработку из них «возбуждающих» аминокислот и усиливает выделение гамма-аминомасляной кислоты – главного тормозного нейромедиатора. Глицин – это природный антидепрессант и стабилизатор нервной системы.
Состав и форма выпуска глицина
Глицин выпускается в форме таблеток белого цвета. В состав одного драже входит 100 мг активного вещества, 1 мг стеарата магния и 1 мг метилцеллюлозы.
В состав одного драже входит 100 мг активного вещества, 1 мг стеарата магния и 1 мг метилцеллюлозы.
Фармакологические свойства
Аминоуксусная кислота регулирует обменные процессы, которые активируют и приостанавливают защитное торможение в ЦНС. Препарат помогает поддерживать психическое и эмоциональное состояние в норме. Он снижает напряжение, тревожность и повышает интеллектуальные способности. Помимо этого, глицин:
- повышает настроение;
- облегчает засыпание;
- улучшает качество сна;
- снижает воздействие угнетающих ЦНС токсинов;
- снимает стресс;
- нормализует самочувствие;
- успокаивает.
Действие на организм
Глицин нормализует деятельность мозга и восстанавливает нормальное функционирование нервной системы. Его можно принимать при наличии следующих состояний и болезней:
- стрессовые ситуации;
- ишемический инсульт;
- неврозы и повышенная возбудимость;
- вегетососудистая дистония;
- энцефалопатия;
- черепно-мозговые травмы.


Противопоказания и побочные эффекты
Относительными противопоказаниями к приему глицина являются беременность, грудное вскармливание. Воздействие препарата на плод и новорожденного ребенка, находящегося на грудном вскармливании, пока не изучено, поэтому лучше отказаться от его применения.
Внимание! При гипотонии и одновременном приеме глицина нужно регулярно проверять артериальное давление. Если будут выявлены нарушения, следует скорректировать дозировку или заменить препарат на альтернативный.
Способы применения и дозировки
Глицин принимают сублингвально: кладут под язык и ждут полного растворения таблетки. Дозы и частота приема зависят от возраста человека, диагноза и клинической картины. Взрослые обычно употребляют по 1-2 таблетке в день на протяжении 30–90 суток. Затем делается как минимум трехмесячный перерыв, после чего можно возобновлять терапию. Детям препарат разрешен с 3 лет. Суточная доза не должна превышать 50% таблетки. Пьют это количество средства трижды в день не дольше 2 недель. По достижении ребенком 7 лет можно повысить дозу до целой таблетки или даже двух.
Суточная доза не должна превышать 50% таблетки. Пьют это количество средства трижды в день не дольше 2 недель. По достижении ребенком 7 лет можно повысить дозу до целой таблетки или даже двух.
Внимание! Прием глицина во время беременности или лактации нужно осуществлять под контролем врача.
Отказ от ответсвенности
Обращаем ваше внимание, что вся информация, размещённая на сайте
Prowellness предоставлена исключительно в ознакомительных целях и не является персональной программой, прямой рекомендацией к действию или врачебными советами. Не используйте данные материалы для диагностики, лечения или проведения любых медицинских манипуляций. Перед применением любой методики или употреблением любого продукта проконсультируйтесь с врачом. Данный сайт не является специализированным медицинским порталом и не заменяет профессиональной консультации специалиста. Владелец Сайта не несет никакой ответственности ни перед какой стороной, понесший косвенный или прямой ущерб в результате неправильного использования материалов, размещенных на данном ресурсе.
Автокаталитический цикл в патогенезе сахарного диабета: биохимические и патофизиологические аспекты метаболической терапии с помощью натуральных аминокислот на примере глицина | Нестеров
Сахарный диабет (СД) – заболевание, характеризующееся нарушением усвоения глюкозы клетками и повышением ее уровня в крови. Выделяют две основные формы СД – диабет первого и второго типов (СД1 и СД2). При СД1 происходит полная гибель β-клеток поджелудочной железы, синтезирующих инсулин, обусловленная формированием аутоиммунной реакции [1]. При СД2 наблюдается потеря чувствительности периферических тканей к инсулину (инсулинорезистентность), а также происходит нарушение функции митохондрий [2], часто нарушается секреция инсулина и глюкагона в ответ на глюкозу [3]. Проблема лечения СД стоит крайне остро, в России этим заболеванием страдают более 4 млн человек [4]. Общими проявлениями диабета обоих типов являются повышенная концентрация глюкозы и холестерина в крови, повышенное гликирование белков и липидов, воспалительные процессы. Основные осложнения СД – микро-и макрососудистые, приводящие к повреждению сердца, глаз, почек, мозга, нижних конечнотей.
Основные осложнения СД – микро-и макрососудистые, приводящие к повреждению сердца, глаз, почек, мозга, нижних конечнотей.
В настоящей работе создана модель развития метаболических нарушений при СД. В основе модели лежат две группы параллельных процессов, которые активируются в ходе развития заболевания: каскад биохимических реакций, связанных с синтезом активных форм кислорода (АФК), и каскад патофизиологических процессов, постепенно приводящих к нарушению микроциркуляции крови и индукции ишемии в тканях больного. Основной результат настоящей работы составляет обнаруженный при анализе банка данных эффект взаимодействия двух указанных выше подсистем. При этом за счет положительных обратных связей возникает согласованное во времени последовательное их автокаталитическое усиление, оказывающее разрушающее воздействие на организм. Этот цикл возникает на втором этапе развития заболевания (рис. 1) и независим от сигнала инсулина и концентрации глюкозы в крови. Важно подчеркнуть, что модель целиком построена на экспериментальном материале и каждое звено отображенных на рис. 1 каскадов наблюдается в реальных экспериментах. Подробное описание всех элементов схемы и ссылки на соответствующие литературные источники приведены в табл. 1, которая также позволяет проследить биологическое действие аминокислоты глицин на каждой стадии развития патологии.
1 каскадов наблюдается в реальных экспериментах. Подробное описание всех элементов схемы и ссылки на соответствующие литературные источники приведены в табл. 1, которая также позволяет проследить биологическое действие аминокислоты глицин на каждой стадии развития патологии.
Рис. 1. Схема нарастания уровня активных форм кислорода и развития воспалительного процесса при сахарном диабете. Описание пронумерованных стадий со ссылками на литературу, а также описание действия глицина на каждой стадии дано в таблице 1. Автокаталитические циклы подробно описаны в основном тексте статьи. Сокращения: АФК – активные формы кислорода; RAGE – рецепторы конечных продуктов гликирования; NOX – NADPH-оксидазы; NF-κB – ядерный фактор каппа-Би; SOD – супероксиддисмутаза.
Поскольку в ряде клинических исследований низкий уровень глицина ассоциируется с нарушением углеводного обмена, в том числе с СД2 [5–7], то на втором этапе работы на базе разработанной модели проведено исследование возможности действия аминокислоты глицин в качестве средства, ослабляющего автокаталитический цикл развития диабета. Проведенный в настоящей работе анализ действия глицина показывает, что дефицит данной аминокислоты является существенным провоспалительным фактором, способствует манифестации болезни и усугубляет ее течение. Компенсация дефицита аминоуксусной кислоты, наоборот, обуславливает разрыв положительных обратных связей, что подтверждается рядом клинических исследований, в которых показана способность глицина нормализовать биохимические параметры крови больных СД, предотвращать и обращать вспять диабетические осложнения [8–12]. Биохимические причины возникновения дефицита глицина при СД также обсуждаются в работе.
Проведенный в настоящей работе анализ действия глицина показывает, что дефицит данной аминокислоты является существенным провоспалительным фактором, способствует манифестации болезни и усугубляет ее течение. Компенсация дефицита аминоуксусной кислоты, наоборот, обуславливает разрыв положительных обратных связей, что подтверждается рядом клинических исследований, в которых показана способность глицина нормализовать биохимические параметры крови больных СД, предотвращать и обращать вспять диабетические осложнения [8–12]. Биохимические причины возникновения дефицита глицина при СД также обсуждаются в работе.
Модель автокаталитического развития метаболических нарушений при диабете
Проведенные ранее экспериментальные исследования и анализ литературных данных позволили разработать схему (см. рис. 1), отображающую развитие метаболических нарушений при СД во времени. Как отмечалось во введении, в рамках разработанной модели развитие заболевания рассмотрено как взаимодействие и взаимоусиление двух групп параллельно протекающих процессов. Одна из групп имеет биохимическую природу и связана с нарастанием уровня АФК в клетках и тканях. Спусковым механизмом активации указанного каскада биохимических реакций является повышение уровня конечных продуктов гликирования (КПГ) при диабете. Вторая группа процессов имеет патофизиологическую природу и связана с нарушением микроциркуляции. Нарушение метаболизма в печени при СД создает атерогенный липидный профиль, что на молекулярном уровне нарушает адекватное функционирование эндотелия и со временем приводит к атеросклеротическому ремоделированию сосудов. Необходимо подчеркнуть, что общей основой указанных патологических изменений является инсулиновая недостаточность и/или инсулинорезистентность, однако уже на ранней стадии развития заболевания процесс приобретает автокаталитический характер. При этом возникают циклические системы реакций, практически не зависящие ни от инсулина, ни от концентрации глюкозы в крови. Данный феномен известен в литературе как «метаболическая память» [47].
Одна из групп имеет биохимическую природу и связана с нарастанием уровня АФК в клетках и тканях. Спусковым механизмом активации указанного каскада биохимических реакций является повышение уровня конечных продуктов гликирования (КПГ) при диабете. Вторая группа процессов имеет патофизиологическую природу и связана с нарушением микроциркуляции. Нарушение метаболизма в печени при СД создает атерогенный липидный профиль, что на молекулярном уровне нарушает адекватное функционирование эндотелия и со временем приводит к атеросклеротическому ремоделированию сосудов. Необходимо подчеркнуть, что общей основой указанных патологических изменений является инсулиновая недостаточность и/или инсулинорезистентность, однако уже на ранней стадии развития заболевания процесс приобретает автокаталитический характер. При этом возникают циклические системы реакций, практически не зависящие ни от инсулина, ни от концентрации глюкозы в крови. Данный феномен известен в литературе как «метаболическая память» [47]. Основные автокаталитические циклы усиления окислительного стресса будут подробнее описаны ниже.
Основные автокаталитические циклы усиления окислительного стресса будут подробнее описаны ниже.
Первичная индукция синтеза АФК при диабете: биохимический автокаталитический цикл АФК-зависимого усиления сигнала RAGE
КПГ являются мощными воспалительными факторами и формируются неферментативным путем в результате присоединения к белкам и липидам глюкозы или дикарбонильных интермедиатов. КПГ накапливаются в крови и внеклеточном матриксе и локально активируют рецептор RAGE, который присутствует в плазматических мембранах клеток эндотелия и гладкой мускулатуры сосудов, нейронов, некоторых типов лейкоцитов и эпителиальных клеток [16]. Одним из основных следствий активации сигнального каскада RAGE является усиление генерации АФК NADPH-оксидазами (NOX) [48] и выброс клетками воспалительных цитокинов, в первую очередь, интерлейкина-6 [49]. Первичная активация RAGE при диабете происходит из-за глюкозозависимого образования и накопления КПГ (звено 2 на рис. 1). При нарастании уровня АФК сигнал RAGE уже на начальной стадии развития диабета перестает зависеть от уровня глюкозы в крови и приобретает автокаталитическую природу, поскольку RAGE-индуцированный синтез АФК увеличивает транскрипцию RAGE [50] посредством активации транскрипционного фактора NF-κB [19]. Кроме того, повышение уровня АФК приводит к глюкозонезависимому образованию КПГ [51], тем самым увеличивая количество агонистов RAGE-рецептора и усиливая его сигнал. Наличие указанных обратных связей доказано экспериментально: при диабете наблюдается хроническая активация фактора транскрипции NF-κB [52] и существенно увеличивается количество белка RAGE-рецептора [47].
Кроме того, повышение уровня АФК приводит к глюкозонезависимому образованию КПГ [51], тем самым увеличивая количество агонистов RAGE-рецептора и усиливая его сигнал. Наличие указанных обратных связей доказано экспериментально: при диабете наблюдается хроническая активация фактора транскрипции NF-κB [52] и существенно увеличивается количество белка RAGE-рецептора [47].
Основной автокаталитический цикл взаимного усиления патофизиологических и АФК-зависимых биохимических процессов
Наряду с активацией АФК-зависимого каскада реакций ослабление инсулиновой сигнализации индуцирует параллельно нарушение функции печени (звено 4 на рис. 1), что приводит к повышению уровня холестерина в крови, формированию атеросклеротических отложений на стенках сосудов, нарушению циркуляции крови и индукции ишемии в тканях (звено 5 на рис. 1). Этот процесс резко усиливается описанными выше реакциями синтеза АФК, связанными с активацией RAGE (звено 3 на рис. 1). Сигнал этого рецептора индуцирует ряд патологических процессов: кальцификацию сосудов [18]; АФК-зависимые повреждения сосудов [25]; выброс мастоцитами (тучными клетками) гистамина [17]. Гистамин индуцирует расширение сосудов и увеличение их проницаемости, что приводит к падению давления крови в капиллярах и их закупорке эритроцитами. Выброс гистамина усугубляет нарушение микроциркуляции крови и ишемию тканей. В зону ишемического поражения происходит хемотаксис нейтрофилов, которые генерируют большие количества АФК, а также способствуют формированию тромбов в сосудах [26].
Гистамин индуцирует расширение сосудов и увеличение их проницаемости, что приводит к падению давления крови в капиллярах и их закупорке эритроцитами. Выброс гистамина усугубляет нарушение микроциркуляции крови и ишемию тканей. В зону ишемического поражения происходит хемотаксис нейтрофилов, которые генерируют большие количества АФК, а также способствуют формированию тромбов в сосудах [26].
Таким образом, АФК-зависимый каскад реакций ухудшает микроциркуляцию, что приводит к формированию значительных интервалов подачи крови в ткани организма. В условиях возникшей гипоксии ослабляется дыхательная функция митохондрий и происходит перевосстановление редокс-центров в митохондриях (звено 6 на рис. 1). Прерывистая подача кислорода создает условия гипоксии – реоксигенации, при которых многократно возрастает синтез АФК [53], усиливается повреждение и тромбоз сосудов [54].
Терминальная стадия окислительного стресса: кризис антиоксидантной системы
Повышение уровня АФК выше определенного критического уровня индуцирует апоптоз и гибель клеток. Роль ловушек свободных радикалов в большинстве случаев выполняют цистеин и глутатион, функционирующие в составе пероксиредуктазной системы клетки. При высоком темпе генерации АФК происходит истощение восстановленных форм этих SH-реагентов, поскольку скорость работы системы их регенерации оказывается ниже скорости синтеза АФК. При истощении пула глутатиона теряется контроль над процессами синтеза и накопления АФК в клетке, происходит повреждение митохондрий [55] и клетка уходит в апоптоз [56] (звено 8 на рис. 1). При СД наблюдается дефицит глутатиона, связанный с недостатком цистеина и глицина, необходимых для его синтеза [11]. Снижение концентрации глицина в крови наблюдается уже на ранних стадиях развития СД [43] и является одной из причин раннего истощения антиоксидантной системы больных СД (звено 7 на рис. 1).
Роль ловушек свободных радикалов в большинстве случаев выполняют цистеин и глутатион, функционирующие в составе пероксиредуктазной системы клетки. При высоком темпе генерации АФК происходит истощение восстановленных форм этих SH-реагентов, поскольку скорость работы системы их регенерации оказывается ниже скорости синтеза АФК. При истощении пула глутатиона теряется контроль над процессами синтеза и накопления АФК в клетке, происходит повреждение митохондрий [55] и клетка уходит в апоптоз [56] (звено 8 на рис. 1). При СД наблюдается дефицит глутатиона, связанный с недостатком цистеина и глицина, необходимых для его синтеза [11]. Снижение концентрации глицина в крови наблюдается уже на ранних стадиях развития СД [43] и является одной из причин раннего истощения антиоксидантной системы больных СД (звено 7 на рис. 1).
Взаимосвязанный дефицит глицина и глутатиона может являться как следствием хронического воспалительного процесса, вызывающего ускоренный расход глутатиона, так и быть вызван иными причинами, включая повышенные скорости расщепления и выведения из организма этих соединений при диабете. Известно, что глицин и глутатион, аналогично глюкуроновой кислоте, конъюгируют с плохо растворимыми соединениями (ксенобиотики, ароматические соединения) для их выведения из организма почками [57]. Именно этот процесс может ускоряться при СД и являться причиной повышенного выделения глицина из организма. В частности, авторы работы [43] предполагают, что глицин может использоваться для выведения продуктов β-окисления и аминокислот с разветвленной цепью, концентрации которых в крови повышаются при СД.
Известно, что глицин и глутатион, аналогично глюкуроновой кислоте, конъюгируют с плохо растворимыми соединениями (ксенобиотики, ароматические соединения) для их выведения из организма почками [57]. Именно этот процесс может ускоряться при СД и являться причиной повышенного выделения глицина из организма. В частности, авторы работы [43] предполагают, что глицин может использоваться для выведения продуктов β-окисления и аминокислот с разветвленной цепью, концентрации которых в крови повышаются при СД.
Необходимо также отметить, что АФК-зависимый воспалительный процесс может сам быть причиной СД1 [1], так и СД2 [58]. Таким образом, гиперпродукция АФК, происходящая из-за формирования автокаталитического цикла и истощения антиоксидантной защиты, препятствует восстановлению нормальной инсулиновой сигнализации в организме больных. Это обстоятельство также может объяснять, почему уровни глицина в крови понижаются еще до манифестации болезни.
Механизмы ослабления патологических автокаталитических циклов глицином
Проведенная работа по систематизации метаболических нарушений при диабете и выявление их автокаталитического характера позволили показать, что наиболее перспективными мишенями для терапевтического воздействия являются процессы патологического синтеза АФК и нарушения микроциркуляции.
Механизм лекарственного действия глицина связан в первую очередь с его действием в качестве сигнальной молекулы, воздействующей на два вида важных рецепторов: глицин является агонистом стрихнин-чувствительных глициновых рецепторов (GlyR) и коагонистом глутаматных NMDA-рецепторов (NMDAR). Глициновые рецепторы локализованы не только в нервных клетках, но также в иммунных [20], эндотелиальных [32] и инсулинпроизводящих клетках поджелудочной железы [13]. Не менее важное значение играет метаболическое действие глицина, в частности, он участвует в синтезе ключевого компонента антиоксидантной защиты клеток – глутатиона. Глицин препятствует глюкозозависимой индукции автокаталитических процессов: увеличивает секрецию инсулина [13] и снижает глюконеогенез в печени [14], что должно способствовать нормализации метаболизма на самых ранних этапах заболевания (звенья 1–2 на рис. 1). В случае, если автокаталитический процесс уже запущен, глицин также способствует ослаблению ключевых положительных обратных связей, связанных с синтезом АФК и нарушением микроциркуляции. Конкретные звенья метаболических и сигнальных каскадов (см. рис. 1), на которые действует глицин, рассмотрены ниже.
Глициновые рецепторы локализованы не только в нервных клетках, но также в иммунных [20], эндотелиальных [32] и инсулинпроизводящих клетках поджелудочной железы [13]. Не менее важное значение играет метаболическое действие глицина, в частности, он участвует в синтезе ключевого компонента антиоксидантной защиты клеток – глутатиона. Глицин препятствует глюкозозависимой индукции автокаталитических процессов: увеличивает секрецию инсулина [13] и снижает глюконеогенез в печени [14], что должно способствовать нормализации метаболизма на самых ранних этапах заболевания (звенья 1–2 на рис. 1). В случае, если автокаталитический процесс уже запущен, глицин также способствует ослаблению ключевых положительных обратных связей, связанных с синтезом АФК и нарушением микроциркуляции. Конкретные звенья метаболических и сигнальных каскадов (см. рис. 1), на которые действует глицин, рассмотрены ниже.
Глицин снижает активность рецептора RAGE
Во-первых, глицин конкурентно ингибирует гликирование белков [9] (№2 в табл. 1), тем самым снижая образование КПГ и количество агонистов RAGE. Во-вторых, глицин снижает окислительный стресс: подавляет синтез АФК иммунными клетками [20] (№3, 6 в табл. 1), способствует поддержанию высокого уровня глутатиона [11] (№8 в табл. 1). Так как экспрессия гена рецептора RAGE осуществляется по сигналу NF-κB [50], который активируется АФК [19], то снижение окислительного стресса будет препятствовать синтезу белка RAGE. Таким образом, глицин снижает активность RAGE рецептора как за счет снижения концентраций его агонистов, так и за счет снижения АФК-зависимой экспрессии гена рецептора (звенья 2–3 на рис. 1).
1), тем самым снижая образование КПГ и количество агонистов RAGE. Во-вторых, глицин снижает окислительный стресс: подавляет синтез АФК иммунными клетками [20] (№3, 6 в табл. 1), способствует поддержанию высокого уровня глутатиона [11] (№8 в табл. 1). Так как экспрессия гена рецептора RAGE осуществляется по сигналу NF-κB [50], который активируется АФК [19], то снижение окислительного стресса будет препятствовать синтезу белка RAGE. Таким образом, глицин снижает активность RAGE рецептора как за счет снижения концентраций его агонистов, так и за счет снижения АФК-зависимой экспрессии гена рецептора (звенья 2–3 на рис. 1).
Таблица 1. Расшифровка рисунка 1 с указанием основных мишеней глицина
№ | Описание ключевых процессов | Биологический эффект глицина |
СТАДИЯ ИНДУКЦИИ ПАТОЛОГИЧЕСКОГО ПРОЦЕССА | ||
1 | При прогрессировании СД β-клетки поджелудочной железы или полностью погибают (1 тип), или заметно снижается их количество (2 тип). | Глицин увеличивает секрецию инсулина посредством стимуляции глицинового рецептора в β-клетках поджелудочной железы [13], что должно способствовать нормализации уровня глюкозы в крови при СД2 и на ранних стадиях СД1. |
2 | Нарушение биологического действия инсулина приводит к снижению активности транспорта глюкозы внутрь клеток, что ведет к повышению концентраций глюкозы и кетоновых тел (ввиду усиленного метаболизма жиров) в крови больных диабетом. Глюкоза и кетоновые тела вызывают неферментативное гликирование белков и липидов и образование конечных продуктов гликирования (КПГ). КПГ циркулируют в крови или удерживаются во внеклеточном матриксе. | Глицин снижает интенсивность глюконеогенеза в печени, что способствует снижению концентрации глюкозы в крови при СД [14]. |
3 | a) Повышение количества КПГ приводит к активации рецепторов RAGE, которые присутствуют во многих типах клеток, в том числе в эндотелиальных клетках, гладкой мускулатуре сосудов, макрофагах [16] и мастоцитах [17]. b) Активация RAGE включает синтез АФК NADPH-оксидазами (NOX) [18]. c) АФК активирует транскрипционный фактор NF-κB [19], который, с одной стороны, активирует антиоксидантную защиту клеток, а с другой – стимулирует развитие воспаления за счет выброса цитокинов. d) Повышение концентрации воспалительных цитокинов вызывает хемотаксис иммунных клеток в зону предвоспаления. Активация RAGE-рецепторов в макрофагах и мастоцитах дополнительно стимулирует их активность. В результате активности иммунной системы происходит дальнейшее увеличение концентрации АФК. | Глицин благодаря стимуляции глицинового рецептора обладает способностью подавлять избыточную активность иммунных клеток [20]. |
4 | При инсулинорезистентности или непосредственном недостатке инсулина происходит нарушение метаболизма липидов в печени, нарушается баланс липопротеидов и повышаются уровни холестерина и триглицеридов в крови [22]. | Глицин стимулирует NMDA-рецепторы в отделе мозга, контролирующем функцию печени, нормализует метаболизм липидов [23] и снижает глюконеогенез [14]. |
СТАДИЯ АКТИВАЦИИ ИШЕМИЧЕСКИХ ПРОЦЕССОВ ПРИ ДИАБЕТЕ | ||
5 | Нарушение микроциркуляции при диабете происходит по следующим причинам. Рано формирующийся атерогенный профиль липидов при СД ускоряет атеросклеротические изменения в стенках сосудов. Активация RAGE в клетках гладкой мускулатуры сосудов стимулирует их трансдифференцировку в остеобласты и кальцификацию сосудов [18]. Мастоциты вырабатывают гистамин, который расширяет сосуды, увеличивает проницаемость их стенок и приводит к резкому падению давления в капиллярах, агрегации эритроцитов и остановке тока крови [24]. Повышение концентрации АФК стимулирует перекисное окисление липидов и образование малонового диальдегида, который вызывает сшивки коллагена и повреждение сосудов [25]. В острой стадии воспаления нейтрофилы вызывают закупорку капилляров агрегатами из нейтрофилов, тромбоцитов и эритроцитов [26]. | Глицин нормализует уровни триглицеридов и холестерина в крови [27], препятствуя развитию атеросклероза при СД. Глицин способствует расширению сосудов у крыс в мозге [28], брюшине [29] и почках [30]. В отличие от гистамина, глицин не вызывает остановку тока крови и даже, наоборот, предотвращает этот негативный эффект гистамина [31]. Глицин при ишемии снижает перекисное окисление липидов и образование малонового диальдегида [33], уменьшая вторичное повреждение сосудов. |
6 | a) Нарушение микроциркуляции вызывает гипоксию тканей и снижение скорости митохондриального дыхания. При гипоксии прекращается окисление жирных кислот, происходит накопление жирных кислот и церамидов, блокирующих соответственно активности митохондриальных комплексов I и III [34]. В результате в условиях гипоксии основные переносчики электронов (NADH, коэнзим Q10, цитохром C) переходят в восстановленное состояние. При недостатке глюкозы (из-за нарушенного транспорта глюкозы при СД) клетки особенно уязвимы к гипоксии, так как прекращение окисления жирных кислот лишает клетку основного источника энергии. b) Спонтанная реоксигенация вызывает резкий всплеск генерации АФК. Усиление воспалительного сигнала (цитокины, АФК) приводит к хемотаксису в очаг ишемии нейтрофилов, которые вызывают еще больший выброс АФК [35]. | Глицин в острой фазе ишемии оказывает цитопротекторное действие, то есть предотвращает апоптоз и некроз [36–38]. Цитопротекция осуществляется за счет влияния на глициновые рецепторы. Глицин при гипоксии-реоксигенации снижает синтез АФК митохондриями, а также способствует восстановлению их фосфорилирующей активности [34, 39, 40], препятствует открыванию митохондриальной поры в кардиомиоцитах [41]. Активация глициновых рецепторов в нейтрофилах снижает выброс ими воспалительных цитокинов и генерацию АФК [42]. |
СТАДИЯ КРИЗИСА АНТИОКСИДАНТНОЙ СИСТЕМЫ | ||
7 | Дефицит глицина возникает уже на ранних стадиях развития СД2 [7]. Этому может способствовать его повышенное выведение из организма [43] (подробнее в основном тексте), увеличение количества глициндекарбоксилазы в гепатоцитах и ускорение его расщепления под влиянием глюкагона [44]. | Потребление глицина компенсирует его дефицит. Нормализация работы печени под действием глицина может способствовать в том числе снижению катаболизма глицина. Глицин также устраняет митохондриальную дисфункцию в клетках, испытывающих его дефицит [45]. |
8 | a) Существенное и долгосрочное повышение концентраций АФК способствует повреждению белков, истощению антиоксидантной системы и возникновению дефицита глутатиона [46]. Дефицит глутатиона обостряется в случае нехватки глицина и/или цистеина для его синтеза [11]. b) Неспособность антиоксидантной системы клеток нейтрализовать АФК ведет к нарастанию окислительных повреждений, дисфункции митохондрий и других органелл, включению стрессовых сигнальных систем и апоптозу. | Потребление глицина и цистеина восстанавливает темпы синтеза глутатиона до нормальных значений, снижает окислительный стресс у больных СД [11]. |
 При более выраженном нарушении функции β-клеток высокие уровни глюкозы в крови перестают подавлять секрецию глюкагона [3]. При СД2 в жировых тканях, мышцах и печени развивается инсулинорезистентность.
При более выраженном нарушении функции β-клеток высокие уровни глюкозы в крови перестают подавлять секрецию глюкагона [3]. При СД2 в жировых тканях, мышцах и печени развивается инсулинорезистентность.
 Сосудорасширяющий эффект глицина, вероятно, вызван его влиянием на глициновые рецепторы в эндотелиальных клетках [21, 32].
Сосудорасширяющий эффект глицина, вероятно, вызван его влиянием на глициновые рецепторы в эндотелиальных клетках [21, 32]. Высокие концентрации АФК усиливают NF-κB/RAGE каскад и повреждение микрососудов (обратная связь).
Высокие концентрации АФК усиливают NF-κB/RAGE каскад и повреждение микрососудов (обратная связь). Дефицит глицина усугубляется его повышенным расходом на синтез глутатиона [11].
Дефицит глицина усугубляется его повышенным расходом на синтез глутатиона [11].
Примечания: нумерация соответствует рисунку 1.
Глицин улучшает микроциркуляцию
Глицин препятствует формированию «порочного круга», связанного с ишемией: способствует нормализации метаболизма в печени и снижению уровня холестерина в крови [23], уменьшает патологическую активность иммунных клеток [20], снижает перекисное окисление липидов и повреждения микрососудов [33] (подробнее в №4–6 в табл. 1). В экспериментальных работах нашего института показано, что глицин вызывает расширение микрососудов у крыс в мозге [28] и брюшине [29]. Важно отметить, что сосудорасширяющий эффект глицина принципиально отличается от действия гистамина. В отличие от гистамина, являющегося типичным медиатором воспаления, глицин не вызывает агрегации эритроцитов и не увеличивает проницаемость сосудов, благодаря чему не вызывает остановки тока крови в капиллярах. Более того, непосредственно показано, что глицин препятствует остановке тока крови под влиянием гистамина и восстанавливает нарушенную гистамином микроциркуляцию в артериолах брюшины [31]. Быстрая динамика наблюдаемого сосудорасширяющего эффекта глицина (1–3 минуты) позволяет предполагать, что глицин улучшает микроциркуляцию посредством действия на глициновые рецепторы в эндотелиальных клетках [32]. Моделирование пространственно-временных распределений концентрации глюкозы вблизи кровеносных сосудов на примере пиальных оболочек крыс показывает, что наблюдаемое экспериментально увеличение калибра артериол обуславливает возрастание амплитуды градиента данного углевода в ткани [59]. Сосудорасширяющее действие глицина в почках может также объясняться активацией NMDAR [30]. Таким образом, за счет предотвращения повреждения микрососудов, сосудорасширяющего и противогистаминного действия глицин нормализует микроциркуляцию, устраняя ключевую причину нарастания окислительного стресса (звено 5 на рис. 1).
Быстрая динамика наблюдаемого сосудорасширяющего эффекта глицина (1–3 минуты) позволяет предполагать, что глицин улучшает микроциркуляцию посредством действия на глициновые рецепторы в эндотелиальных клетках [32]. Моделирование пространственно-временных распределений концентрации глюкозы вблизи кровеносных сосудов на примере пиальных оболочек крыс показывает, что наблюдаемое экспериментально увеличение калибра артериол обуславливает возрастание амплитуды градиента данного углевода в ткани [59]. Сосудорасширяющее действие глицина в почках может также объясняться активацией NMDAR [30]. Таким образом, за счет предотвращения повреждения микрососудов, сосудорасширяющего и противогистаминного действия глицин нормализует микроциркуляцию, устраняя ключевую причину нарастания окислительного стресса (звено 5 на рис. 1).
Глицин нормализует структуру и функцию митохондрий
Результаты экспериментов, проведенных в нашем институте, показывают, что глицин снижает синтез АФК митохондриями после гипоксии в мозге [39] и в сердце [34]. Кроме того, в тканях мозга при аноксии глицин предотвращает нарушения ультраструктуры митохондрий [40]. Согласно литературным данным, глицин препятствует апоптозу в сердечных тканях за счет предотвращения открывания митохондриальной поры [41]. Необходимо также отметить, что дефицит глицина сам по себе может служить причиной повреждения митохондрий, а его восполнение восстанавливает их функцию [45]. Приведенные данные показывают, что глицин нормализует работу митохондрий и снижает синтез ими АФК, а также предотвращает митохондриально-зависимый апоптоз (звено 6 на рис. 1). Способность глицина нормализовать функцию митохондрий представляется важной также в той связи, что высокие темпы митохондриального дыхания обеспечивают высокую чувствительность к инсулину и препятствуют развитию СД2 [60].
Кроме того, в тканях мозга при аноксии глицин предотвращает нарушения ультраструктуры митохондрий [40]. Согласно литературным данным, глицин препятствует апоптозу в сердечных тканях за счет предотвращения открывания митохондриальной поры [41]. Необходимо также отметить, что дефицит глицина сам по себе может служить причиной повреждения митохондрий, а его восполнение восстанавливает их функцию [45]. Приведенные данные показывают, что глицин нормализует работу митохондрий и снижает синтез ими АФК, а также предотвращает митохондриально-зависимый апоптоз (звено 6 на рис. 1). Способность глицина нормализовать функцию митохондрий представляется важной также в той связи, что высокие темпы митохондриального дыхания обеспечивают высокую чувствительность к инсулину и препятствуют развитию СД2 [60].
Глицин защищает клетки от апоптоза
В острой фазе ишемии глицин защищает клетки от апоптоза, позволяя сохранить функциональность тканей и органов, включая мозг, сердце, почки, печень и сосуды [37]. Эффективность глицина в качестве противоишемического препарата подтверждена клиническими исследованиями [36], глицин назначают при реабилитации после ишемических инсультов. Особенности цитопротекторного действия глицина при ишемии подробно разобраны в обзоре [38], и выделен ряд характерных для него признаков, а именно: глицин имеет быструю кинетику действия и не требует транспортировки внутрь клетки; глицин должен присутствовать во время острой фазы ишемии; максимальный защитный эффект глицина достигается при его внеклеточной концентрации около 2 мМ. Перечисленные признаки указывают на то, что цитопротекторное действие глицина, возможно, осуществляется посредством особой формы глицинового рецептора, активация которой предотвращает повреждение и патологическую проницаемость клеточных мембран при ишемии. Дополнительный прием глицина и цистеина также позволяет повысить скорость синтеза глутатиона и компенсировать его дефицит при СД [11]. Таким образом, глицин препятствует терминальной стадии развития окислительного стресса, ведущей к апоптозу (или некрозу) клеток (звенья 7–8 на рис.
Эффективность глицина в качестве противоишемического препарата подтверждена клиническими исследованиями [36], глицин назначают при реабилитации после ишемических инсультов. Особенности цитопротекторного действия глицина при ишемии подробно разобраны в обзоре [38], и выделен ряд характерных для него признаков, а именно: глицин имеет быструю кинетику действия и не требует транспортировки внутрь клетки; глицин должен присутствовать во время острой фазы ишемии; максимальный защитный эффект глицина достигается при его внеклеточной концентрации около 2 мМ. Перечисленные признаки указывают на то, что цитопротекторное действие глицина, возможно, осуществляется посредством особой формы глицинового рецептора, активация которой предотвращает повреждение и патологическую проницаемость клеточных мембран при ишемии. Дополнительный прием глицина и цистеина также позволяет повысить скорость синтеза глутатиона и компенсировать его дефицит при СД [11]. Таким образом, глицин препятствует терминальной стадии развития окислительного стресса, ведущей к апоптозу (или некрозу) клеток (звенья 7–8 на рис. 1).
1).
Клинические доказательства действия глицина при сахарном диабете
Проведенный поиск в базах данных научной литературы показал, что исследования действия глицина на больных СД проведены в последние годы в разных странах. Основные параметры проведенных клинических исследований отображены в табл. 2. Несмотря на существенные различия исследуемых групп (различные типы диабета и выраженности диабетических осложнений), во всех работах отмечается положительная динамика состояния больных СД при терапии глицином. Приведенные данные показывают, что наиболее заметно действие глицина отмечалось на параметры, связанные с микроциркуляцией и интенсивностью воспалительных процессов, что находится в согласии с результатами проведенного в настоящей работе анализа биологического действия глицина. Отдельно стоит отметить, что использование в терапии очень высоких доз глицина (20 г в день), а также его потребление натощак [12] могут провоцировать повышение секреции глюкагона и приводить к повышению концентрации глюкозы в крови, а в долгосрочной перспективе – к повышению уровня гликированного гемоглобина [10]. В то же время умеренные дозы глицина вместе с пищей (до 5 г), наоборот, способствуют снижению гликирования [9], что совпадает с результатами испытаний на модельных животных [15]. Также следует подчеркнуть, что ни в одном из приведенных исследований не отмечается заметных побочных эффектов и жалоб пациентов на ухудшение состояния при потреблении глицина.
В то же время умеренные дозы глицина вместе с пищей (до 5 г), наоборот, способствуют снижению гликирования [9], что совпадает с результатами испытаний на модельных животных [15]. Также следует подчеркнуть, что ни в одном из приведенных исследований не отмечается заметных побочных эффектов и жалоб пациентов на ухудшение состояния при потреблении глицина.
Таблица 2. Краткое описание клинических исследований глицина при сахарном диабете
Исследуемая группа/контроль | Дозировка глицина, сроки | Основные результаты исследования | Страна, методика, ссылка |
Больные СД1 с выраженной энцефалопатией: 31 чел./28 чел. | 100 мг глицина (+50 мг лимонной и 200 мг янтарной кислот), 3 раза в день, 3 мес | На 16,8% снизилось число пациентов с препролиферативной и пролиферативной ретинопатией (переход в непролиферативную форму). Значимая положительная динамика показателей тромбоцитарного звена гемостаза: размера агрегатов (25,6%) и скорости агрегации (20,5%). | Россия, открытое рандомизированное исследование [8] |
Больные СД2 без осложнений: 38 чел./36 чел. | 5 г глицина один раз в день, 3 мес | В группе, принимавшей глицин, значимо (и сильнее, чем в группе плацебо) снизился гликированный гемоглобин (с 8,3±1,9 до 6,9±1,3%). Интерферон-гамма повысился, снизилась секреция рецептора фактора некроза опухоли TNFα (воспалительный маркер) | Мексика, двойное слепое исследование [9] |
28 больных СД2 со слуховой невропатией/ 15 больных СД2 без нарушений | 5 г глицина 4 раза в день, 6 мес | В группе, получавшей глицин, улучшилась проводимость слухового нерва, в контрольной группе ухудшилась. Глицин повысил показатели глюкозы натощак и гликированного гемоглобина до уровня контроля (с 7,2±1,9 до 8,1±1,8). (Эффект может быть вызван стимуляцией секреции глюкагона высокими дозами аминокислоты) | Мексика, открытое исследование [10] |
12 чел. | 100 мг/кг глицина (+100 мг/кг цистеина) в день, 14 дней | Концентрация глутатиона в эритроцитах больных СД2 существенно ниже, чем в контрольной группе (1,65±0,16 vs. 6,75±0,47 мкмоль/г гемоглобина). Терапия привела к повышению концентрации глутатиона на 64,4%, увеличению скорости его синтеза, снижению окислительного стресса | США, открытое исследование [11] |
9 здоровых чел./ нет | 75 мг/кг глицина, одноразово | Глицин в отсутствие глюкозы увеличивает секрецию глюкагона. Потребление глицина вместе с глюкозой снижает максимальный подъем уровня глюкозы в крови, увеличивает скорость ее поглощения тканями | США, открытое исследование [12] |
 Значимо улучшились показатели неврологического и нейропсихологического статуса всех больных
Значимо улучшились показатели неврологического и нейропсихологического статуса всех больных с СД2/ 12 здоровых чел., схожих по возрасту, полу, индексу массы тела
с СД2/ 12 здоровых чел., схожих по возрасту, полу, индексу массы телаЗаключение
В настоящей работе сформулирована структурированная модель стадийного развития заболевания, которое возникает в условиях сильного ослабления или исчезновения инсулинового сигнала. В модели выделена начальная инсулин- и глюкозозависимая стадия заболевания, которая включает два первичных параллельно протекающих процесса. Во-первых, это процессы гликирования белков и липидов, которые активируют RAGE-рецептор и включают процессы синтеза АФК. Во-вторых, это нарушение метаболизма липидов в печени, которое приводит к повышению уровня холестерина в крови, развитию и прогрессированию эндотелиальной дисфункции.
В модели выделена начальная инсулин- и глюкозозависимая стадия заболевания, которая включает два первичных параллельно протекающих процесса. Во-первых, это процессы гликирования белков и липидов, которые активируют RAGE-рецептор и включают процессы синтеза АФК. Во-вторых, это нарушение метаболизма липидов в печени, которое приводит к повышению уровня холестерина в крови, развитию и прогрессированию эндотелиальной дисфункции.
Вторая стадия заболевания не связана непосредственно с нарушением метаболизма глюкозы и сигналом инсулина. Модель последующего развития заболевания построена на взаимодействии двух групп параллельно протекающих патологических процессов, которые эффективно усиливают друг друга. Каскады АФК-зависимых биохимических реакций и процесс постадийного нарушения микроциркуляции связаны друг с другом положительными обратными связями, в результате чего возникает автономно усиливающийся автокаталитический цикл нарушения метаболизма и АФК сигнализации тканей (рис. 2). Этот цикл проявляется как известный эффект «метаболической памяти» и является основой глюкозонезависимой стадии развития диабета.
Рис. 2. Схема основного автокаталитического цикла взаимного усиления АФК-зависимых процессов и патофизиологического процесса нарушения микроциркуляции. Степень активности иммунных клеток растет за счет выброса воспалительных цитокинов при АФК-зависимой активации NF-κB. В результате автокаталитического нарастания окислительного стресса происходит истощение антиоксидантной защиты клеток и апоптоз (или некроз). Сокращения: АФК – активные формы кислорода; КПГ – конечные продукты гликирования; RAGE – рецепторы КПГ; NOX – NADPH-оксидазы.
Полученный результат позволяет по-новому рассматривать СД как заболевание, которое несет в себе важнейшие черты самоускоряющегося циклического автокаталитического процесса. Очевидно, что при автокаталитическом цикле нарушение значимых положительных обратных связей позволяет остановить или резко затормозить весь циклический процесс в целом. В связи с этим существует перспектива нового подхода к лечению этого заболевания путем целевого блокирования автокатализа при диабете. С другой стороны, можно предполагать, что, согласно принятой модели, наиболее сильного терапевтического эффекта удастся добиться путем направленного одновременного блокирования обеих систем: АФК-связанных процессов и патофизиологического каскада нарушения микроциркуляции.
С другой стороны, можно предполагать, что, согласно принятой модели, наиболее сильного терапевтического эффекта удастся добиться путем направленного одновременного блокирования обеих систем: АФК-связанных процессов и патофизиологического каскада нарушения микроциркуляции.
На основании полученных результатов и анализа биологического действия глицина в настоящей работе удалось показать, что эта аминокислота является блокатором как биохимического каскада реакций, связанных с синтезом АФК, так и процессов нарушения микроциркуляции, в результате чего глицин должен эффективно ослаблять основной автокаталитический цикл при диабете (см. рис. 2). Таким образом, проделанная работа выявила ключевые звенья изменения метаболизма при диабете, которые являются потенциальными мишенями воздействия лекарственных препаратов, в том числе аминокислоты глицин.
Дополнительная информация
Источник финансирования. Поисково-аналитическая работа по подготовке рукописи проведена при финансовом обеспечении института цитохимии и молекулярной фармакологии.
Конфликт интересов. Авторы декларируют отсутствие явных и потенциальных конфликтов интересов, связанных с публикацией настоящей статьи.
Участие авторов. Нестеров С.В. – сбор и обработка материалов, написание текста; Ягужинский Л.С. – анализ данных, написание текста; Подопригора Г.И. – концепция исследования, редактирование текста рукописи; Нарциссов Я.Р. – концепция исследования, редактирование текста рукописи. Все авторы внесли существенный вклад в проведение поисково-аналитической работы и подготовку статьи, прочли и одобрили финальную версию перед публикацией.
1. Marre ML, Piganelli JD. Environmental Factors Contribute to beta Cell Endoplasmic Reticulum Stress and Neo-Antigen Formation in Type 1 Diabetes. Front Endocrinol (Lausanne). 2017;8:262. doi: 10.3389/fendo.2017.00262
2. Montgomery MK, Turner N. Mitochondrial dysfunction and insulin resistance: an update. Endocr Connect. 2015;4(1):R1-R15. doi: 10.1530/EC-14-0092
3. Li C, Liu C, Nissim I, et al. Regulation of glucagon secretion in normal and diabetic human islets by gamma-hydroxybutyrate and glycine. J Biol Chem. 2013;288(6):3938-3951. doi: 10.1074/jbc.M112.385682
Li C, Liu C, Nissim I, et al. Regulation of glucagon secretion in normal and diabetic human islets by gamma-hydroxybutyrate and glycine. J Biol Chem. 2013;288(6):3938-3951. doi: 10.1074/jbc.M112.385682
4. Дедов И.И., Шестакова М.В., Викулова О.К. Государственный регистр сахарного диабета в Российской Федерации: статус 2014 г. и перспективы развития // Сахарный диабет. – 2015. – Т. 18. – №3. – С. 5-22. [Dedov II, Shestakova MV, Vikulova OK. National register of diabetes mellitus in Russian Federation. Diabetes Mellitus. 2015;18(3):5-22. (In Russ.)] doi: 10.14341/DM201535-22
5. Lustgarten MS, Price LL, Phillips EM, Fielding RA. Serum glycine is associated with regional body fat and insulin resistance in functionally-limited older adults. PLoS One. 2013;8(12):e84034. doi: 10.1371/journal.pone.0084034
6. Palmer ND, Stevens RD, Antinozzi PA, et al. Metabolomic profile associated with insulin resistance and conversion to diabetes in the Insulin Resistance Atherosclerosis Study. J Clin Endocrinol Metab. 2015;100(3):E463-468. doi: 10.1210/jc.2014-2357
J Clin Endocrinol Metab. 2015;100(3):E463-468. doi: 10.1210/jc.2014-2357
7. Wang-Sattler R, Yu Z, Herder C, et al. Novel biomarkers for pre-diabetes identified by metabolomics. Mol Syst Biol. 2012;8:615. doi: 10.1038/msb.2012.43
8. Чуйко М.Р., Ефремова Н.М., Скворцова В.И. Эффективность и безопасность применения глицина и лимонтара в комплексной терапии дисциркуляторной энцефалопатии и энцефалопатии при инсулинзависимом сахарном диабете // Журнал неврологии и психиатрии им. С.С. Корсакова. – 2010. – Т. 110. – №6. – С. 44-48. [Chuyko MR, Efremova NM, Skvortsova VI. Efficacy and safety of glycine and limontar in the complex therapy of discirculatory encephalopathy and encephalopathy in diabetes mellitus type I. Zh Nevrol Psikhiatr Im SS Korsakova. 2010;110(6):44-48. (In Russ.)]
9. Cruz M, Maldonado-Bernal C, Mondragón-Gonzalez R, et al. Glycine treatment decreases proinflammatory cytokines and increases interferon-γ in patients with Type 2 diabetes. J Endocrinol Invest. 2014;31(8):694-699. doi: 10.1007/bf03346417
2014;31(8):694-699. doi: 10.1007/bf03346417
10. Munoz-Carlin Mde L, Rodriguez-Moctezuma JR, Gomez Latorre JG, et al. Effects of glycine on auditory evoked potentials among diabetic patients with auditory pathway neuropathy. Rev Med Chil. 2010;138(10):1246-1252. doi: /S0034-98872010001100006
11. Sekhar RV, McKay SV, Patel SG, et al. Glutathione synthesis is diminished in patients with uncontrolled diabetes and restored by dietary supplementation with cysteine and glycine. Diabetes Care. 2011;34(1):162-167. doi: 10.2337/dc10-1006
12. Gannon MC, Nuttall JA, Nuttall FQ. The metabolic response to ingested glycine. Am J Clin Nutr. 2002;76(6):1302-1307. doi: 10.1093/ajcn/76.6.1302
13. Yan-Do R, Duong E, Manning Fox JE, et al. A Glycine-Insulin Autocrine Feedback Loop Enhances Insulin Secretion From Human beta-Cells and Is Impaired in Type 2 Diabetes. Diabetes. 2016;65(8):2311-2321. doi: 10.2337/db15-1272
14. Lam CK, Chari M, Su BB, et al. Activation of N-methyl-D-aspartate (NMDA) receptors in the dorsal vagal complex lowers glucose production. J Biol Chem. 2010;285(29):21913-21921. doi: 10.1074/jbc.M109.087338
J Biol Chem. 2010;285(29):21913-21921. doi: 10.1074/jbc.M109.087338
15. Alvarado-Vásquez N, Zamudio P, Cerón E, et al. Effect of glycine in streptozotocin-induced diabetic rats. Comp Biochem Physiol C Toxicol Pharmacol. 2003;134(4):521-527. doi: 10.1016/s1532-0456(03)00046-2
16. Hudson BI, Bucciarelli LG, Wendt T, et al. Blockade of receptor for advanced glycation endproducts: a new target for therapeutic intervention in diabetic complications and inflammatory disorders. Arch Biochem Biophys. 2003;419(1):80-88. doi: 10.1016/j.abb.2003.08.030
17. Sick E, Brehin S, André P, et al. Advanced glycation end products (AGEs) activate mast cells. Br J Pharmacol. 2010;161(2):442-455. doi: 10.1111/j.1476-5381.2010.00905.x
18. Kay AM, Simpson CL, Stewart JA, Jr. The Role of AGE/RAGE Signaling in Diabetes-Mediated Vascular Calcification. J Diabetes Res. 2016;2016:6809703. doi: 10.1155/2016/6809703
19. Morgan MJ, Liu ZG. Crosstalk of reactive oxygen species and NF-kappaB signaling. Cell Res. 2011;21(1):103-115. doi: 10.1038/cr.2010.178
Cell Res. 2011;21(1):103-115. doi: 10.1038/cr.2010.178
20. Froh M, Thurman RG, Wheeler MD. Molecular evidence for a glycine-gated chloride channel in macrophages and leukocytes. Am J Physiol Gastrointest Liver Physiol. 2002;283(4):G856-863. doi: 10.1152/ajpgi.00503.2001
21. McCarty MF, Barroso-Aranda J, Contreras F. The hyperpolarizing impact of glycine on endothelial cells may be anti-atherogenic. Med Hypotheses. 2009;73(2):263-264. doi: 10.1016/j.mehy.2008.12.021
22. Borggreve SE, de Vries R, Dullaart RPF. Alterations in high-density lipoprotein metabolism and reverse cholesterol transport in insulin resistance and type 2 diabetes mellitus: role of lipolytic enzymes, lecithin:cholesterol acyltransferase and lipid transfer proteins. Eur J Clin Invest. 2003;33(12):1051-1069. doi: 10.1111/j.1365-2362.2003.01263.x
23. Yue JT, Mighiu PI, Naples M, et al. Glycine normalizes hepatic triglyceride-rich VLDL secretion by triggering the CNS in high-fat fed rats. Circ Res. 2012;110(10):1345-1354. doi: 10.1161/CIRCRESAHA.112.268276
2012;110(10):1345-1354. doi: 10.1161/CIRCRESAHA.112.268276
24. Hammdy N, Salam R, El GNA, Mahmoud E. Mast cell a new player in Type 2 diabetes. Endocrine Abstracts. 2016. doi: 10.1530/endoabs.41.EP476
25. Slatter DA, Bolton CH, Bailey AJ. The importance of lipid-derived malondialdehyde in diabetes mellitus. Diabetologia. 2000;43(5):550-557. doi: 10.1007/s001250051342
26. Looney MR, Matthay MA. Neutrophil sandwiches injure the microcirculation. Nat Med. 2009;15(4):364-366. doi: 10.1038/nm0409-364
27. Sugiyama K, Kanamori H, Tanaka S. Correlation of the Plasma Cholesterol-lowering Effect of Dietary Glycine with the Alteration of Hepatic Phospholipid Composition in Rats. Biosci Biotechnol Biochem. 2014;57(9):1461-1465. doi: 10.1271/bbb.57.1461
28. Podoprigora GI, Nartsissov YR, Aleksandrov PN. Effect of Glycine on Microcirculation in Pial Vessels of Rat Brain. Bull Exp Biol Med. 2005;139(6):675-677. doi: 10.1007/s10517-005-0375-2
29. Podoprigora GI, Nartsissov YR. Effect of Glycine on the Microcirculation in Rat Mesenteric Vessels. Bull Exp Biol Med. 2009;147(3):308-311. doi: 10.1007/s10517-009-0498-y
Effect of Glycine on the Microcirculation in Rat Mesenteric Vessels. Bull Exp Biol Med. 2009;147(3):308-311. doi: 10.1007/s10517-009-0498-y
30. Deng A. Vasodilatory N-Methyl-D-Aspartate Receptors Are Constitutively Expressed in Rat Kidney. J Am Soc Nephrol. 2002;13(5):1381-1384. doi: 10.1097/01.asn.0000013293.11876.4e
31. Podoprigora GI, Blagosklonov O, Angoue O, et al. Assessment of microcirculatory effects of glycine by intravital microscopy in rats. Conf Proc IEEE Eng Med Biol Soc. 2012;2012:2651-2654. doi: 10.1109/EMBC.2012.6346509
32. Yamashina S, Konno A, Wheeler MD, et al. Endothelial cells contain a glycine-gated chloride channel. Nutr Cancer. 2001;40(2):197-204. doi: 10.1207/S15327914NC402_17
33. Meyer KF, Martins JL, Freitas Filho LGd, et al. Glycine reduces tissue lipid peroxidation in hypoxia-reoxygenation-induced necrotizing enterocolitis in rats. Acta Cir Bras. 2006;21(3):161-167. doi: 10.1590/s0102-86502006000300008
34. Selin AA, Lobysheva NV, Nesterov SV, et al. On the regulative role of the glutamate receptor in mitochondria. Biol Chem. 2016;397(5):445-458. doi: 10.1515/hsz-2015-0289
On the regulative role of the glutamate receptor in mitochondria. Biol Chem. 2016;397(5):445-458. doi: 10.1515/hsz-2015-0289
35. Schofield ZV, Woodruff TM, Halai R, et al. Neutrophils—a key component of ischemia-reperfusion injury. Shock. 2013;40(6):463-470. doi: 10.1097/SHK.0000000000000044
36. Gusev EI, Skvortsova VI, Dambinova SA, et al. Neuroprotective effects of glycine for therapy of acute ischaemic stroke. Cerebrovasc Dis. 2000;10(1):49-60. doi: 10.1159/000016025
37. Van den Eynden J, Ali SS, Horwood N, et al. Glycine and glycine receptor signalling in non-neuronal cells. Front Mol Neurosci. 2009;2:9. doi: 10.3389/neuro.02.009.2009
38. Weinberg JM, Bienholz A, Venkatachalam MA. The role of glycine in regulated cell death. Cell Mol Life Sci. 2016;73(11-12):2285-2308. doi: 10.1007/s00018-016-2201-6
39. Selin AA, Lobysheva NV, Vorontsova ON, et al. Mechanism Underlying the Protective Effect of Glycine in Energetic Disturbances in Brain Tissues under Hypoxic Conditions. Bull Exp Biol Med. 2012;153(1):44-47. doi: 10.1007/s10517-012-1638-3
Bull Exp Biol Med. 2012;153(1):44-47. doi: 10.1007/s10517-012-1638-3
40. Tonshin AA, Lobysheva NV, Yaguzhinsky LS, et al. Effect of the inhibitory neurotransmitter glycine on slow destructive processes in brain cortex slices under anoxic conditions. Biochemistry (Moscow). 2007;72(5):509-517. doi: 10.1134/s0006297907050070
41. Ruiz-Meana M, Pina P, Garcia-Dorado D, et al. Glycine protects cardiomyocytes against lethal reoxygenation injury by inhibiting mitochondrial permeability transition. J Physiol. 2004;558(Pt 3):873-882. doi: 10.1113/jphysiol.2004.068320
42. Wheeler M, Stachlewitz RF, Yamashina S, et al. Glycine-gated chloride channels in neutrophils attenuate calcium influx and superoxide production. FASEB J. 2000;14(3):476-484. doi: 10.1096/fasebj.14.3.476
43. Yan-Do R, MacDonald PE. Impaired «Glycine»-mia in Type 2 Diabetes and Potential Mechanisms Contributing to Glucose Homeostasis. Endocrinology. 2017;158(5):1064-1073. doi: 10.1210/en.2017-00148
44. Jog R, Wang J, Leff T. Hormonal Regulation of Glycine Metabolism And Its Potential Role in Diabetes Susceptibility. FASEB J. 2017;31(1 Supplement):626.
Jog R, Wang J, Leff T. Hormonal Regulation of Glycine Metabolism And Its Potential Role in Diabetes Susceptibility. FASEB J. 2017;31(1 Supplement):626.
45. Hashizume O, Ohnishi S, Mito T, et al. Epigenetic regulation of the nuclear-coded GCAT and SHMT2 genes confers human age-associated mitochondrial respiration defects. Sci Rep. 2015;5:10434. doi: 10.1038/srep10434
46. Ramen T M. Depletion of Glutathione during Oxidative Stress and Efficacy of N-Acetyl Cysteine: An Old Drug with New Approaches. Med Chem (Los Angeles). 2015;05(01). doi: 10.4172/2161-0444.1000240
47. Ceriello A. The emerging challenge in diabetes: the «metabolic memory». Vascul Pharmacol. 2012;57(5-6):133-138. doi: 10.1016/j.vph.2012.05.005
48. San Martin A, Foncea R, Laurindo FR, et al. Nox1-based NADPH oxidase-derived superoxide is required for VSMC activation by advanced glycation end-products. Free Radic Biol Med. 2007;42(11):1671-1679. doi: 10.1016/j.freeradbiomed.2007.02.002
49. Serban AI, Stanca L, Geicu OI, Dinischiotu A. AGEs-Induced IL-6 Synthesis Precedes RAGE Up-Regulation in HEK 293 Cells: An Alternative Inflammatory Mechanism? Int J Mol Sci. 2015;16(9):20100-20117. doi: 10.3390/ijms160920100
AGEs-Induced IL-6 Synthesis Precedes RAGE Up-Regulation in HEK 293 Cells: An Alternative Inflammatory Mechanism? Int J Mol Sci. 2015;16(9):20100-20117. doi: 10.3390/ijms160920100
50. Li J, Schmidt AM. Characterization and Functional Analysis of the Promoter of RAGE, the Receptor for Advanced Glycation End Products. J Biol Chem. 1997;272(26):16498-16506. doi: 10.1074/jbc.272.26.16498
51. Huebschmann AG, Regensteiner JG, Vlassara H, Reusch JE. Diabetes and advanced glycoxidation end products. Diabetes Care. 2006;29(6):1420-1432. doi: 10.2337/dc05-2096
52. Bierhaus A, Schiekofer S, Schwaninger M, et al. Diabetes-Associated Sustained Activation of the Transcription Factor Nuclear Factor- κB. Diabetes. 2001;50(12):2792-2808. doi: 10.2337/diabetes.50.12.2792
53. Li C, Jackson RM. Reactive species mechanisms of cellular hypoxia-reoxygenation injury. Am J Physiol Cell Physiol. 2002;282(2):C227-241. doi: 10.1152/ajpcell.00112.2001
54. Tang YH, Vital S, Russell J, et al. Transient ischemia elicits a sustained enhancement of thrombus development in the cerebral microvasculature: effects of anti-thrombotic therapy. Exp Neurol. 2014;261:417-423. doi: 10.1016/j.expneurol.2014.07.004
Transient ischemia elicits a sustained enhancement of thrombus development in the cerebral microvasculature: effects of anti-thrombotic therapy. Exp Neurol. 2014;261:417-423. doi: 10.1016/j.expneurol.2014.07.004
55. Ghosh S, Pulinilkunnil T, Yuen G, et al. Cardiomyocyte apoptosis induced by short-term diabetes requires mitochondrial GSH depletion. Am J Physiol Heart Circ Physiol. 2005;289(2):H768-776. doi: 10.1152/ajpheart.00038.2005
56. Franco R, Cidlowski JA. Apoptosis and glutathione: beyond an antioxidant. Cell Death Differ. 2009;16(10):1303-1314. doi: 10.1038/cdd.2009.107
57. Badenhorst CP, Erasmus E, van der Sluis R, et al. A new perspective on the importance of glycine conjugation in the metabolism of aromatic acids. Drug Metab Rev. 2014;46(3):343-361. doi: 10.3109/03602532.2014.908903
58. Shoelson SE, Lee J, Goldfine AB. Inflammation and insulin resistance. J Clin Invest. 2006;116(7):1793-1801. doi: 10.1172/JCI29069
59. Nartsissov YR, Tyukina ES, Boronovsky SE, Sheshegova EV. Computer modeling of spatial-time distribution of metabolite concentrations in phantoms of biological objects by example of rat brain pial. Biophysics. 2014;58(5):703-711. doi: 10.1134/s0006350913050102
Computer modeling of spatial-time distribution of metabolite concentrations in phantoms of biological objects by example of rat brain pial. Biophysics. 2014;58(5):703-711. doi: 10.1134/s0006350913050102
60. Cho J, Zhang Y, Park SY, et al. Mitochondrial ATP transporter depletion protects mice against liver steatosis and insulin resistance. Nat Commun. 2017;8:14477. doi: 10.1038/ncomms14477
ГЛИЦИН — инструкция, состав, применение, дозировка, показания, противопоказания, отзывы
Лікар-педіатр
Дозировка:
таблетки сублингв. по 100 мг №50
Производитель:
ООО Арпимед, Республика Армения, Котайкська обл., г. Абовян, 2-й микрорайон, д. 19.
Адаптированная инструкция
Состав:
Лекарственная форма:
таблетки
Производитель:
Непосредственное производство: ООО Арпимед, Республика Армения, Котайкська обл. , г. Абовян, 2-й микрорайон, д. 19.
, г. Абовян, 2-й микрорайон, д. 19.
Фармакотерапевтическая группа:
Средства, действующие на нервную сиcтему.
Фармакологические свойства:
Показания к применению:
Противопоказания:
Сверхчувствительность к составляющим л/с, пониженное давление, дети, младше трех лет.
Надлежащие меры безопасности при применении:
Глицин таблетки обладают гипотензивным эффектом. Это следует учитывать людям с пониженным давлением.
Применение в период беременности или кормления грудью:
В виду отсутствия доказательств о безопасности, таблетки Глицин не рекомендованы.
Способность влиять на скорость реакции при управлении автотранспортом:
Стоит быть осмотрительным при вождении/управлении автомеханизмами.
Способ применения и дозы:
Передозировка:
Сообщения отсутствуют.
Побочные действия:
Аллергия в виде насморка, раздраженной слизистой в горле, крапивницы, конъюнктивита и общей слабости.
Лекарственное взаимодействие:
Срок годности:
36 месяцев.
Условия хранения:
T помещения: 25 С, вдали от детей.
Форма выпуска / упаковка:
50 таб-ок в пачке из картона.
Категория отпуска:
Без рецепта.
Внимание
Приведенная научная информация является обобщающей, основана на официально утвержденной инструкции по применению и не может быть использована для принятия решения о возможности применения конкретного лекарственного препарата.
Официальная инструкция
Состав:
Лекарственная форма:
таблетки
Основные физико-химические свойства:
Производитель:
ООО Арпимед
Местонахождение производителя:
Республика Армения, Котайкська обл., г. Абовян, 2-й микрорайон, д. 19.
Фармакотерапевтическая группа:
Средства, действующие на нервную сиcтему.
Фармакологические свойства:
Показания к применению:
Противопоказания:
Индивидуальная непереносимость препарата и повышенная чувствительность к отдельным его компонентов; артериальная гипотензия. Возраст до 3 лет.
Возраст до 3 лет.
Особенности применения:
Пациентам со склонностью к артериальной гипотензии необходимо контролировать уровень артериального давления и при необходимости проводить коррекцию дозы препарата. Глицин назначают в меньших дозах и при условии регулярного контроля артериального давления. При показателях артериального давления ниже обычного уровня прием препарата прекращают.
Применение в период беременности или кормления грудью:
Влияние Глицина на организм в период беременности и кормления грудью подробно не исследовался, поэтому применение препарата не рекомендуется.
Способность влиять на скорость реакции при управлении автотранспортом:
Необходимо соблюдать осторожность при управлении автотранспортом или работе с другими механизмами, а также занятии потенциально опасными видами деятельности.
Дети:
Препарат применяют детям в возрасте от 3 лет.
Способ применения и дозы:
Передозировка:
О клинических проявлениях передозировки сведений нет.
Побочные действия:
В отдельных случаях при индивидуальной повышенной чувствительности возможно развитие аллергических реакций, а именно: ринит, конъюнктивит, крапивница, раздражение в горле, слабость.
Лекарственное взаимодействие:
Глицин снижает токсичность антиконвульсантов, антипсихотических средств, антидепрессантов, противосудорожных средств. При сочетании со снотворными, транквилизаторами и антипсихотическими средствами усиливается эффект торможения центральной нервной системы.
Срок годности:
3 года. Не применять после истечения срока годности.
Не применять после истечения срока годности.
Условия хранения:
Хранить в оригинальной упаковке в недоступном для детей месте при температуре не выше 25 ° С.
Форма выпуска / упаковка:
По 50 таблеток сублингвальный в блистере. По 1 блистера в картонной коробке.
Категория отпуска:
Без рецепта.
Дополнительные данные
Дозировка:
таблетки сублингв. по 100 мг №50
Регистрация:
UA/14604/01/01 от 01.09.2015
Производитель:
ООО Арпимед, Республика Армения, Котайкська обл., г. Абовян, 2-й микрорайон, д. 19.
Код АТХ N07XX:
Внимание
Приведенная научная информация является обобщающей, основана на официально утвержденной инструкции по применению и не может быть использована для принятия решения о возможности применения конкретного лекарственного препарата.
Редакция
Автор: Жарікова Юлія
Жарікова Юлія
Лікар-педіатр
неонатолог
Донецький національний медичний університет ім. М. Горького, педіатричниий факультет
Реклама
Реклама
Реклама
Реклама
Экзогенное применение бетаина глицина поддерживает биоактивные соединения, антиоксидантную активность и физико-химические свойства плодов красного апельсина при длительном хранении в холодильнике
Графический реферат .
Введение
Плоды красного апельсина ( Citrus sinensis L. Osbeck) являются богатым источником биологически активных соединений по сравнению с другими видами цитрусовых, поскольку они могут способствовать синтезу антоциановых пигментов (1). Концентрация антоцианов может определять внутреннее качество плодов красного апельсина и повышать антиоксидантную способность, что очень полезно для здоровья человека, предотвращая некоторые заболевания (2).
Концентрация антоцианов может определять внутреннее качество плодов красного апельсина и повышать антиоксидантную способность, что очень полезно для здоровья человека, предотвращая некоторые заболевания (2).
В настоящее время увеличилась послеуборочная обработка элиситорными соединениями для сохранения качества плодов при хранении в холодильнике (1). Например, глицин-бетаин (ГБ) представляет собой соединение четвертичного аммония и может играть жизненно важную роль в осмотическом давлении клеток, стабилизации белка и макромолекул, а также функции фермента для повышения устойчивости к холоду у высших растений (3, 4). В последнее время проводят послеуборочную обработку ГБ плодоовощных культур, в том числе перца сладкого (3), мушмулы (4), персика (5–7), кабачков (8), боярышника (9).), а также груша (10–12), гранат (13) и банан (14).
Хранение в холодильнике является наилучшей стратегией для увеличения послеуборочной жизни плодов красного апельсина на несколько месяцев и обеспечивает присутствие этой культуры на рынке далеко от сезона сбора урожая (15). Однако восприимчивость плодов красного апельсина к низким температурам может ограничивать длительное хранение в холодильнике (16). Напротив, хранение красных апельсинов при температуре окружающей среды увеличивает потерю веса, старение и грибковые заболевания, а процесс порчи впоследствии сокращает срок хранения (1). Низкая температура может снизить физиологический метаболизм и процесс старения, повысить лежкость и сохранить биологически активные соединения плодов красного апельсина при длительном хранении. Длительное хранение красных апельсинов может привести к снижению качества, влияя на вкус фруктов и некоторые биологически активные соединения (2). В этом смысле существует потребность в новых методах, которые имеют решающее значение для увеличения срока хранения и сохранения качества плодов красного апельсина при хранении в холодильнике.
Однако восприимчивость плодов красного апельсина к низким температурам может ограничивать длительное хранение в холодильнике (16). Напротив, хранение красных апельсинов при температуре окружающей среды увеличивает потерю веса, старение и грибковые заболевания, а процесс порчи впоследствии сокращает срок хранения (1). Низкая температура может снизить физиологический метаболизм и процесс старения, повысить лежкость и сохранить биологически активные соединения плодов красного апельсина при длительном хранении. Длительное хранение красных апельсинов может привести к снижению качества, влияя на вкус фруктов и некоторые биологически активные соединения (2). В этом смысле существует потребность в новых методах, которые имеют решающее значение для увеличения срока хранения и сохранения качества плодов красного апельсина при хранении в холодильнике.
В последние годы послеуборочное применение элиситоров применялось для повышения лежкости и сохранения биологически активных соединений плодов красного апельсина при длительном хранении в холодильнике (17). Впервые оценено влияние послеуборочной обработки ГБ на сохранение внутренних качеств плодов красного апельсина при хранении в холодильнике. Таким образом, целью данного исследования было оценить влияние послеуборочной обработки ГБ на биологически активные соединения, антиоксидантную активность и физико-химические свойства плодов красного апельсина «Моро» при хранении в холодильнике.
Впервые оценено влияние послеуборочной обработки ГБ на сохранение внутренних качеств плодов красного апельсина при хранении в холодильнике. Таким образом, целью данного исследования было оценить влияние послеуборочной обработки ГБ на биологически активные соединения, антиоксидантную активность и физико-химические свойства плодов красного апельсина «Моро» при хранении в холодильнике.
Материалы и методы
Обработка плодов и условия хранения
Плоды красного апельсина ( Citrus sinensis L. Osbeck cv. Moro) собирали на стадии товарной зрелости из коммерческого цитрусового сада в Джахроме, провинция Фарс, Иран, и сразу транспортируют в послеуборочную лабораторию. После сбора плоды проверяли на однородность размера и отсутствие повреждений кожуры и дезинфицировали погружением в 2% раствор гипохлорита натрия (NaOCl). После высушивания воды плоды обрабатывали водным раствором ГБ концентрацией 15 и 30 мМ методом вакуумной инфильтрации при 30 кПа в течение 8 мин. Контрольные образцы не подвергались никакой обработке. Обработку проводили в трех повторностях по пять плодов. Все образцы хранились в течение 90 дней при 3°C и относительной влажности 90%. Продолжительность хранения и температура были выбраны на основе нашей предыдущей работы (18). Следующие параметры оценивали через 1, 30, 60 и 90 дней хранения в холодильнике плюс 2 дня при 20°C для определения срока годности.
Контрольные образцы не подвергались никакой обработке. Обработку проводили в трех повторностях по пять плодов. Все образцы хранились в течение 90 дней при 3°C и относительной влажности 90%. Продолжительность хранения и температура были выбраны на основе нашей предыдущей работы (18). Следующие параметры оценивали через 1, 30, 60 и 90 дней хранения в холодильнике плюс 2 дня при 20°C для определения срока годности.
Потеря веса и твердость
Процент потери веса плодов (WL) определяли путем взвешивания исходного веса (IW) перед хранением и конечного веса (FW) для каждого времени отбора проб (19).
Твердость плода измеряли с помощью анализатора текстуры (TA-XT2, Великобритания) путем сжатия 10% экваториальной площади и выражали в единицах Ньютона (2).
Химические характеристики сока
Общее количество растворимых сухих веществ (TSS) и титруемую кислотность (TA) определяли с помощью рефрактометра и метода титрования соответственно, а отношение TSS к TA определяли путем их деления (20).
Биоактивные соединения и антиоксидантная активность
Общая концентрация антоцианов (ТАС) измерялась спектрофотометрически. Фруктовый сок разбавляли (1:4) буфером хлорида калия (KCl) (pH 1,0) и ацетатом натрия (C 2 H 9).0039 3 NaO 2 ) буфера (pH 4,5), оптическую плотность определяли при 510 и 700 нм, а TAC сообщали как мг л -1 (20).
Общее содержание фенолов (TPC) измеряли по методу Folin-Ciocalteu (21). Вкратце, 700 мкл сока смешивали с 900 мкл 2% карбоната натрия (Na 2 CO 3 ), а затем после 3-минутной реакции добавляли 180 мкл 50% фолина. Образцы считывали при 750 нм через 30 мин выдерживания в темном месте при температуре окружающей среды. TPC рассчитывали, используя стандартную кривую, полученную для различных концентраций галловой кислоты, и выражали в мг эквивалента галловой кислоты (GAE) L −1 .
Общая антиоксидантная активность (ТАА) измерялась методом 2,2-дифенил-1-пикрилгидразила (DPPH) (22). Вкратце, 100 мкл фруктового сока смешивали с 1 мл DPPH (0,1 мМ) и 1 мл Tris-HCl (pH = 7,5) буфера, и считывали поглощение при 517 нм через 30 минут выдерживания при температуре окружающей среды и сообщали в процентах.
Вкратце, 100 мкл фруктового сока смешивали с 1 мл DPPH (0,1 мМ) и 1 мл Tris-HCl (pH = 7,5) буфера, и считывали поглощение при 517 нм через 30 минут выдерживания при температуре окружающей среды и сообщали в процентах.
Индивидуальные антоцианы
Индивидуальные антоцианы были количественно определены с помощью высокоэффективной жидкостной хроматографии (ВЭЖХ) на основе Martinez-Esplá et al. (23) метод. Для приготовления смеси использовали 0,5 г лиофилизированной целлюлозы, как описано ранее (2). Колонка для ВЭЖХ и подвижные фазы (муравьиная кислота и ацетонитрил) использовались ранее. Основные антоцианы [цианидин-3-глюкозид и цианидин-3-(6″-малонилглюкозид)] были обнаружены при 520 нм и количественно определены по стандартным кривым и выражены в мг л −1 .
Отдельные сахара и органические кислоты
Отдельные сахара и органические кислоты были определены Martinez-Esplá et al. (23) метод. Для количественного определения использовали 0,5 г лиофилизированной мякоти красного апельсина и готовили смесь, как описано ранее (2). Отдельные органические кислоты (лимонная кислота, аскорбиновая кислота, яблочная кислота, щавелевая кислота и янтарная кислота) и отдельные сахара (сахароза, глюкоза и фруктоза) выражали как г кг -1 с использованием стандартной кривой.
Отдельные органические кислоты (лимонная кислота, аскорбиновая кислота, яблочная кислота, щавелевая кислота и янтарная кислота) и отдельные сахара (сахароза, глюкоза и фруктоза) выражали как г кг -1 с использованием стандартной кривой.
Анализ активности ферментов
Активность ферментов определяли спектрофотометрически в мясе. Сначала 500 мг мякоти плодов гомогенизировали экстракционным буфером (фосфат калия, рН=7). После центрифугирования супернатант разделяли на активность каталазы (CAT), пероксидазы (POD), аскорбатпероксидазы (APX) и супероксиддисмутазы (SOD) и содержание общего белка. Активность CAT и POD оценивали по методу Chance and Maehly (24). Активность КАТ оценивали по разложению перекиси водорода (h3O2) по уменьшению поглощения при 240 нм (24). Активность POD оценивали с использованием гваякола в качестве субстрата по поглощению при 470 нм (24). Активность APX определяли путем измерения оптической плотности при 290 нм как количество фермента, окислившего аскорбат в минуту (25). Активность СОД определяли в реакционной смеси, содержащей 100 мкл неочищенного экстракта фермента при 560 нм, и рассчитывали как количество фермента, вызывающее 50%-ное восстановление нитросинего тетразолия (НСТ) (26).
Активность СОД определяли в реакционной смеси, содержащей 100 мкл неочищенного экстракта фермента при 560 нм, и рассчитывали как количество фермента, вызывающее 50%-ное восстановление нитросинего тетразолия (НСТ) (26).
Для определения активности PAL, кратко, 500 мг мякоти плодов гомогенизировали с буфером на основе бората натрия (pH = 7). Затем 0,5 мл неочищенного ферментного экстракта, натрий-боратного буфера (рН 8,8) и L-фенилаланина и инкубируют при 37°С в течение 60 мин. Активность ФАЛ измеряли при 290 нм (27).
Для экстракции ферментом ПФО 200 мг мякоти плодов гомогенизировали в калий-фосфатном буфере (pH = 7,8). Реакционную смесь, содержащую 100 мкл неочищенного ферментного экстракта, калий-фосфатный буфер (рН = 7) и раствор пирокатехина, инкубировали при 25°С в течение 10 мин. Затем определяли активность фермента ПФО при 425 нм (28). Общее содержание белка в экстракте фермента определяли при 595 нм (29), а удельную активность фермента сообщали как U мг -1 белка.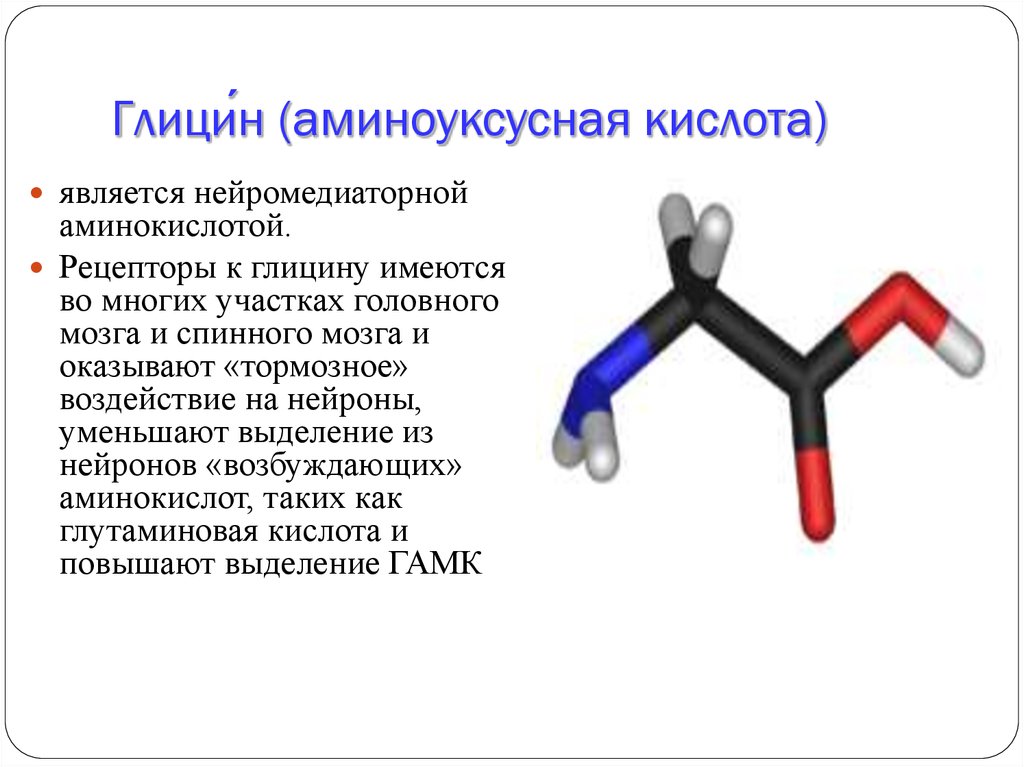
Статистический анализ
Это исследование планировалось как факторное на основе полностью рандомизированного дизайна (CRD) с тремя повторами. Факторами были лечение и время отбора проб. Анализ данных, сравнение средних значений (LSD) и стандартные ошибки (SE) были выполнены с помощью программного обеспечения SAS (v. 9.4) при p < 0,05.
Результаты
Потеря веса и упругость
Увеличение потери веса (WL) при хранении в холодильнике. В этом смысле у контрольных плодов значительно выше WL, чем у плодов, обработанных ГБ (рис. 1). Обработка GB облегчала WL, и самый низкий WL был при применении 30 мМ GB. Напротив, WL в контрольных плодах был значительно выше по сравнению с фруктами, обработанными ГБ. Фактически, во время последнего отбора проб WL плодов, обработанных ГБ при 15 и 30 мМ, составлял 19.на 0,64 и 36,55% ниже, чем у контрольных плодов соответственно.
Рисунок 1 . Изменения потери веса и твердости плодов в контроле и обработанных ГБ плодах красного апельсина «Моро» при 3°C. Различные буквы и вертикальные полосы показывают значительную разницу ( p <0,05) и стандартные ошибки (±SE) средних значений соответственно.
Различные буквы и вертикальные полосы показывают значительную разницу ( p <0,05) и стандартные ошибки (±SE) средних значений соответственно.
Повышенная потеря твердости при хранении в холодильнике. Обработка ГБ поддерживала значительно более высокую твердость плодов красного апельсина во время хранения красного апельсина «Моро» (рис. 1). В целом обработка 30 мМ ГБ была эффективной дозой для сохранения твердости плодов при хранении в холодильнике. В конце хранения твердость плодов, обработанных ГБ при 15 и 30 мМ, была на 18,64 и 30,66% выше, чем у контрольных образцов.
Химические характеристики сока
TSS слегка увеличивался во всех образцах до 30 дней, а затем оставался постоянным или снижался при хранении в холодильнике (рис. 2), при этом самый высокий и самый низкий TSS наблюдался в 30 мМ ГБ и контрольных образцах соответственно. .
Рисунок 2 . Изменения общего количества растворимых сухих веществ (TSS), титруемой кислотности (TA) и TSS/TA в контрольных и обработанных ГБ плодах красного апельсина «Моро» при 3°C. Различные буквы и вертикальные полосы показывают значительную разницу ( p < 0,05) и стандартных ошибок (±SE) средних соответственно.
Различные буквы и вертикальные полосы показывают значительную разницу ( p < 0,05) и стандартных ошибок (±SE) средних соответственно.
ТА уменьшилась в контроле и при лечении ГБ во время хранения в холодильнике (рис. 2). Однако контрольные образцы имели самую низкую ТА в конце холодового хранения. Обработка ГБ снижала потери ТА, особенно при концентрации 30 мМ. После 90 дней хранения в холодильнике ТА плодов, обработанных 15 и 30 мМ ГБ, была на 15,89 и 23,29% выше, чем у контрольных плодов соответственно.
На соотношение TSS/TA влияли все обработки, применяемые при хранении в холодильнике (рис. 2). Плоды, обработанные GB, имели самое низкое соотношение TSS/TA во время последнего отбора проб. В контрольных образцах TSS/TA резко увеличивалась при хранении на холоде. В целом, 30 мМ ГБ были наиболее эффективной дозой для сохранения соотношения TSS/TA во время хранения в холодильнике.
Отдельные сахара и органические кислоты
Отдельные сахара подверглись воздействию послеуборочной обработки ГБ во время хранения (рис. 3), в которой сахароза была обнаружена в более высокой концентрации, чем глюкоза и фруктоза. Сахароза, глюкоза и фруктоза увеличивались до 30 дней во всех обработках, а затем снижались к концу хранения в холодильнике. Снижение содержания отдельных сахаров в фруктах, обработанных ГБ, было отсрочено по сравнению с контрольными фруктами после 90 дней хранения в холодильнике. Например, самые высокие обнаруженные уровни сахарозы, глюкозы и фруктозы были в фруктах, обработанных 30 мМ ГБ.
3), в которой сахароза была обнаружена в более высокой концентрации, чем глюкоза и фруктоза. Сахароза, глюкоза и фруктоза увеличивались до 30 дней во всех обработках, а затем снижались к концу хранения в холодильнике. Снижение содержания отдельных сахаров в фруктах, обработанных ГБ, было отсрочено по сравнению с контрольными фруктами после 90 дней хранения в холодильнике. Например, самые высокие обнаруженные уровни сахарозы, глюкозы и фруктозы были в фруктах, обработанных 30 мМ ГБ.
Рисунок 3 . Изменения содержания сахарозы, глюкозы и фруктозы в контрольных и обработанных ГБ плодах красного апельсина «Моро» при 3°C. Различные буквы и вертикальные полосы показывают значительную разницу ( p <0,05) и стандартные ошибки (±SE) средних значений соответственно.
Содержание лимонной, яблочной, янтарной и щавелевой кислот значительно снизилось при хранении в холодильнике как в контрольных, так и в обработанных плодах (рис. 4). Плоды, обработанные ГБ, имели самое высокое содержание лимонной, яблочной, янтарной и щавелевой кислот, особенно для 30 мМ ГБ в течение 90 суток хранения, тогда как в контрольных образцах резко снижается. По окончании холодового хранения содержание лимонной, яблочной, янтарной и щавелевой кислот в плодах, обработанных 30 мМ ГБ, превышало контрольные образцы на 24,2, 30,12, 29,33 и 23,93 % соответственно.
По окончании холодового хранения содержание лимонной, яблочной, янтарной и щавелевой кислот в плодах, обработанных 30 мМ ГБ, превышало контрольные образцы на 24,2, 30,12, 29,33 и 23,93 % соответственно.
Рисунок 4 . Изменения содержания лимонной, яблочной, янтарной и щавелевой кислот в контрольных и обработанных ГБ плодах красного апельсина «Моро» при 3°C. Различные буквы и вертикальные полосы показывают значительную разницу ( p < 0,05) и стандартных ошибок (±SE) средних соответственно.
Биоактивные соединения и антиоксидантная активность
Общая концентрация антоцианов (ОАЦ) увеличивалась при хранении в холодильнике и значительно повышалась при обработке ГБ (рис. 5). Наибольшее содержание ТАС наблюдалось в конце срока хранения в плодах, обработанных ГБ. Во время последнего отбора проб TAC в плодах, обработанных 15 и 30 мМ ГБ, был на 25,23 и 45,01% выше, чем в необработанных плодах, соответственно. В целом, 30 мМ GB был наиболее эффективным средством для повышения TAC во время холодного хранения.
Рисунок 5 . Изменения общей концентрации антоцианов (ТАС), цианидин-3-гликозида, цианидин-3-(6″-малонилглюкозида), общего содержания фенолов, общей антиоксидантной активности (ТАА) и аскорбиновой кислоты в контрольной и обработанной ГБ крови Моро оранжевый при 3°С. Различные буквы и вертикальные столбцы показывают значительную разницу ( p <0,05) и стандартные ошибки (±SE) средних значений соответственно.
Основные отдельные антоцианы, в том числе цианидин-3-глюкозид и цианидин-3-(6″-малонилглюкозид), подверглись воздействию обработки во время хранения в холодильнике (рис. 5). Тенденции основных индивидуальных антоцианов были аналогичны изменениям TAC. Содержание цианидин-3-глюкозида и цианидин-3-(6″-малонилглюкозида) было выше, чем в плодах, обработанных 30 мМ ГБ, при хранении в холодильнике. В конце холодного хранения цианидин-3-глюкозид и цианидин-3-(6″-малонилглюкозид) в плодах, обработанных 30 мМ ГБ, были на 56,35 и 41,08% выше, чем в контрольных образцах, соответственно.
Общее содержание фенолов (TPC) увеличивалось до 60 дней хранения во всех вариантах обработки, а затем оставалось постоянным в обработанных ГБ или уменьшалось в контрольных образцах (рис. 5). Контрольные плоды показали самую низкую ТФХ при хранении в холодильнике. Обработка 30 мМ GB повышала TPC в плодах красного апельсина во время хранения в холодильнике. После 90 дней хранения в холодильнике ТФХ в плодах, обработанных 15 и 30 мМ ГБ, увеличилась на 37,73 и 47,86% по сравнению с контрольными образцами соответственно.
Общая антиоксидантная активность (ТАА) увеличивалась при хранении в холодильнике и на нее влияла обработка ГБ (рис. 5). ТАА повышали до 60 суток хранения и затем снижали к концу хранения. Самый высокий и самый низкий уровень TAA наблюдался у плодов, обработанных 30 мМ ГБ, и у контрольных образцов соответственно.
Количество аскорбиновой кислоты (АК) уменьшилось при хранении в холодильнике (рис. 5). У контрольных фруктов было значительно более высокое снижение содержания АК, чем у плодов, обработанных ГБ, при хранении в холодильнике. Обработка ГБ замедляла снижение содержания АК в обработанных плодах, а самый высокий уровень АК был обнаружен при 30 мМ ГБ. В конце хранения содержание АК в плодах, обработанных ГБ, в концентрациях 15 и 30 мМ было на 29,07 и 41,59% выше, чем в контрольных плодах соответственно.
Обработка ГБ замедляла снижение содержания АК в обработанных плодах, а самый высокий уровень АК был обнаружен при 30 мМ ГБ. В конце хранения содержание АК в плодах, обработанных ГБ, в концентрациях 15 и 30 мМ было на 29,07 и 41,59% выше, чем в контрольных плодах соответственно.
Активность антиоксидантных ферментов
Активность POD не была обнаружена в мякоти плодов при всех обработках в течение всего времени отбора проб.
Активность КАТ увеличивалась до 60 дней хранения в холодильнике, а затем снижалась к концу эксперимента (рис. 6). Послеуборочная обработка ГБ значительно повышала активность КАТ в плодах. Наименьшая активность КАТ была обнаружена в контрольных образцах. В конце холодового хранения активность КАТ у плодов, обработанных 15 и 30 мМ ГБ, была на 13,53 и 23,32% выше, чем у контрольных плодов соответственно.
Рисунок 6 . Изменения активности каталазы (КАТ), аскорбатпероксидазы (АПХ) и супероксиддисмутазы (СОД) в контроле и ГБ у красного апельсина «Моро» при 3°С. Различные буквы и вертикальные полосы показывают значительную разницу ( p < 0,05) и стандартных ошибок (±SE) средних соответственно.
Различные буквы и вертикальные полосы показывают значительную разницу ( p < 0,05) и стандартных ошибок (±SE) средних соответственно.
Активность APX была значительно повышена при обработке ГБ по сравнению с контрольными плодами при хранении в холодильнике (рис. 6). Активность APX увеличивалась до 60 дней хранения, а затем снижалась во всех обработках. 30 мМ GB был наиболее эффективным средством для повышения активности APX во время хранения в холодильнике.
На активность СОД также влияли обработки ГБ во время хранения (рис. 6). Активность СОД увеличивалась до 60 сут хранения, а затем снижалась в контрольных плодах. В 9На 0 сут холодового хранения активность СОД в плодах, обработанных 15 и 30 мМ ГБ, была выше (38,39 и 47,28% соответственно), чем в контроле.
Активность PAL и PPO
Активность PAL значительно увеличилась после обработки ГБ во время хранения в холодильнике (рис. 7). Активность ФАЛ резко увеличивалась в плодах, обработанных ГБ, до 60 дней хранения, а затем оставалась постоянной. Однако активность PAL снизилась в контрольных плодах после 60 дней хранения в холодильнике. Самая высокая и самая низкая активность PAL наблюдалась в 30 мМ ГБ и контрольных плодах соответственно. В конце хранения активность PAL в плодах, обработанных 15 и 30 мМ ГБ, составила 33,59.и на 43,75% больше, чем в контрольных плодах соответственно.
Однако активность PAL снизилась в контрольных плодах после 60 дней хранения в холодильнике. Самая высокая и самая низкая активность PAL наблюдалась в 30 мМ ГБ и контрольных плодах соответственно. В конце хранения активность PAL в плодах, обработанных 15 и 30 мМ ГБ, составила 33,59.и на 43,75% больше, чем в контрольных плодах соответственно.
Рисунок 7 . Изменения активности фенилаланин-аммиак-лиазы (ФАЛ) и полифенолоксидазы (ПФО) в контроле и ГБ у красного апельсина «Моро» при 3°С. Различные буквы и вертикальные столбцы показывают значительную разницу ( p <0,05) и стандартные ошибки (±SE) средних значений соответственно.
Активность PPO увеличивалась до 60 дней хранения в холодильнике и постоянно снижалась до конца хранения (рис. 7). Наибольшая активность ПФО наблюдалась во флаведо контрольных образцов. Обработка ГБ подавляла активность ПФО при хранении в холодильнике, и самая низкая активность ПФО наблюдалась во флаведо обработанных плодов с 30 мМ ГБ. После 9Через 0 дней хранения активность ПФО в плодах, обработанных 15 и 30 мМ ГБ, была на 15,21 и 35,86% ниже, чем в контрольных плодах соответственно.
После 9Через 0 дней хранения активность ПФО в плодах, обработанных 15 и 30 мМ ГБ, была на 15,21 и 35,86% ниже, чем в контрольных плодах соответственно.
Обсуждение
Потребление красных апельсинов приобрело повышенное признание и популярность среди потребителей из-за содержания в них антоцианинового пигмента. Длительное хранение в холодильнике увеличивает срок хранения плодов красного апельсина после сбора урожая и продлевает присутствие этого товара на рынке далеко от сезона сбора урожая (30). Однако длительное хранение приводит к потерям органических кислот, антиоксидантной активности и биоактивных соединений и впоследствии влияет на качественные характеристики плодов красных апельсинов. Таким образом, перед производителями и фруктовой отраслью стоит задача решить эту проблему. Послеуборочная обработка элиситорами может быть полезным подходом для повышения сохранности красных апельсинов при низких температурах (17). В последние годы послеуборочная обработка элиситорными соединениями сохраняла антиоксидантную активность и биологически активные соединения плодов красного апельсина при хранении в холодильнике (2, 17, 20). Многочисленные соединения, в том числе полиамины (путресцин, пут) (20), γ-аминомасляная кислота (ГАМК) (2), метилжасмонат (MeJA) (2), метилсалицилат (MeSA) (2) и 24-эпибрассинолид (24- EBR) (17), применяли после сбора урожая на красных апельсинах. Эти соединения классифицируются как алифатические поликатионы (Пут), аминокислоты (ГАМК) и растительные гормоны (МеЖА, МеСК и 24-ЭБР). Нанесение элиситоров осуществляли методами вакуумной инфильтрации (2, 20), обработки паром (2) или погружением (17). Выбор методов лечения зависит от соединений и средств. Однако обработка паром и метод погружения просты по сравнению с вакуумной инфильтрацией (30). Кроме того, применяемый метод зависит от природы вещества. Например, возможно использование MeJA и MeSA путем обработки паром из-за их летучести (2). Кроме того, некоторые соединения можно наносить с помощью вакуума или погружением в водный раствор. Хотя метод погружения прост и быстр, вакуумная инфильтрация более эффективна и обеспечивает оптимальное проникновение малых доз в плод (20).
Многочисленные соединения, в том числе полиамины (путресцин, пут) (20), γ-аминомасляная кислота (ГАМК) (2), метилжасмонат (MeJA) (2), метилсалицилат (MeSA) (2) и 24-эпибрассинолид (24- EBR) (17), применяли после сбора урожая на красных апельсинах. Эти соединения классифицируются как алифатические поликатионы (Пут), аминокислоты (ГАМК) и растительные гормоны (МеЖА, МеСК и 24-ЭБР). Нанесение элиситоров осуществляли методами вакуумной инфильтрации (2, 20), обработки паром (2) или погружением (17). Выбор методов лечения зависит от соединений и средств. Однако обработка паром и метод погружения просты по сравнению с вакуумной инфильтрацией (30). Кроме того, применяемый метод зависит от природы вещества. Например, возможно использование MeJA и MeSA путем обработки паром из-за их летучести (2). Кроме того, некоторые соединения можно наносить с помощью вакуума или погружением в водный раствор. Хотя метод погружения прост и быстр, вакуумная инфильтрация более эффективна и обеспечивает оптимальное проникновение малых доз в плод (20). В данном исследовании мы впервые применили послеуборочную обработку ГБ для оценки ее влияния на биоактивные соединения, физико-химические показатели, антиоксидантную активность и качество плодов красного апельсина «Моро» при хранении при 3°С.
В данном исследовании мы впервые применили послеуборочную обработку ГБ для оценки ее влияния на биоактивные соединения, физико-химические показатели, антиоксидантную активность и качество плодов красного апельсина «Моро» при хранении при 3°С.
Обработка GB снижала WL по сравнению с контрольными образцами при хранении в холодильнике. WL плода является результатом диффузии воды из кожуры. Вода является основным компонентом кожуры цитрусовых, и поддержание влажности кожуры может сохранить вес плода при хранении в холодильнике. Потеря влаги в основном происходит из-за диффузии воды через поверхность кожуры и, впоследствии, попадание в WL плода (19). С другой стороны, WL обусловлен дыханием, процессом транспирации и уменьшением естественного воска фруктов во время хранения в холодильнике. Испарение можно уменьшить с помощью натурального фруктового воска (31). В этом исследовании 30 мМ ГБ были эффективны для облегчения WL во время холодного хранения, вероятно, из-за сохранения целостности мембраны при низких температурах (18). Точно так же вакуумная инфильтрация ГАМК снижала WL в плодах красного апельсина. В этом смысле вакуумная инфильтрация более эффективна, чем методы погружения (20). Кроме того, кажется, что аминокислоты, применяемые экзогенно, могут поддерживать влажность фруктов, сохраняя структуру эпидермиса кожуры (2).
Точно так же вакуумная инфильтрация ГАМК снижала WL в плодах красного апельсина. В этом смысле вакуумная инфильтрация более эффективна, чем методы погружения (20). Кроме того, кажется, что аминокислоты, применяемые экзогенно, могут поддерживать влажность фруктов, сохраняя структуру эпидермиса кожуры (2).
Плотность плодов красного апельсина является важным фактором приемлемости на рынке (19). В этом исследовании фрукты, обработанные ГБ, имели меньшую потерю твердости, чем контрольные фрукты, при хранении в холодильнике. Пектиновые соединения представляют собой структурные полисахариды, поддерживающие упругость плодов. Механизм потери плотности обусловлен деполимеризацией пектина ферментами полигалактуроназой, пектинлиазой, пектинметилэстеразой и целлюлазой, которые могут вызывать некоторые изменения в составе клеточной стенки и происходить размягчение (31). В этом исследовании лечение ГБ, вероятно, влияло на активность ферментов, разрушающих клеточную стенку, из-за поддержания стабильности и прочности мембраны. Вакуумная инфильтрация путресцином сортов красного апельсина для поддержания плотности показала, что обработанные плоды были более плотными, чем контрольные образцы, благодаря связыванию с нуклеиновыми кислотами, фосфолипидами и карбоксильными группами полигалактуроновой кислоты. Интересно, что этот ответ зависел от концентрации и сорта (20).
Вакуумная инфильтрация путресцином сортов красного апельсина для поддержания плотности показала, что обработанные плоды были более плотными, чем контрольные образцы, благодаря связыванию с нуклеиновыми кислотами, фосфолипидами и карбоксильными группами полигалактуроновой кислоты. Интересно, что этот ответ зависел от концентрации и сорта (20).
Сахароза, глюкоза и фруктоза являются основными компонентами TSS (примерно 80%) цитрусовых, а другие компоненты представляют собой органические кислоты (10%), свободные аминокислоты, глюкозиды, белки и витамины (2). В этом исследовании фрукты, обработанные GB, имели самый высокий TSS. Увеличение TSS, вероятно, связано с превращением органических кислот в сахара в результате ферментативного процесса при хранении в холодильнике (1).
В нашем исследовании основными отдельными сахарами были сахароза, глюкоза и фруктоза. Все отдельные сахара повышались до 30 дней, а затем снижались к концу холодового хранения. Первоначальное увеличение количества отдельных сахаров происходит за счет конверсии органических кислот. Снижение содержания отдельных сахаров, вероятно, было связано с потреблением сахара для получения энергии в процессе дыхания при длительном хранении (31). Кроме того, снижение содержания сахарозы в красных апельсинах является следствием явления гидролитического расщепления, которое в основном влияет на сахарозу и другие гликозилированные компоненты сока при хранении в холодильнике (17). Однако обработка ГБ поддерживала более высокую концентрацию сахарозы, глюкозы и фруктозы и оказывала положительное влияние на предотвращение снижения уровня сахара при хранении в холодильнике. В плодах груши обработка ГБ влияла на уровни сахарозы, глюкозы и фруктозы во время хранения в холодильнике, а контрольные образцы содержали больше глюкозы и фруктозы, чем фрукты, обработанные ГБ (12).
Снижение содержания отдельных сахаров, вероятно, было связано с потреблением сахара для получения энергии в процессе дыхания при длительном хранении (31). Кроме того, снижение содержания сахарозы в красных апельсинах является следствием явления гидролитического расщепления, которое в основном влияет на сахарозу и другие гликозилированные компоненты сока при хранении в холодильнике (17). Однако обработка ГБ поддерживала более высокую концентрацию сахарозы, глюкозы и фруктозы и оказывала положительное влияние на предотвращение снижения уровня сахара при хранении в холодильнике. В плодах груши обработка ГБ влияла на уровни сахарозы, глюкозы и фруктозы во время хранения в холодильнике, а контрольные образцы содержали больше глюкозы и фруктозы, чем фрукты, обработанные ГБ (12).
Титруемая кислотность (ТК) снижалась во время хранения, а плоды, обработанные ГБ, имели более высокую ТК в конце хранения. ТА коррелирует с содержанием органических кислот и важных компонентов для качественного цитрусового сока (20). В этом исследовании основными органическими кислотами были лимонная, яблочная, аскорбиновая, янтарная и щавелевая кислоты в плодах красного апельсина «Моро». Лимонная и яблочная кислоты были первой и второй основными органическими кислотами соответственно. Синтез сахаров из органических кислот, синтез фенольных соединений и производство энергии являются основными причинами снижения содержания органических кислот при хранении в холодильнике (1). В этом исследовании фрукты, обработанные 30 мМ ГБ, имели более высокое содержание органических кислот при хранении в холодильнике. Более высокое содержание органических кислот в фруктах, обработанных ГБ, вероятно, связано с влиянием на скорость дыхания за счет снижения активности гликолитических ферментов, таким образом поддерживая органические кислоты и замедляя старение, увеличивая срок хранения плодов (32).
В этом исследовании основными органическими кислотами были лимонная, яблочная, аскорбиновая, янтарная и щавелевая кислоты в плодах красного апельсина «Моро». Лимонная и яблочная кислоты были первой и второй основными органическими кислотами соответственно. Синтез сахаров из органических кислот, синтез фенольных соединений и производство энергии являются основными причинами снижения содержания органических кислот при хранении в холодильнике (1). В этом исследовании фрукты, обработанные 30 мМ ГБ, имели более высокое содержание органических кислот при хранении в холодильнике. Более высокое содержание органических кислот в фруктах, обработанных ГБ, вероятно, связано с влиянием на скорость дыхания за счет снижения активности гликолитических ферментов, таким образом поддерживая органические кислоты и замедляя старение, увеличивая срок хранения плодов (32).
В этом исследовании соотношение TSS/TA увеличивалось при хранении в холодильнике. Изменение содержания органических кислот может повлиять на это соотношение. С другой стороны, кислотность цитрусовых связана с уровнями органических кислот, а потеря органических кислот из-за преобразования может изменить соотношение TSS и TA (33). В этом исследовании плоды, обработанные 30 мМ ГБ, имели более низкое соотношение TSS/TA при хранении в холодильнике, вероятно, из-за наблюдаемого более высокого содержания органических кислот.
С другой стороны, кислотность цитрусовых связана с уровнями органических кислот, а потеря органических кислот из-за преобразования может изменить соотношение TSS и TA (33). В этом исследовании плоды, обработанные 30 мМ ГБ, имели более низкое соотношение TSS/TA при хранении в холодильнике, вероятно, из-за наблюдаемого более высокого содержания органических кислот.
После сбора плоды красного апельсина могут синтезировать антоцианы путем активации ферментов, участвующих в метаболизме фенилпропаноидов, в основном фермента PAL при низких температурах (34). В этом исследовании активность PAL повышалась при всех вариантах лечения, и 30 мМ ГБ были наиболее эффективным средством для повышения активности PAL. Это усиление, вероятно, связано с увеличением содержания АТФ и АДФ, которые могут потребляться для клеточного метаболизма (5). Кроме того, послеуборочная обработка ГБ повышала активность PAL в плодах боярышника, усиливая накопление антоцианов (9).). Кроме того, подобно TAC, цианидин-3-глюкозид и цианидин-3-(6″-малонилглюкозид) в качестве основных антоцианов в плодах красного апельсина были выше в фруктах, обработанных ГБ. Однако усиление TAC, цианидин-3-глюкозида и цианидин-3-(6″-малонилглюкозида) в контрольных образцах было ниже, чем в фруктах, обработанных ГБ, вероятно, из-за потери органических кислот или сахаров, и, следовательно, более низкого образования АТФ для обеспечения клеточной активности. энергия при холодовом стрессе (1, 5). Кроме того, на деградацию антоцианов может влиять активность PPO и POD (34). В этом исследовании активность POD не была обнаружена при всех вариантах лечения. Кроме того, активность ПФО была выше, чем у плодов, обработанных ГБ. Следовательно, активность ПФО приводит к деградации молекул антоцианов. Более высокая активность PPO и более низкая активность PAL могут объяснить, почему накопление антоцианов было ниже по сравнению с фруктами, обработанными ГБ. Однако активность ПФО увеличивалась во время хранения, вероятно, из-за повышения рН, снижения содержания органических кислот и задержки старения плодов и обработки ГБ (1). В этом смысле плоды граната, обработанные GB, имели больше TAC, чем контрольный образец при хранении в холодильнике (13).
Однако усиление TAC, цианидин-3-глюкозида и цианидин-3-(6″-малонилглюкозида) в контрольных образцах было ниже, чем в фруктах, обработанных ГБ, вероятно, из-за потери органических кислот или сахаров, и, следовательно, более низкого образования АТФ для обеспечения клеточной активности. энергия при холодовом стрессе (1, 5). Кроме того, на деградацию антоцианов может влиять активность PPO и POD (34). В этом исследовании активность POD не была обнаружена при всех вариантах лечения. Кроме того, активность ПФО была выше, чем у плодов, обработанных ГБ. Следовательно, активность ПФО приводит к деградации молекул антоцианов. Более высокая активность PPO и более низкая активность PAL могут объяснить, почему накопление антоцианов было ниже по сравнению с фруктами, обработанными ГБ. Однако активность ПФО увеличивалась во время хранения, вероятно, из-за повышения рН, снижения содержания органических кислот и задержки старения плодов и обработки ГБ (1). В этом смысле плоды граната, обработанные GB, имели больше TAC, чем контрольный образец при хранении в холодильнике (13). Кроме того, TAC и основные антоцианы увеличились в красных апельсинах с MeJA и MeSA больше, чем с обработкой GABA. MeJA и MeSA показали более высокий эффект при экзогенном применении по сравнению с аминокислотами (ГАМК). Похоже, что растительные гормоны могут оказывать более благоприятное воздействие, чем аминокислоты, благодаря своей природе и действию на физиологию плодов после сбора урожая (2). В этом исследовании TPC увеличился в обработанных плодах и уменьшился в контрольных образцах в конце хранения. Увеличение фенольных соединений связано с активностью PAL и накоплением антоцианов, что привело к увеличению TPC (20). Это основная причина более высоких значений TPC в фруктах, обработанных ГБ. Снижение ТФХ в конце хранения в контрольных образцах, вероятно, связано с ферментативным окислением или деградацией под действием ферментов POD и PPO (2). Одновременно с результатами активность PAL увеличивалась при обработке ГБ и увеличивалась TPC в плодах груши, хранящихся в холодильнике (12).
Кроме того, TAC и основные антоцианы увеличились в красных апельсинах с MeJA и MeSA больше, чем с обработкой GABA. MeJA и MeSA показали более высокий эффект при экзогенном применении по сравнению с аминокислотами (ГАМК). Похоже, что растительные гормоны могут оказывать более благоприятное воздействие, чем аминокислоты, благодаря своей природе и действию на физиологию плодов после сбора урожая (2). В этом исследовании TPC увеличился в обработанных плодах и уменьшился в контрольных образцах в конце хранения. Увеличение фенольных соединений связано с активностью PAL и накоплением антоцианов, что привело к увеличению TPC (20). Это основная причина более высоких значений TPC в фруктах, обработанных ГБ. Снижение ТФХ в конце хранения в контрольных образцах, вероятно, связано с ферментативным окислением или деградацией под действием ферментов POD и PPO (2). Одновременно с результатами активность PAL увеличивалась при обработке ГБ и увеличивалась TPC в плодах груши, хранящихся в холодильнике (12).
TAA увеличилось, а затем уменьшилось до конца памяти. Фрукты в условиях холодового стресса накапливают активные формы кислорода (АФК) и нуждаются в очистке с помощью ферментативных и неферментативных антиоксидантных систем (35). В этом исследовании CAT, APX и SOD увеличивались до 60 дней хранения в холодильнике, а затем снижались в результате перепроизводства АФК. Кроме того, аскорбиновая кислота и фенольные соединения могут помочь в нейтрализации АФК в качестве неферментативных антиоксидантов (31). В этом исследовании снижение содержания аскорбиновой кислоты, вероятно, было связано с удалением АФК как донора электронов, который может нейтрализовать свободные радикалы при субоптимальных температурах (2). Кроме того, активность аскорбатоксидазы и POD может разрушать аскорбиновую кислоту (1). У груши активность APX, CAT и SOD также быстро увеличивалась в фруктах, обработанных ГБ, во время холодного хранения (10).
Выводы
Впервые сообщалось о влиянии послеуборочного применения ГБ на биологически активные соединения и антиоксидантную активность плодов красного апельсина при длительном хранении на холоде. Обработка ГБ замедляла потерю веса и упругости, а также накопление органических кислот и сахара. Кроме того, обработка GB улучшала биоактивные соединения и поддерживала качество за счет увеличения PAL и подавления активности PPO, что приводило к более высокому TAC и TPC, увеличивая TAA. Можно сделать вывод, что 30 мМ ГБ были наиболее эффективным средством и могут быть перспективной стратегией для сохранения физико-химической, биоактивной и антиоксидантной активности плодов красного апельсина сорта. Моро при 3°С.
Обработка ГБ замедляла потерю веса и упругости, а также накопление органических кислот и сахара. Кроме того, обработка GB улучшала биоактивные соединения и поддерживала качество за счет увеличения PAL и подавления активности PPO, что приводило к более высокому TAC и TPC, увеличивая TAA. Можно сделать вывод, что 30 мМ ГБ были наиболее эффективным средством и могут быть перспективной стратегией для сохранения физико-химической, биоактивной и антиоксидантной активности плодов красного апельсина сорта. Моро при 3°С.
Заявление о доступности данных
Первоначальные материалы, представленные в исследовании, включены в статью/дополнительный материал. Дальнейшие запросы можно направлять соответствующему автору.
Вклад авторов
FH и FG задумали и разработали эксперимент. FH провел исследования, аналитическое определение и анализ данных, а также написал первоначальный проект. Д.В., М.С. и Ф.Г. отредактировали первоначальный черновик. Все авторы внесли свой вклад в статью и одобрили представленную версию.
Финансирование
Эта рукопись принадлежит проекту Инновационные и экологически безопасные предуборочные и послеуборочные стратегии с использованием натуральных соединений для улучшения качества фруктов, финансируемому Conselleria d’Innovació, Universitats, Ciència i Societat Digital (Generalitat Valenciana) через программу Prometeo ( PROMETEO/2021/089).
Конфликт интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могли бы быть истолкованы как потенциальный конфликт интересов.
Примечание издателя
Все претензии, изложенные в этой статье, принадлежат исключительно авторам и не обязательно представляют претензии их дочерних организаций или издателя, редакторов и рецензентов. Любой продукт, который может быть оценен в этой статье, или претензии, которые могут быть сделаны его производителем, не гарантируются и не поддерживаются издателем.
Благодарности
Авторы выражают признательность Университету Шираза и Университету Мигеля Эрнандеса (UMH) за участие в исследованиях, приглашенных научным сотрудником FH.
Ссылки
1. Хабиби Ф., Рамезанян А., Гильен Ф., Кастильо С., Серрано М., Валеро Д. Изменения биологически активных соединений, антиоксидантной активности и питательного качества сортов красного апельсина при различных температурах хранения. Антиоксиданты. (2020) 9:1016. doi: 10.3390/antiox16
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
2. Хабиби Ф., Рамезанян А., Гильен Ф., Серрано М., Валеро Д. Кровавые апельсины сохраняют биологически активные соединения и питательные свойства благодаря послеуборочной обработке γ-аминомасляной кислотой, метилжасмонатом или метилсалицилатом во время хранения в холодильнике. Пищевая хим. (2020) 306:125634. doi: 10.1016/j.foodchem.2019.125634
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
3. Wang Q, Ding T, Zuo J, Gao L, Fan L. Улучшение последствий послеуборочного охлаждения сладкого перца с помощью глицина бетаина. Послеуборочная биотехнология. (2016) 112:114–20. doi: 10.1016/j.postharvbio.2015.07.008
(2016) 112:114–20. doi: 10.1016/j.postharvbio.2015.07.008
CrossRef Полный текст | Google Scholar
4. Zhang Y, Jin P, Huang Y, Shan T, Wang L, Li Y, et al. Воздействие горячей воды в сочетании с глицин-бетаином облегчает повреждение плодов мушмулы, хранящихся в холоде. Послеуборочная биотехнология. (2016) 118:141–7. doi: 10.1016/j.postharvbio.2016.04.010
CrossRef Полный текст | Google Scholar
5. Shan T, Jin P, Zhang Y, Huang Y, Wang X, Zheng Y. Экзогенная обработка глицин-бетаином повышает устойчивость плодов персика к холоду при хранении в холодильнике. Послеуборочная биотехнология. (2016) 114:104–10. doi: 10.1016/j.postharvbio.2015.12.005
CrossRef Полный текст | Google Scholar
6. Wang L, Bokhary SUF, Xie B, Hu S, Jin P, Zheng Y. Биохимические и молекулярные эффекты обработки глицин-бетаином на мембранный метаболизм жирных кислот в персиках, хранящихся в холодильнике. Послеуборочная биотехнология. (2019) 154:58–69. doi: 10.1016/j.postharvbio.2019.04.007
doi: 10.1016/j.postharvbio.2019.04.007
CrossRef Полный текст | Google Scholar
7. Ван Л.И., Шань Т., Се Б., Линг С., Шао С., Джин П. и др. Глицин-бетаин уменьшает повреждение плодов персика от переохлаждения за счет усиления метаболизма фенолов и сахаров. Пищевая хим. (2019) 272:530–8. doi: 10.1016/j.foodchem.2018.08.085
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
8. Yao W, Xu T, Farooq SU, Jin P, Zheng Y. Обработка глицином бетаином облегчает повреждение плодов цуккини (Cucurbita pepo L) вследствие переохлаждения путем модулирования антиоксидантных ферментов и мембранного метаболизма жирных кислот. Послеуборочная биотехнология. (2018) 144:20–8. doi: 10.1016/j.postharvbio.2018.05.007
Полный текст CrossRef | Google Scholar
9. Разави Ф., Махмуди Р., Рабией В., Агдам М.С., Сулеймани А. Обработка глицином-бетаином смягчает последствия переохлаждения и сохраняет питательные свойства плодов боярышника при хранении при низкой температуре. Научный Хортик. (2018) 233:188–94. doi: 10.1016/j.scienta.2018.01.053
Научный Хортик. (2018) 233:188–94. doi: 10.1016/j.scienta.2018.01.053
Полный текст CrossRef | Google Scholar
10. Luo M, Zhou X, Sun H, Zhou Q, Wei B, Cheng S, et al. Обработка глицином-бетаином снижает потерю эфиров, связанных с ароматом, в хранящихся в холодильнике грушах сорта «Нангуо», регулируя липоксигеназный путь. Пищевая хим. (2020) 316:126335. doi: 10.1016/j.foodchem.2020.126335
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
11. Sun H, Luo M, Zhou X, Zhou Q, Sun Y, Ge W и др. Экзогенная обработка глицин-бетаином смягчает вызванное низкой температурой потемнение околоплодника груш сорта «Нангуо» за счет регулирования антиоксидантных ферментов и метаболизма пролина. Пищевая хим. (2020) 306:125626. doi: 10.1016/j.foodchem.2019.125626
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
12. Wang J, Lv M, He H, Jiang Y, Yang J, Ji S. Бетаин с глицином облегчал потемнение кожуры хранящихся в холодильнике груш Nanguo в течение срока годности, регулируя метаболизм фенилпропаноида и растворимого сахара. Научный Хортик. (2020) 262:109100. doi: 10.1016/j.scienta.2019.109100
Научный Хортик. (2020) 262:109100. doi: 10.1016/j.scienta.2019.109100
Полный текст CrossRef | Google Scholar
13. Molaei S, Rabiei V, Soleimani A, Razavi F. Экзогенное применение глицина-бетаина повышает устойчивость плодов граната к холоду. malase Saveh при хранении в холодильнике. J Консервант для пищевых продуктов. (2021) 45:e15315. doi: 10.1111/jfpp.15315
Полный текст CrossRef | Google Scholar
14. Chen LL, Shan W, Cai DL, Chen JY, Lu WJ, Su XG и другие. Послеуборочное применение бетаина глицина уменьшает повреждение плодов банана, хранящихся в холодильнике, за счет усиления антиоксидантной системы. Научный Хортик. (2021) 287:110264. doi: 10.1016/j.scienta.2021.110264
Полный текст CrossRef | Google Scholar
15. Хабиби Ф., Рамезанян А., Гильен Ф., Мартинес-Ромеро Д., Серрано М., Валеро Д. Восприимчивость сортов красного апельсина к холодовому повреждению на основе антиоксидантной системы и физиологических и биохимических реакций при различных температурах хранения. Еда. (2020) 9:1609. doi: 10.3390/foods09
Еда. (2020) 9:1609. doi: 10.3390/foods09
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
16. Хабиби Ф., Рамезанян А., Гильен Ф., Серрано М., Валеро Д. Влияние различных послеуборочных обработок на летучие ароматические соединения плодов красного апельсина, подвергшихся воздействию температуры охлаждения после длительного хранения. Технология пищевых биопроцессов. (2020) 13:2054–64. doi: 10.1007/s11947-020-02547-1
Полный текст CrossRef | Google Scholar
17. Хабиби Ф., Серрано М., Закариас Л., Валеро Д., Гильен Ф. Послеуборочное применение 24-эпибрассинолида уменьшает симптомы обморожения и повышает содержание биоактивных соединений и антиоксидантную активность плодов красного апельсина. Передний завод Sci . (2021) 12:629733. doi: 10.3389/fpls.2021.629733
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
18. Хабиби Ф., Рамезанян А., Рахеми М., Эшги С., Гильен Ф., Серрано М. и др. Послеуборочная обработка γ-аминомасляной кислотой, метилжасмонатом или метилсалицилатом повышает устойчивость плодов красного апельсина к холоду при длительном хранении в холоде. J Sci Food Agric. (2019) 99: 6408–17. doi: 10.1002/jsfa.9920
и др. Послеуборочная обработка γ-аминомасляной кислотой, метилжасмонатом или метилсалицилатом повышает устойчивость плодов красного апельсина к холоду при длительном хранении в холоде. J Sci Food Agric. (2019) 99: 6408–17. doi: 10.1002/jsfa.9920
PubMed Abstract | Полный текст перекрестной ссылки | Академия Google
19. Хабиби Ф., Гильен Ф., Серрано М., Валеро Д. Физико-химические изменения, цвет кожуры и свойства сока сортов красного апельсина, хранящихся при разных температурах. Садоводство. (2021) 7:320. doi: 10.3390/horticulturae70
Полный текст CrossRef | Google Scholar
20. Habibi F, Ramezanian A. Вакуумная инфильтрация путресцина усиливает биоактивные соединения и сохраняет качество красного апельсина при хранении в холодильнике. Пищевая хим. (2017) 227:1–8. doi: 10.1016/j.foodchem.2017.01.057
Реферат PubMed | Полный текст перекрестной ссылки | Google Scholar
21. Мейерс К.Дж., Уоткинс С.Б., Приттс М. П., Лю Р.Х. Антиоксидантная и антипролиферативная активность клубники. J Agric Food Chem. (2003) 51:6887–92. doi: 10.1021/jf034506n
П., Лю Р.Х. Антиоксидантная и антипролиферативная активность клубники. J Agric Food Chem. (2003) 51:6887–92. doi: 10.1021/jf034506n
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
22. Brand-Williams W, Cuvelier ME, Berset C. Использование свободнорадикального метода для оценки антиоксидантной активности. LWT. (1995) 28:25–30. дои: 10.1016/S0023-6438(95)80008-5
Полнотекстовая перекрестная ссылка | Google Scholar
23. Martinez-Esplá A, Zapata PJ, Valero D, Garcia-Viguera C, Castillo S, Serrano M. Внесение щавелевой кислоты перед сбором урожая увеличивает размер плодов, биологически активные соединения и антиоксидантную способность сортов черешни (Prunus avium). л). J Agric Food Chem. (2014) 62:3432–7. doi: 10.1021/jf500224g
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
24. Chance B, Maehly A. Анализ каталаз и пероксидаз. Мет Энзимол . (1955) 2: 764–75. doi: 10. 1016/S0076-6879(55)02300-8
1016/S0076-6879(55)02300-8
Полный текст CrossRef | Google Scholar
25. Nakano Y, Asada K. Перекись водорода удаляется аскорбат-специфичной пероксидазой в хлоропластах шпината. Физиология клеток растений. (1981) 22:867–80.
Google Scholar
26. Beauchamp C, Fridovich I. Супероксиддисмутаза: улучшенные анализы и анализ, применимый к акриламидным гелям. Анальная биохимия. (1971) 44: 276–87. дои: 10.1016/0003-2697(71)
-8
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
27. Liu Q, Xi Z, Gao J, Meng Y, Lin S, Zhang Z. Влияние экзогенного 24-эпибрассинолида на борьбу с серой гнилью и поддержание послеуборочного качества столового винограда. Int J Food Sci. (2016) 51:1236–43. doi: 10.1111/ijfs.13066
Полный текст CrossRef | Google Scholar
28. Silva CRd, Koblitz MGB. Частичная характеристика и инактивация пероксидаз и полифенолоксидаз умбу-чая ( Spondias spp.). Пищевая наука . (2010) 30:790–6. doi: 10.1590/S0101-20612010000300035
(2010) 30:790–6. doi: 10.1590/S0101-20612010000300035
Полный текст CrossRef | Google Scholar
29. Брэдфорд М.М. Быстрый и чувствительный метод количественного определения белка в микрограммах, использующий принцип связывания белка с красителем. Анальная биохимия. (1976) 72: 248–54. doi: 10.1016/0003-2697(76)-3
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
30. Хабиби Ф., Гарсия-Пастор М.Э., Гильен Ф., Серрано М., Валеро Д. Состав жирных кислот в связи с восприимчивостью сортов красного апельсина к холоду при различных температурах хранения. Завод Физиол Биохим. (2021) 166: 770–6. doi: 10.1016/j.plaphy.2021.06.013
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
31. Валеро Д., Серрано М. Послеуборочная биология и технология сохранения качества фруктов : Бока-Ратон, Флорида: CRC Press (2010). doi: 10.1201/9781439802670
Полный текст CrossRef | Google Scholar
32. Валеро Д., Диас-Мула Х.М., Запата П.Дж., Кастильо С., Гильен Ф., Мартинес-Ромеро Д. и другие. Послеуборочная обработка салициловой кислотой, ацетилсалициловой кислотой или щавелевой кислотой задерживает созревание и увеличивает биоактивные соединения и антиоксидантную способность черешни. J Сельскохозяйственная пищевая химия. (2011) 59:5483–9. doi: 10.1021/jf200873j
Валеро Д., Диас-Мула Х.М., Запата П.Дж., Кастильо С., Гильен Ф., Мартинес-Ромеро Д. и другие. Послеуборочная обработка салициловой кислотой, ацетилсалициловой кислотой или щавелевой кислотой задерживает созревание и увеличивает биоактивные соединения и антиоксидантную способность черешни. J Сельскохозяйственная пищевая химия. (2011) 59:5483–9. doi: 10.1021/jf200873j
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
33. Ладо Дж., Родриго М.Дж., Закариас Л. Показатели зрелости и качество цитрусовых. Stewart Postharvest Rev. (2014) 10:1–6.
Google Scholar
34. Ло Пьеро А.Р. Современное состояние биосинтеза антоцианов и его регуляции в пигментированных сладких апельсинах [( Citrus sinensis ) L. Osbeck]. J Agric Food Chem. (2015) 63:4031–41. doi: 10.1021/acs.jafc.5b01123
PubMed Abstract | Полный текст перекрестной ссылки | Google Scholar
35. Сайяри М., Бабалар М., Калантари С., Мартинес-Ромеро Д. , Гильен Ф., Серрано М. и др. Обработка парами метилсалицилата или метилжасмоната облегчала повреждение от переохлаждения и повышала антиоксидантный потенциал во время послеуборочного хранения граната. Пищевая хим. (2011) 124:964–70. doi: 10.1016/j.foodchem.2010.07.036
, Гильен Ф., Серрано М. и др. Обработка парами метилсалицилата или метилжасмоната облегчала повреждение от переохлаждения и повышала антиоксидантный потенциал во время послеуборочного хранения граната. Пищевая хим. (2011) 124:964–70. doi: 10.1016/j.foodchem.2010.07.036
CrossRef Полный текст | Академия Google
Влияние местного применения глицина и пролина на резистентные язвы ног при дефиците пролидазы | JAMA Dermatology
Влияние местного применения глицина и пролина на резистентные язвы ног при дефиците пролидазы | JAMA Дерматология | Сеть ДЖАМА [Перейти к навигации]Эта проблема
- Скачать PDF
- Полный текст
Поделиться
Твиттер Фейсбук Эл.
адрес
LinkedIn- Процитировать это
- Разрешения
 адрес
LinkedIn
адрес
LinkedInАртикул
Июнь 1986 г.
Дзиро Арата, MD ; Кеничи Хатакенака, MD ; Такаши Оно, MD
Принадлежности автора
Кафедра дерматологии Медицинская школа Коти Кочи, 781-51 Нанкоку-ши, Япония
Арка Дерматол. 1986;122(6):626-627. doi: 10.1001 / archderm.1986.01660180026006
Полный текст
Абстрактный
В редакцию.— Наиболее важной дерматологической проблемой дефицита пролидазы 1 являются хронические рецидивирующие язвы на ногах, которые плохо заживают. Мы использовали 5% глицин-5% пролиновую мазь при язвах на ногах с отличными результатами.
Отчет о деле.— Пациентка, сейчас 22-летняя женщина, та же пациентка, что и описанная ранее. 1 У нее никогда не было рецидивирующих язв на ногах, несмотря на различные традиционные методы лечения, включая местное применение антибиотиков (сульфат гентамицина, сульфат колистина, сульфат полимиксина В и т. д.), ванны для ног с 0,01% раствором перманганата калия, пероральное введение сульфата цинка ( 180 мг в сутки) и/или аскорбиновой кислоты (3 г в сутки). Пероральный L-пролин (1 г в день) также пробовали в течение двух месяцев без видимой пользы. Мы решили использовать местный пролин, а затем комбинацию местного пролина и глицина на ее язвах на ногах. L-пролин и/или L-глицин, растертые в ступке пестиком, смешивали с жидким парафином (30 г) и
Полный текст
Добавить или изменить учреждение
- Кислотно-щелочное, электролиты, жидкости
- Лечение зависимости
- Аллергия и клиническая иммунология
- Анестезиология
- Антикоагулянты
- Искусство и изображения в психиатрии
- Кровотечение и переливание
- Кардиология
- Уход за тяжелобольным пациентом
- Проблемы клинической электрокардиографии
- Клинический вызов
- Поддержка принятия клинических решений
- Клинические последствия базовой нейронауки
- Клиническая фармация и фармакология
- Дополнительная и альтернативная медицина
- Заявления о консенсусе
- Коронавирус (COVID-19)
- Медицина интенсивной терапии
- Культурная компетенция
- Стоматология
- Дерматология
- Диабет и эндокринология
- Интерпретация диагностического теста
- Разнообразие, равенство и инклюзивность
- Разработка лекарств
- Электронные медицинские карты
- Скорая помощь
- Конец жизни
- Гигиена окружающей среды
- Этика
- Пластическая хирургия лица
- Гастроэнтерология и гепатология
- Генетика и геномика
- Геномика и точное здоровье
- Гериатрия
- Глобальное здравоохранение
- Справочник по статистике и медицине
- Рекомендации
- Заболевания волос
- Модели оказания медицинской помощи
- Экономика здравоохранения, страхование, оплата
- Качество медицинской помощи
- Реформа здравоохранения
- Медицинская безопасность
- Медицинские работники
- Различия в состоянии здоровья
- Несправедливость в отношении здоровья
- Информатика здравоохранения
- Политика здравоохранения
- Гематология
- История медицины
- Гуманитарные науки
- Гипертония
- Изображения в неврологии
- Наука внедрения
- Инфекционные болезни
- Инновации в оказании медицинской помощи
- Инфографика JAMA
- Право и медицина
- Ведущее изменение
- Чем меньше, тем лучше
- ЛГБТК
- Образ жизни
- Медицинский код
- Медицинские приборы и оборудование
- Медицинское образование
- Медицинское образование и обучение
- Медицинские журналы и публикации
- Меланома
- Мобильное здравоохранение и телемедицина
- Нарративная медицина
- Нефрология
- Неврология
- Неврология и психиатрия
- Примечательные примечания
- Сестринское дело
- Питание
- Питание, Ожирение, Упражнения
- Ожирение
- Акушерство и гинекология
- Гигиена труда
- Онкология
- Офтальмологические изображения
- Офтальмология
- Ортопедия
- Отоларингология
- Лекарство от боли
- Патология и лабораторная медицина
- Уход за пациентами
- Информация для пациентов
- Педиатрия
- Повышение производительности
- Показатели эффективности
- Периоперационный уход и консультация
- Фармакоэкономика
- Фармакоэпидемиология
- Фармакогенетика
- Фармация и клиническая фармакология
- Физическая медицина и реабилитация
- Физиотерапия
- Руководство врачей
- Поэзия
- Здоровье населения
- Профилактическая медицина
- Профессиональное благополучие
- Профессионализм
- Психиатрия и поведенческое здоровье
- Общественное здравоохранение
- Легочная медицина
- Радиология
- Регулирующие органы
- Исследования, методы, статистика
- Реанимация
- Ревматология
- Управление рисками
- Научные открытия и будущее медицины
- Совместное принятие решений и общение
- Медицина сна
- Спортивная медицина
- Трансплантация стволовых клеток
- Хирургия
- Хирургические инновации
- Хирургический жемчуг
- Обучаемый момент
- Технологии и финансы
- Искусство JAMA
- Искусство и медицина
- Рациональное клиническое обследование
- Табак и электронные сигареты
- Токсикология
- Травмы и травмы
- Приверженность лечению
- УЗИ
- Урология
- Руководство пользователя по медицинской литературе
- Вакцинация
- Венозная тромбоэмболия
- Здоровье ветеранов
- Насилие
- Женское здоровье
- Рабочий процесс и процесс
- Уход за ранами, инфекция, заживление
Сохранить настройки
Политика конфиденциальности | Условия использования
Трансформация аминокислот глицина при ударах малых тел Солнечной системы, смоделированная методом кручения под высоким давлением химическая модификация как земных, так и малых тел Солнечной системы.
 В этом контексте открытие аминокислоты глицина в метеоритах и кометах привело к гипотезе о том, что удары астрономических тел могли способствовать доставке и полимеризации аминокислот на ранней Земле с образованием белков как необходимых молекул для жизни. Помимо возможности абиотической полимеризации глицина, его разложение под действием ударов может генерировать реакционноспособные группы с образованием других важных органических биомолекул. В данном исследовании к глицину был применен метод кручения под высоким давлением (HPT) как новая платформа для моделирования ударов малых тел Солнечной системы. По сравнению с экспериментами с ударом высокого давления метод КВД одновременно вводит высокое давление и деформационную деформацию. Было обнаружено, что глицин не полимеризовался в исследуемых экспериментальных условиях, а частично разложился до этанола при давлении 1 и 6 ГПа и деформации сдвига < 120 м/м. Обнаружение этанола предполагает присущую ему доступность оставшихся азотсодержащих групп, которые могут включаться в образование других органических молекул в месте удара.
В этом контексте открытие аминокислоты глицина в метеоритах и кометах привело к гипотезе о том, что удары астрономических тел могли способствовать доставке и полимеризации аминокислот на ранней Земле с образованием белков как необходимых молекул для жизни. Помимо возможности абиотической полимеризации глицина, его разложение под действием ударов может генерировать реакционноспособные группы с образованием других важных органических биомолекул. В данном исследовании к глицину был применен метод кручения под высоким давлением (HPT) как новая платформа для моделирования ударов малых тел Солнечной системы. По сравнению с экспериментами с ударом высокого давления метод КВД одновременно вводит высокое давление и деформационную деформацию. Было обнаружено, что глицин не полимеризовался в исследуемых экспериментальных условиях, а частично разложился до этанола при давлении 1 и 6 ГПа и деформации сдвига < 120 м/м. Обнаружение этанола предполагает присущую ему доступность оставшихся азотсодержащих групп, которые могут включаться в образование других органических молекул в месте удара. Кроме того, это открытие подчеркивает возможность происхождения этанола, ранее обнаруженного в кометах.
Кроме того, это открытие подчеркивает возможность происхождения этанола, ранее обнаруженного в кометах.Введение
РНК (рибонуклеиновая кислота) и белок являются двумя важными полимерными молекулами для жизни, но неясно, как эти биомолекулы образовались около четырех миллиардов лет назад на Земле из нуклеозидов (таких как аденозинмонофосфат) и мономеров аминокислот. , соответственно 1 . Открытие молекул аминокислот, таких как глицин (C 2 H 5 NO 2 ) в метеорите Мерчисон (упал в Австралии в 1969) предположил, что столкновения с небольшими телами Солнечной системы (такими как метеороиды, астероиды, кометы и переходные объекты) могли принести важные органические молекулы для жизни на Землю 2 . Недавнее исследование различных типов углеродистых хондритовых метеоритов, в том числе метеорита озера Тагиш (упал в Канаде в 2000 г.), также подтвердило наличие аминокислот, которые могут выжить при входе в атмосферу, ударном воздействии и водном чередовании 3 . Более того, теоретические расчеты показали, что высокое давление, высокая температура и высокая скорость деформации во время астрономических ударов могут образовывать белковые цепи из аминокислот 4 . До сих пор не было убедительных экспериментальных доказательств образования белка из аминокислот путем моделирования удара с помощью экспериментов с ударом высокого давления 5,6,7,8 .
Более того, теоретические расчеты показали, что высокое давление, высокая температура и высокая скорость деформации во время астрономических ударов могут образовывать белковые цепи из аминокислот 4 . До сих пор не было убедительных экспериментальных доказательств образования белка из аминокислот путем моделирования удара с помощью экспериментов с ударом высокого давления 5,6,7,8 .
При ударах малых тел Солнечной системы давление, температура и скорость деформации в точке удара высоки, но они уменьшаются по мере удаления от точки столкновения 9 . Причем деформационная деформация возникает на разном расстоянии от точки удара, как схематично показано на рис. 1а 9.0045 10 . В то время как ударные эксперименты подходят для моделирования условий высокого давления в точке удара, они неэффективны для моделирования условий на некоторых расстояниях от точки столкновения, где деформационная деформация высока, а давление и температура довольно низки 11 . До сих пор предпринималось мало попыток рассмотреть одновременное влияние высокого давления и высокой деформации на полимеризацию или стабильность аминокислот.
До сих пор предпринималось мало попыток рассмотреть одновременное влияние высокого давления и высокой деформации на полимеризацию или стабильность аминокислот.
( a ) Иллюстрация столкновений метеороидов, астероидов, комет и переходных объектов с Землей 10 ; ( b ) схема метода кручения под высоким давлением и его наковальни 13 ; и ( c ) поверхность торсионной наковальни высокого давления после небольшого взрыва, вызванного обработкой глицином под давлением 6 ГПа.
Изображение в натуральную величину
Метод кручения под высоким давлением (HPT) может быть подходящим методом для моделирования событий астрономического удара, поскольку в дополнение к давлению, температуре и скорости деформации в этом методе можно одновременно контролировать деформацию 12 . Образец в форме диска сжимается между двумя наковальнями под высоким давлением, и в этом методе деформация вызывается вращением одной из наковальней относительно другой, как показано на рис. 1b 13 . В прошлом веке ученые из США 14,15 и СССР 16,17 применили метод КВД к некоторым органическим соединениям и сообщили об усилении полимеризации или деполимеризации; тем не менее, эти интересные исследования были практически проигнорированы позже, как обсуждалось в обзорной статье 9.0045 13 . Метод HPT в настоящее время используется в качестве метода интенсивной пластической деформации 18,19 для контроля размера кристаллов 12,20 и фазового превращения 21,22 в неорганических материалах. Этот метод считается достаточно эффективным для соединения атомов различных чистых элементов с образованием более крупных молекул металлических сплавов 23 и многокомпонентной керамики 24 .
1b 13 . В прошлом веке ученые из США 14,15 и СССР 16,17 применили метод КВД к некоторым органическим соединениям и сообщили об усилении полимеризации или деполимеризации; тем не менее, эти интересные исследования были практически проигнорированы позже, как обсуждалось в обзорной статье 9.0045 13 . Метод HPT в настоящее время используется в качестве метода интенсивной пластической деформации 18,19 для контроля размера кристаллов 12,20 и фазового превращения 21,22 в неорганических материалах. Этот метод считается достаточно эффективным для соединения атомов различных чистых элементов с образованием более крупных молекул металлических сплавов 23 и многокомпонентной керамики 24 .
В этом исследовании мы впервые применяем метод HPT к аминокислоте глицину, чтобы получить представление о выживании аминокислот или образовании белка в результате астрономических ударов. Результаты этого исследования имеют значение для астробиологии, поскольку степень выживания органических материалов при ударах малых тел Солнечной системы является важным вопросом в понимании доставки необходимых для жизни молекул на Землю кометами и другими малыми телами Солнечной системы 25,26 . Более того, механизм образования белков из аминокислот в условиях ранней Земли до сих пор остается ключевым вопросом в понимании происхождения жизни 1,8 .
Более того, механизм образования белков из аминокислот в условиях ранней Земли до сих пор остается ключевым вопросом в понимании происхождения жизни 1,8 .
Методы
Порошок глицина с химическим составом C 2 H 5 NO 2 , степенью чистоты > 99%, плотностью 1,2 кг/м 3 кристаллической структуры и моноклинной (α-фаза с пространственной группой P21/n) была выбрана для данного исследования. Порошок прессовали под давлением 0,3 МПа в диски диаметром 10 мм и толщиной 0,8 мм (почти 80 мг массы каждого диска). Образцы дисков обрабатывались между двумя цилиндрическими наковальнями ТВД диаметром 50 мм и высотой 50 мм, изготовленными из композита WC — 11 мас.% Со. Поверхности наковален имели наклон 5° от центра к краю с плоскодонным отверстием диаметром 10 мм и высотой 0,25 мм в центре. Спрессованные диски обрабатывались под давлением P = 1 и 6 ГПа при температуре окружающей среды ( T = 300 К). Хотя более ранние исследования предполагали, что давление может находиться в диапазоне 5–25 ГПа 25 или обычно ниже 30 ГПа 5 во время столкновения с внеземными объектами, текущее исследование считает, что более низкие давления, такие как 1 ГПа, могут быть естественным образом созданы с помощью увеличение расстояния от точки столкновения за счет распределения нормального давления по отношению к расстоянию 19 . Деформация сдвига, γ ( γ = 2π rN / h , r : расстояние от центра диска, N : обороты наковальни, h : вращение наковальни, h : толщина образца ниже 900) 1900 наковальня относительно верхней со скоростью вращения 1 об/мин (оборот в минуту) для Н = 3 оборотов. Деформация сдвига теоретически была нулевой в центре диска и увеличивалась до 120 м/м на краю диска. Образец, обработанный под давлением 6 ГПа, испытал небольшой взрыв в начале вращения, который вызвал значительный удар по образцу. На рисунке 1c показан внешний вид поверхности наковальни после обработки глицином под давлением 6 ГПа. Исходный порошок глицина и образцы, обработанные ГВД, исследовали различными методами, как описано ниже.
Деформация сдвига, γ ( γ = 2π rN / h , r : расстояние от центра диска, N : обороты наковальни, h : вращение наковальни, h : толщина образца ниже 900) 1900 наковальня относительно верхней со скоростью вращения 1 об/мин (оборот в минуту) для Н = 3 оборотов. Деформация сдвига теоретически была нулевой в центре диска и увеличивалась до 120 м/м на краю диска. Образец, обработанный под давлением 6 ГПа, испытал небольшой взрыв в начале вращения, который вызвал значительный удар по образцу. На рисунке 1c показан внешний вид поверхности наковальни после обработки глицином под давлением 6 ГПа. Исходный порошок глицина и образцы, обработанные ГВД, исследовали различными методами, как описано ниже.
Кристаллическую структуру исследовали методом рентгеновской дифракции (РД) с использованием α-излучения Cu K с длиной волны 0,15406 нм (45 кВ и 200 мА) и методом рамановской спектроскопии с использованием лазерного источника с длиной волны 532 нм. Профили XRD были получены с использованием полного диска, и, таким образом, они представляют кристаллическую структуру от центра к краю диска, то есть от деформации сдвига от нуля до 120 м/м. Спектры комбинационного рассеяния были записаны в области сильного напряжения на расстоянии 4 мм от центра диска, хотя не было обнаружено четкой разницы между спектрами комбинационного рассеяния в центре и на краю диска.
Профили XRD были получены с использованием полного диска, и, таким образом, они представляют кристаллическую структуру от центра к краю диска, то есть от деформации сдвига от нуля до 120 м/м. Спектры комбинационного рассеяния были записаны в области сильного напряжения на расстоянии 4 мм от центра диска, хотя не было обнаружено четкой разницы между спектрами комбинационного рассеяния в центре и на краю диска.
Полимеризацию или разложение образцов исследовали с помощью спектроскопии ядерного магнитного резонанса (ЯМР). Около 20 мг образцов глицина растворяли в 0,6 мл D 2 O и помещали в пробирку для ЯМР (внутренний диаметр: 5 мм). C 6 D 6 в стеклянном капилляре также помещали в ЯМР-пробирку в качестве внутреннего стандарта. Спектры ЯМР 1 H и 13 C ЯМР регистрировали при 600 и 150 МГц соответственно.
Результаты
Исследование кристаллических структур методом РФА показано на рис. 2а. Сравнение профиля XRD исходного порошка и справочных данных (№ карты PDF: 00-032-1702) подтверждает, что исходный порошок содержит только α-глицин. Кристаллическая структура остается неизменной после обработки HPT, и не происходит превращений β-, γ- или δ-фаз или других полиморфов глицина 9 под высоким давлением.0045 27,28 . Исследование кристаллической структуры методом КР-спектроскопии, как показано на рис. 2б, также подтверждает, что исходный порошок и образцы, обработанные КВД под давлением 1 и 6 ГПа, имеют близкие спектры. Сравнение этих спектров с указанными в литературе волновыми числами 29,30 подтверждает, что три образца содержат чистый α-глицин. В целом, рис. 2 предполагает, что необратимые фазовые превращения не происходят в глицине при обработке HPT, и это согласуется с сообщениями о стабильности α-глицина при комнатной температуре и при давлениях до 50 ГПа 31 .
Кристаллическая структура остается неизменной после обработки HPT, и не происходит превращений β-, γ- или δ-фаз или других полиморфов глицина 9 под высоким давлением.0045 27,28 . Исследование кристаллической структуры методом КР-спектроскопии, как показано на рис. 2б, также подтверждает, что исходный порошок и образцы, обработанные КВД под давлением 1 и 6 ГПа, имеют близкие спектры. Сравнение этих спектров с указанными в литературе волновыми числами 29,30 подтверждает, что три образца содержат чистый α-глицин. В целом, рис. 2 предполагает, что необратимые фазовые превращения не происходят в глицине при обработке HPT, и это согласуется с сообщениями о стабильности α-глицина при комнатной температуре и при давлениях до 50 ГПа 31 .
Отсутствие фазового превращения в глицине после обработки кручением под высоким давлением. ( a ) Профили XRD и ( b ) Спектры комбинационного рассеяния глицина до и после обработки при 1 и 6 ГПа.
Изображение полного размера
Полимеризацию глицина исследовали с помощью (а) 1 H и (b) 13 C ЯМР-спектроскопии, как показано на рис. 3. Для исходного порошка только пики, соответствующие чистым глицина хорошо согласуются со справочными данными 32 . После обработки КВД под давлением 1 и 6 ГПа обнаруживаются пики, происходящие от пиков глицина, что хорошо согласуется с данными РФА и КР-спектроскопии. Отсутствие уширения или смещения пиков глицина свидетельствует об отсутствии полимеризации. Помимо пиков глицина и эталона C 6 D 6 , пики при 3,65 и 1,17 м.д. в 1 H ЯМР и 17,5 и 58,0 м.д. в 13 C ЯМР могут быть отнесены к этанолу (этиловый спирт, C Х 6 О) после обработки КВД 25 . Образование этанола свидетельствует о том, что глицин разлагается, а не полимеризуется в условиях обработки КВД. Азотосодержащие продукты или продукты разложения на основе аминогрупп не могут быть отслежены методом ЯМР, что позволяет предположить, что они испаряются/реагируют быстро и перемещаются в атмосферу воздуха, в то время как выделяющийся этанол остается в образце в течение достаточного времени во время измерений ЯМР-спектроскопии.
Отсутствие полимеризации и наличие превращения в этанол в глицине при обработке скручиванием под высоким давлением. ЯМР ( a ) 1 H и ( b ) 13 C спектры глицина до и после обработки под давлением 1 и 6 ГПа.
Увеличенное изображение
Обсуждение
Текущие результаты первого применения метода ГВД к глициновой аминокислоте не дают никаких доказательств полимеризации и образования белка, но большие количества глицина могут выживать при одновременном воздействии высоких давление и деформация сдвига. Поскольку глицин является одной из стабильных аминокислот с низкой теплотой образования, отсутствие полимеризации неудивительно. Более ранние шоковые эксперименты Петерсона и др. . показали, что полимеризация аминокислот не происходит в диапазоне давлений от 3,5 до 32 ГПа 5 . Бланк и др. . также показал, что значительное количество аминокислот может выживать при высоких давлениях (5,1–21 ГПа) и температурах (412–870 К) 6 . Однако образование этанола из глицина за счет эффекта дополнительной деформации сдвига, впервые наблюдаемое в настоящей работе, довольно необычно, поскольку этанол имеет более высокую теплоту образования по сравнению с глицином. Образование этанола должно быть связано со значительной энергией, индуцированной в процессе КВД 12,13 , которые могут изменять термодинамическую устойчивость при высоком давлении и деформации 21,22 . Более ранние исследования по обработке HPT подтвердили, что такие термодинамические изменения могут двигаться в разных направлениях 19 , например, распад больших молекул 22 или объединение малых атомов с образованием более крупных молекул 22,23,24 , даже когда малые атомы термодинамически несмешивающийся 33 . Для органических материалов классические исследования также показали, что разложение или полимеризация могут происходить под высоким давлением и высокой деформацией, в зависимости от состава материала, предполагая, что на термодинамическую стабильность можно влиять в разных направлениях и в органических материалах 14,15,16,17 .
Однако образование этанола из глицина за счет эффекта дополнительной деформации сдвига, впервые наблюдаемое в настоящей работе, довольно необычно, поскольку этанол имеет более высокую теплоту образования по сравнению с глицином. Образование этанола должно быть связано со значительной энергией, индуцированной в процессе КВД 12,13 , которые могут изменять термодинамическую устойчивость при высоком давлении и деформации 21,22 . Более ранние исследования по обработке HPT подтвердили, что такие термодинамические изменения могут двигаться в разных направлениях 19 , например, распад больших молекул 22 или объединение малых атомов с образованием более крупных молекул 22,23,24 , даже когда малые атомы термодинамически несмешивающийся 33 . Для органических материалов классические исследования также показали, что разложение или полимеризация могут происходить под высоким давлением и высокой деформацией, в зависимости от состава материала, предполагая, что на термодинамическую стабильность можно влиять в разных направлениях и в органических материалах 14,15,16,17 .
Здесь следует отметить, что считается, что спирты, включая этанол, могли способствовать эволюции органических соединений на ранней Земле 34 , а также есть недавние сообщения об открытии этанола в кометах 35 ; однако направление таких вкладов еще предстоит изучить. Поскольку наличие глицина было подтверждено как минимум у двух комет 81P/Wild-2 7 и 67P/Чурюмова-Герасименко 36 , текущие эксперименты предполагают , что глицин может быть естественным источником огромного количества этанола , обнаруженного в некоторых кометах 35 . Следует отметить, что кометы могут испытывать деформации сдвига под действием приливных сил планет и звезд. В то время как некоторые большие кометы, такие как C/2014 Q2 (Лавджой), в которой был обнаружен спирт, могут выдержать эти приливные силы 35 , некоторые другие кометы испытывают разрушение под действием этих сил, например, кометы Крейца, разрушенные солнцем 37 или кометой D. /1993 F2 Шумейкер-Леви 9 разрушен Юпитером 38 . Более того, гравитационные силы планет и звезд могут привести к столкновению этих расколотых комет с другими астрономическими телами, создающими достаточное давление для превращения аминокислот в спирт, особенно в присутствии ледяной воды, водных альтераторов и минеральных катализаторов 39 .
/1993 F2 Шумейкер-Леви 9 разрушен Юпитером 38 . Более того, гравитационные силы планет и звезд могут привести к столкновению этих расколотых комет с другими астрономическими телами, создающими достаточное давление для превращения аминокислот в спирт, особенно в присутствии ледяной воды, водных альтераторов и минеральных катализаторов 39 .
Несмотря на образование этанола из глицина в процессе ГВД, все еще трудно прояснить путь этого превращения, поскольку эксперименты ГВД проводятся в атмосфере окружающей среды, и поэтому газовые фазы не могут быть проанализированы. Превращение глицина широко изучалось в различных условиях, таких как высокое давление 8 , термический нагрев 40 , ионизирующее излучение 41 , протонированное состояние 42 , вода высокого давления/температуры 43 и электрохимическое состояние 44 . Ни в одном из этих исследований не сообщалось об образовании этанола из глицина, хотя электрохимическое образование метанольных и пропаноловых спиртов упоминалось в публикации 45 . Хотя отследить путь превращения глицина экспериментально сложно из-за короткого времени жизни радикалов 46 , а также за счет образования различных газообразных соединений (таких как H 2 , H 2 O, CO, CO 2 , N 2 и NH 3 )
Хотя отследить путь превращения глицина экспериментально сложно из-за короткого времени жизни радикалов 46 , а также за счет образования различных газообразных соединений (таких как H 2 , H 2 O, CO, CO 2 , N 2 и NH 3 )
 Отсутствие запаха аммиака в процессе КВД свидетельствует о том, что NH 3 в заметных количествах, возможно, не образовался, хотя во многих публикациях сообщалось об образовании NH 3 при разложении глицина 41,42,43,44,45,49 . Отсутствие NH 3 свидетельствует о том, что высокая энергия, вносимая КВД, не только способствует реакции отщепления с образованием аминогруппы путем разрыва связей C-N, но и разрывает связи N-H, что впоследствии может привести к образованию азота и водорода. газовые фазы. Другой возможностью является реакция аминогруппы с кислородом и образованием газообразного водорода и NO, который впоследствии может превратиться в NO 9.0039 2 в присутствии кислорода. Более того, как предполагается в некоторых вычислительных исследованиях 47,48 , ожидается, что декарбоксилирование, связанное с переносом протона, приведет к образованию СО или СО 2 . Здесь предлагаются три возможных примера (1) разложения, (2) реакции с водой и (3) окисления глицина.
Отсутствие запаха аммиака в процессе КВД свидетельствует о том, что NH 3 в заметных количествах, возможно, не образовался, хотя во многих публикациях сообщалось об образовании NH 3 при разложении глицина 41,42,43,44,45,49 . Отсутствие NH 3 свидетельствует о том, что высокая энергия, вносимая КВД, не только способствует реакции отщепления с образованием аминогруппы путем разрыва связей C-N, но и разрывает связи N-H, что впоследствии может привести к образованию азота и водорода. газовые фазы. Другой возможностью является реакция аминогруппы с кислородом и образованием газообразного водорода и NO, который впоследствии может превратиться в NO 9.0039 2 в присутствии кислорода. Более того, как предполагается в некоторых вычислительных исследованиях 47,48 , ожидается, что декарбоксилирование, связанное с переносом протона, приведет к образованию СО или СО 2 . Здесь предлагаются три возможных примера (1) разложения, (2) реакции с водой и (3) окисления глицина.
$${\text{2C}}_{{2}} {\text{H}}_{{5}} {\text{NO}}_{{2}} \to {\text{C }}_{{2}} {\text{H}}_{{6}} {\text{O}} + {\text{H}}_{{2}} + {\text{H}} _{{2}} {\text{O}} + {\text{2CO}} + {\text{N}}_{{2}}$$
(1)
$${\text{2C}}_{{2}} {\text{H}}_{{5}} {\text{NO}}_{{2}} + { \text{H}}_{{2}} {\text{O}} \to {\text{C}}_{{2}} {\text{H}}_{{6}} {\text {O}} + {\text{3H}}_{{2}} + {\text{2CO}} + {\text{N}}_{{2}}$$
(2)
$ ${\text{2C}}_{{2}} {\text{H}}_{{5}} {\text{NO}}_{{2}} + { 7}/{\text{2O }}_{{2}} \to {\text{C}}_{{2}} {\text{H}}_{{6}} {\text{O}} + {\text{2H} }_{{2}} {\text{O}} + {\text{CO}}_{{2}} + {\text{NO}}_{{2}}$$
(3)
Эти предположительные пути трансформации показывают возможность трансформации глицина в этанол под действием внешней энергии, но необходимы дальнейшие исследования, чтобы уточнить точный путь химической реакции и лежащий в ее основе механизм.
Хотя текущие эксперименты HPT не предоставили каких-либо положительных подтверждений образования белка из глицина в результате астрономических событий, образование белка из других аминокислот с помощью процессинга HPT необходимо исследовать в будущем. Стабильность значительных количеств глицина после процесса HPT указывает на то, что столкновения с кометами могли в основном доставлять значительные количества биомолекул на раннюю Землю, как предполагалось в некоторых других исследованиях с помощью экспериментов с ударами высокого давления 9.0045 25,26 . Как обсуждалось в предыдущем абзаце, превращение глицина в этанол подразумевает выделение в систему азотсодержащих молекул и некоторых реакционноспособных газов, которые неизбежно доступны для взаимодействия с другими молекулами с образованием других органических соединений на Земле. Одно из основных отличий этого исследования от реальных столкновений малых тел Солнечной системы состоит в том, что текущие эксперименты HPT проводились с глицином высокой чистоты; в то время как аминокислоты в условиях ранней Земли или в кометах присутствуют вместе с некоторыми минералами, химическими веществами и водой 1,2,8,34 . Наличие воды и водного чередования в углеродистых хондритовых метеоритах может влиять на образование органических соединений 39,50 .
Стабильность значительных количеств глицина после процесса HPT указывает на то, что столкновения с кометами могли в основном доставлять значительные количества биомолекул на раннюю Землю, как предполагалось в некоторых других исследованиях с помощью экспериментов с ударами высокого давления 9.0045 25,26 . Как обсуждалось в предыдущем абзаце, превращение глицина в этанол подразумевает выделение в систему азотсодержащих молекул и некоторых реакционноспособных газов, которые неизбежно доступны для взаимодействия с другими молекулами с образованием других органических соединений на Земле. Одно из основных отличий этого исследования от реальных столкновений малых тел Солнечной системы состоит в том, что текущие эксперименты HPT проводились с глицином высокой чистоты; в то время как аминокислоты в условиях ранней Земли или в кометах присутствуют вместе с некоторыми минералами, химическими веществами и водой 1,2,8,34 . Наличие воды и водного чередования в углеродистых хондритовых метеоритах может влиять на образование органических соединений 39,50 . Кроме того, реактивные минералы в метеоритах могут действовать как катализаторы для образования сложных органических соединений, включая аминокислоты и спирты, особенно в присутствии горячей воды 51,52 . Таким образом, хотя текущие результаты представляют собой новый экспериментальный путь для моделирования воздействия ударов комет или других малых тел Солнечной системы на органические молекулы, будущие эксперименты HPT с различными аминокислотами и аденозинмонофосфатом с различными уровнями чистоты и различными добавками как в сухом, так и в требуются влажные условия. Более того, теоретические расчеты, подобные тем, о которых сообщалось ранее 4 может быть полезен для нахождения энергетически благоприятного направления реакции в процессинге HPT с образованием белка и РНК.
Кроме того, реактивные минералы в метеоритах могут действовать как катализаторы для образования сложных органических соединений, включая аминокислоты и спирты, особенно в присутствии горячей воды 51,52 . Таким образом, хотя текущие результаты представляют собой новый экспериментальный путь для моделирования воздействия ударов комет или других малых тел Солнечной системы на органические молекулы, будущие эксперименты HPT с различными аминокислотами и аденозинмонофосфатом с различными уровнями чистоты и различными добавками как в сухом, так и в требуются влажные условия. Более того, теоретические расчеты, подобные тем, о которых сообщалось ранее 4 может быть полезен для нахождения энергетически благоприятного направления реакции в процессинге HPT с образованием белка и РНК.
Выводы
Получить представление о влиянии ударов малых тел Солнечной системы (например, метеороидов, астероидов, комет и переходных объектов) на стабильность аминокислот или образование белков, одновременное воздействие высокого давления деформацию сдвига при полимеризации аминокислоты глицина исследовали методом кручения под высоким давлением. Глицин не полимеризовался, но частично разложился и при обработке превратился в этанол. Обнаружение этанола не только указывает на природный источник этанола, уже обнаруженный в кометах, но также предполагает образование других продуктов разложения, которые могли способствовать образованию других органических биомолекул на Земле миллиарды лет назад. Хотя это исследование не предоставило никаких доказательств образования белков, оно представило метод кручения под высоким давлением в качестве нового исследовательского инструмента для моделирования влияния ударов малых тел Солнечной системы на поведение органических материалов в условиях ранней Земли.
Глицин не полимеризовался, но частично разложился и при обработке превратился в этанол. Обнаружение этанола не только указывает на природный источник этанола, уже обнаруженный в кометах, но также предполагает образование других продуктов разложения, которые могли способствовать образованию других органических биомолекул на Земле миллиарды лет назад. Хотя это исследование не предоставило никаких доказательств образования белков, оно представило метод кручения под высоким давлением в качестве нового исследовательского инструмента для моделирования влияния ударов малых тел Солнечной системы на поведение органических материалов в условиях ранней Земли.
Ссылки
Патель, Б., Персиваль, К., Ритсон, Д. Дж., Даффи, С. Д. и Сазерленд, Дж. Д. Общее происхождение предшественников РНК, белков и липидов в цианосульфидном протометаболизме. Нац. хим. 7 , 310–307 (2015).
Google ученый
Александр, К.О., Коди, Г.Д., Де Грегорио, Б.Т., Ниттлер, Л.Р. и Страуд, Р.М. Природа, происхождение и модификация нерастворимого органического вещества в хондритах, основном источнике углерода и азота на Земле. Chem. Эрде Гечем. 77 , 227–256 (2017).
ОБЪЯВЛЕНИЕ КАС Google ученый
Kroonblawd, M.P., Lindsey, R.K. & Goldman, N. Синтез функционализированных азотсодержащих полициклических ароматических углеводородов и других пребиотических соединений в растворах глицина. Хим. науч. 10 , 6091–6098 (2019).
КАС пабмед ПабМед Центральный Google ученый
Бланк, Дж. Г., Миллер, Г. Х. и Винанс, Р. Э. Экспериментальная ударная химия водных растворов аминокислот и кометная доставка пребиотических соединений. Происхождение Life Evol. биосф. 31 , 15–51 (2001).
ОБЪЯВЛЕНИЕ КАС Google ученый
Мартинс, З., Прайс, М. К., Голдман, Н., Сефтон, М. А. и Берчелл, М. Дж. Ударный синтез аминокислот при столкновении с кометами и аналогами поверхности ледяных планет. Нац. Geosci. 6 , 1045–1049 (2013).
ОБЪЯВЛЕНИЕ КАС Google ученый
Китадай Н. и Маруяма С.
Происхождение строительных блоков жизни: обзор. Геофизика. Фронт. 9 , 1117–1153 (2018).КАС Google ученый
Фриц Дж., Грешак А. и Фернандес В.А. Пересмотр классификации ударов метеоритов. Меторит. Планета. науч. 52 , 1216–1232 (2017).
ОБЪЯВЛЕНИЕ КАС Google ученый
Дэвис, Д. Библиотека изображений и видео НАСА , ARC-1991-AC91-0193 (1991).
Даффи Т.С. и Смит Р.Ф. Динамическое сжатие геологических материалов при сверхвысоком давлении. Фронт. наук о Земле. 7 , 23 (2019).
ОБЪЯВЛЕНИЕ Google ученый
Жиляев А. П., Лэнгдон Т. Г. Использование кручения под высоким давлением для обработки металлов: основы и приложения. Прог. Матер.
науч. 53 , 893–979 (2008).КАС Google ученый
Эдалати, К. и Хорита, З. Обзор кручения под высоким давлением (HPT) от 1935 по 1988 год. Матер. науч. англ. А 652 , 325–352 (2016).
КАС Google ученый
Бриджмен, П. В. Влияние высокого напряжения сдвига в сочетании с высоким гидростатическим давлением. Физ. 48 , 825–847 (1935).
ОБЪЯВЛЕНИЕ КАС Google ученый
Bridgman, P.W. Полиморфные переходы до 50 000 кг/см 2 нескольких органических веществ. Проц. Являюсь. акад. Наук искусств. 72 , 227–268 (1938).
КАС Google ученый
Пахомова И.Е., Жулин В.М., Гоникберг М.Г., Иванов В.
П. Превращения нитрилов в твердой фазе при высоких давлениях в сочетании со сдвигом. Бык. акад. науч. дивизия СССР хим. науч. 21 , 813–818 (1972).Google ученый
Жорин В. А., Кисин Ю. В., Луизо Ю. В., Фридман Н. М., Ениколопян Н. С. Структурные изменения в полиолефинах, обусловленные сочетанием высокого давления и сдвиговой деформации. Полим. науч. СССР 18 , 3057–3061 (1976).
Google ученый
Валиев Р. З. и др. Производство сыпучих ультрамелкозернистых материалов путем интенсивной пластической деформации. JOM 58 (4), 33–39 (2006).
ОБЪЯВЛЕНИЕ Google ученый
Эдалати, К. и др. Наноматериалы при сильной пластической деформации: обзор исторических событий и последних достижений.
Матер. Рез. лат. 10 , 163–256 (2022).КАС Google ученый
Пиппан, Р. и др. Насыщение фрагментации при интенсивной пластической деформации. год. Преподобный Матер. Рез. 40 , 319–343 (2010).
ОБЪЯВЛЕНИЕ КАС Google ученый
Левитас В. И. Фазовые превращения при высоком давлении при интенсивной пластической деформации кручением во вращающихся наковальнях. Матер. Транс. 60 , 1294–1301 (2019).
КАС Google ученый
Мазилкин А., Страумал Б., Килмаметов А., Страумал П. и Барецки Б. Фазовые превращения, вызванные интенсивной пластической деформацией. Матер. Транс. 60 , 1489–1499 (2019).
КАС Google ученый
Акрами С., Эдалати П., Фуджи М. и Эдалати К. Высокоэнтропийная керамика: обзор принципов, производства и применения. Матер. науч. англ. Р 146 , 100644 (2021).
Google ученый
МакКаффри, В.П., Целльнер, Н.Э.Б., Вон, К.М., Беннетт, Э.Р. и Эрл, Э.К. Реакционная способность и живучесть гликолевого альдегида в экспериментах по моделированию удара метеорита. Происхождение Life Evol. Биосферы. 44 , 29–42 (2014).
ОБЪЯВЛЕНИЕ КАС Google ученый
Zellner, N.E.B., McCaffrey, VP & Butler, J.H.E. Кометный гликолевый альдегид как источник молекул пре-РНК.
Астробиология 20 , 1377–1388 (2020).ОБЪЯВЛЕНИЕ КАС пабмед Google ученый
Доусон А. и др. Влияние высокого давления на кристаллические структуры полиморфов глицина. Кристалл. Рост Des. 5 , 1415–1427 (2005).
КАС Google ученый
Герра Д., Гомес Л. А., Рестрепо А. и Дэвид Дж. Новые стабильные фазы кристаллов глицина. Хим. физ. 530 , 110645 (2021).
Google ученый
Парамесвари А., Премкумар С., Премкумар Р. и Милтон Франклин Бениал А. Рамановская спектроскопия с усилением поверхности и квантово-химические исследования монокристалла глицина. Дж. Мол. Структура 1116 , 180–187 (2016).
ОБЪЯВЛЕНИЕ КАС Google ученый
Хинтон, Дж. К. и др. Влияние давления на структуру и динамику решетки α-глицина: комбинированное экспериментальное и теоретическое исследование. CrystEngComm 21 , 4457–4464 (2019).
КАС Google ученый
Freeman, R. Справочник по ядерно-магнитному резонансу (Longman, 1987).
Google ученый
Эдалати, К. и др. Формирование метастабильных фаз в системе магний-титан методом кручения под высоким давлением и их способность аккумулировать водород. Acta Mater. 99 , 150–156 (2015).
ОБЪЯВЛЕНИЕ КАС Google ученый
Умеда Ю. и др. Выживаемость и реакционная способность глицина и аланина в ранних океанах: последствия ударов метеоритов. Дж. Биол. физ. 42 , 177–198 (2016).
КАС пабмед Google ученый
Бивер, Н. и др. Этиловый спирт и сахар в комете C/2014 Q2 (Лавджой). Науч. Доп. 1 , e1500863 (2015 г.).
ОБЪЯВЛЕНИЕ пабмед ПабМед Центральный Google ученый
Хадрауи, К. и др. Распределенный глицин в комете 67P/Чурюмова-Герасименко. Астрон. Астрофиз. 630 , А32 (2019).
КАС Google ученый
Найт, М. М. и др. Фотометрическое исследование комет Крейца, наблюдаемых SOHO с 1995 по 2005 год.
Астрон. J. 139 , 926 (2010).ОБЪЯВЛЕНИЕ КАС Google ученый
Мовшовиц, Н., Асфауг, Э. и Корыкански, Д. Численное моделирование разрушения кометы D/1993 F2 Шумейкера-Леви 9, представляющей прародителя в виде гравитационно связанного набора многогранников случайной формы. Астрофиз. J. 759 , 93 (2012).
ОБЪЯВЛЕНИЕ Google ученый
Триго-Родригес, Дж. М., Римола, А., Танбакуэй, С., Сото, В. К. и Ли, М. Аккреция воды в углеродистых хондритах: текущие данные и последствия для доставки воды на раннюю землю. Космические науки. 215 , 18 (2019).
ОБЪЯВЛЕНИЕ Google ученый
Окудэ С., Мацусима Ф., Кузе Х. и Симидзу Т. Молекулярно-лучевые исследования термического разложения глицина на твердых поверхностях.
япон. Дж. Заявл. физ. 26 , 627–632 (1987).ОБЪЯВЛЕНИЕ КАС Google ученый
Гоби, С., Фёрстель, М., Максютенко, П. и Кайзер, Р. И. Рефлектронное времяпролетное масс-спектрометрическое исследование путей разложения глицина на Марсе в присутствии перхлоратов и ионизирующего излучения. Астрофиз. J. 835 , 241 (2017).
ОБЪЯВЛЕНИЕ Google ученый
Классен, Дж. С. и Кебарле, П. Пороговые энергии диссоциации, индуцированной столкновением, протонированного глицина, глицинамида и некоторых родственных малых пептидов и аминоамидов пептидов. Дж. Ам. хим. соц. 119 , 6552–6563 (1997).
КАС Google ученый
Сато, Н., Даймон, Х. и Фуджи, К. Разложение глицина в воде при высокой температуре и высоком давлении.
Кагаку Когаку Ронбуншу 28 , 113–117 (2002).КАС Google ученый
Мохан Р. и др. Электрохимическое окисление глицина на биметаллических никель-марганцевых оксидных катализаторах. ХимЭлектроХим 7 , 561–568 (2020).
КАС Google ученый
Огура, К., Кобаяши, М., Накаяма, М. и Михо, Ю. Электрохимические и in situ FTIR исследования адсорбции и окисления глицина и лизина в щелочной среде. Дж. Электроанал. хим. 449 , 101–109 (1998).
КАС Google ученый
Hug, G.L. & Fessenden, R.W. Идентификация радикалов и определение их выхода при радиолитическом окислении глицина. Методика СОЭ с временным разрешением. J. Phys. хим. А 104 , 7021–7029 (2000 г.).
КАС Google ученый
Катао, А. Дж. Л. и Кастильо, А. Л. О пути деградации глифосата и глицина. Окружающая среда. науч. Процесс. Воздействия 20 , 1148–1157 (2018).
Google ученый
Weiss, I.M., Muth, C., Drumm, R. & Kirchner, H.O.K. Термическое разложение аминокислот глицина, цистеина, аспарагиновой кислоты, аспарагина, глутаминовой кислоты, глутамина, аргинина и гистидина. Биофиз. 11 , 2 (2018).
ПабМед ПабМед Центральный Google ученый
Триго-Родригес, Дж. М., Рубин, А. Э. и Уоссон, Дж.
Т. Нетуманное происхождение темных мантий вокруг хондр и включений в CM хондритах. Геохим. Космохим. Acta 70 , 1271–1290 (2006 г.).ОБЪЯВЛЕНИЕ КАС Google ученый
Rotelli, L. и др. Ключевая роль метеоритов в формировании соответствующих пребиотических молекул в формамидно-водной среде. науч. Респ. 6 , 38888 (2016).
ОБЪЯВЛЕНИЕ КАС пабмед ПабМед Центральный Google ученый
Кабедо, В., Льорка, Дж., Триго-Родригес, Дж. М. и Римола, А. Изучение реакций типа Фишера-Тропша на хондритовых метеоритах. Астрон. Астрофиз. 650 , А160 (2021).
ОБЪЯВЛЕНИЕ КАС Google ученый
 «>
«>Квенволден, К. и др. Доказательства внеземных аминокислот и углеводородов в метеорите Мерчисон. Природа 228 , 923–926 (1970).
ОБЪЯВЛЕНИЕ КАС пабмед Google ученый
 «>
«>Петерсон Э., Хорц Ф. и Чанг С. Модификация аминокислот при ударном давлении от 3,5 до 32 ГПа. Геохим. Космохим. Acta 61 , 3937–3950 (1997).
ОБЪЯВЛЕНИЕ КАС пабмед Google ученый
 Происхождение строительных блоков жизни: обзор. Геофизика. Фронт. 9 , 1117–1153 (2018).
Происхождение строительных блоков жизни: обзор. Геофизика. Фронт. 9 , 1117–1153 (2018). науч. 53 , 893–979 (2008).
науч. 53 , 893–979 (2008). П. Превращения нитрилов в твердой фазе при высоких давлениях в сочетании со сдвигом. Бык. акад. науч. дивизия СССР хим. науч. 21 , 813–818 (1972).
П. Превращения нитрилов в твердой фазе при высоких давлениях в сочетании со сдвигом. Бык. акад. науч. дивизия СССР хим. науч. 21 , 813–818 (1972). Матер. Рез. лат. 10 , 163–256 (2022).
Матер. Рез. лат. 10 , 163–256 (2022). «>
«>Эдалати, К. Металлургическая алхимия сверхсильной пластической деформацией посредством процесса кручения под высоким давлением. Матер. Транс. 60 , 1221–1229 (2019).
КАС Google ученый
 Астробиология 20 , 1377–1388 (2020).
Астробиология 20 , 1377–1388 (2020).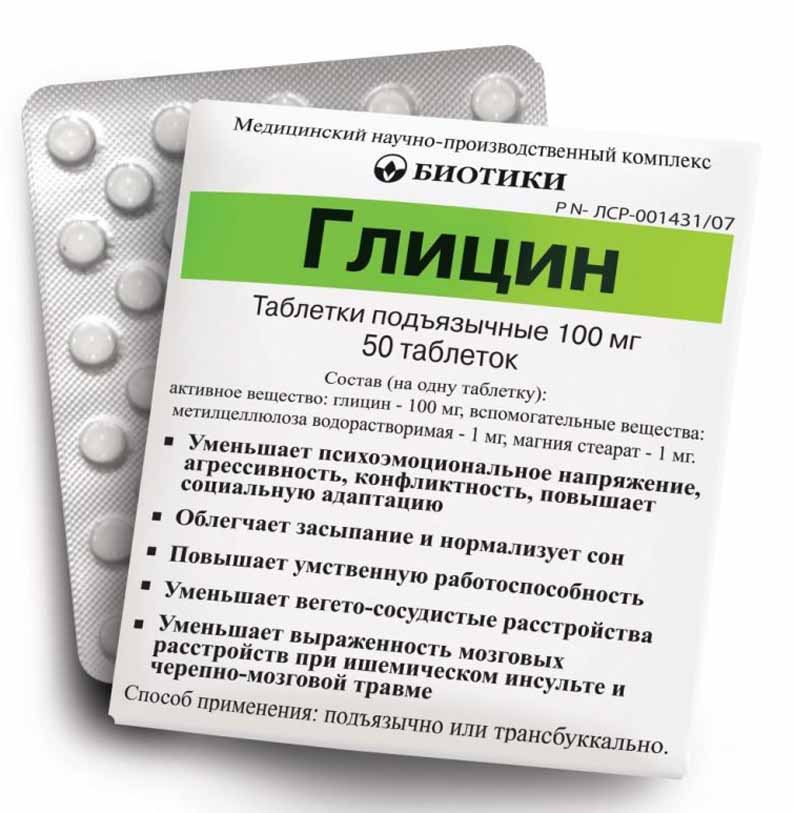 «>
«>Суровцев Н.В. и др. Глициновые фазы, образованные из замороженных водных растворов: пересмотрено. J. Chem. физ. 137 , 065103 (2012).
ОБЪЯВЛЕНИЕ КАС пабмед Google ученый

 Астрон. J. 139 , 926 (2010).
Астрон. J. 139 , 926 (2010). япон. Дж. Заявл. физ. 26 , 627–632 (1987).
япон. Дж. Заявл. физ. 26 , 627–632 (1987). Кагаку Когаку Ронбуншу 28 , 113–117 (2002).
Кагаку Когаку Ронбуншу 28 , 113–117 (2002). «>
«>O’Hair, R.A.J., Broughton, P.S., Styles, M.L., Frink, B.T. & Hadad, C.M. Пути фрагментации протонированного глицина: вычислительное исследование. Дж. Ам. соц. Масс-спектр. 11 , 687–696 (2000).
ПабМед Google ученый
 Т. Нетуманное происхождение темных мантий вокруг хондр и включений в CM хондритах. Геохим. Космохим. Acta 70 , 1271–1290 (2006 г.).
Т. Нетуманное происхождение темных мантий вокруг хондр и включений в CM хондритах. Геохим. Космохим. Acta 70 , 1271–1290 (2006 г.).Ссылки на скачивание
Благодарности
Эта работа частично поддерживается WPI-I2CNER, Япония, и частично Грантами в помощь научным исследованиям в инновационных областях от MEXT, Япония (19H05176 и 21H00150).
Author information
Authors and Affiliations
WPI International Institute for Carbon-Neutral Energy Research (WPI-I2CNER), Kyushu University, Fukuoka, Japan
Kaveh Edalati & Ikuo Taniguchi
School of Applied Sciences, Университет Кампинас (UNICAMP), Лимейра, Сан-Паулу, Бразилия
Рикардо Флориано и Аугусто Дукати Лучесси
Институт биологических наук, Государственный университет Сан-Паулу (UNESP), Рио-Кларо, Сан-Паулу, Бразилия
Augusto Ducati Luchessi
Авторы
- Kaveh Edalati
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
- Ikuo Taniguchi
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Scholar
- Ricardo Floriano
Просмотр публикаций автора
Вы также можете искать этого автора в PubMed Google Академия
- Augusto Ducati Luchessi
Посмотреть публикации автора
Вы также можете искать этого автора в PubMed Google Scholar
Contributions
KE, IT, RF, ADL: концептуализация, методология, проверка, письменная работа.
Автор, ответственный за переписку
Каве Эдалати.
Заявление об этике
Конкурирующие интересы
Авторы не заявляют об отсутствии конкурирующих интересов.
Дополнительная информация
Примечание издателя
Springer Nature остается нейтральной в отношении юрисдикционных претензий в опубликованных картах и институциональной принадлежности.
Права и разрешения
Открытый доступ Эта статья находится под лицензией Creative Commons Attribution 4.0 International License, которая разрешает использование, совместное использование, адаптацию, распространение и воспроизведение на любом носителе или в любом формате при условии, что вы укажете соответствующую ссылку на оригинальный автор(ы) и источник, предоставьте ссылку на лицензию Creative Commons и укажите, были ли внесены изменения. Изображения или другие сторонние материалы в этой статье включены в лицензию Creative Commons на статью, если иное не указано в кредитной строке материала.