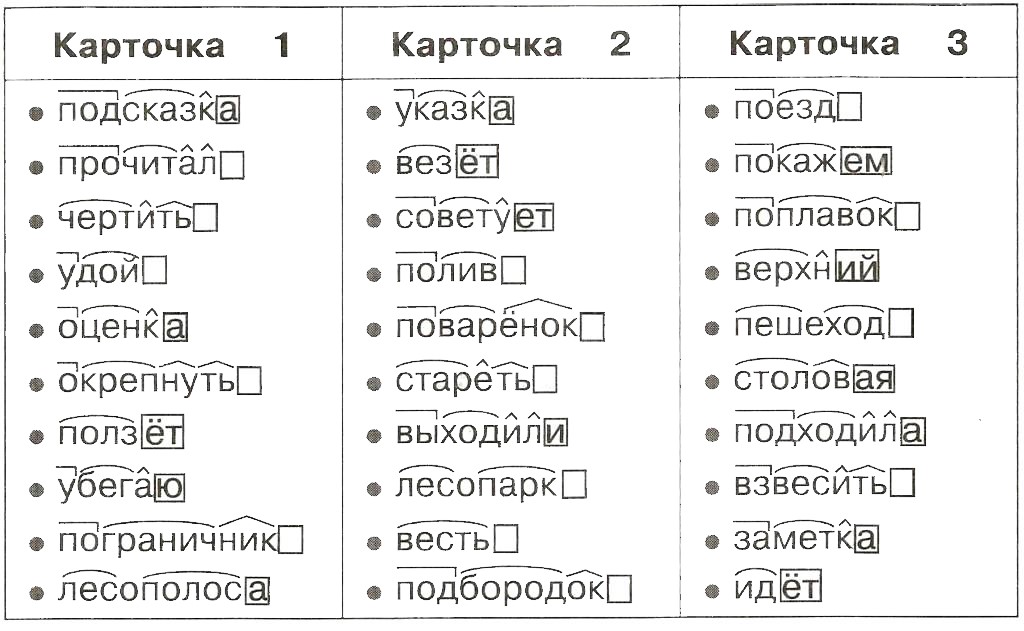
ГДЗ по Русскому языку 6 класс: Разумовская. Решебник
ГДЗ по русскому языку для 6 класса Разумовской – это сборник готовых домашних заданий по упражнениям из учебника, составленного известными российскими филологами – М.М. Разумовской, С.И. Львовой, В.И. Капинос и др. Его используют во многих общеобразовательных школах России.
Структура ГДЗ по учебнику русского языка шестого класса от Разумовской
Шестой класс – это время знакомства с главными частями речи, их морфологическим разбором и синтаксическим разбором предложений. Школьники учатся различать глухие и звонкие, мягкие и твердые согласные, формировать фонетическую структуру слова, определять лексическое значение. Они выявляют синонимы, омонимы и антонимы и строят с ними предложения.
ГДЗ по русскому языку 6 класса Разумовской, составленные на основе учебника 2018-2019 гг., включают в себя готовые упражнения, сочинения и изложения, а также разборы по темам:
- стили и типы речи;
- фонетика и орфоэпия;
- причастие и деепричастие;
- местоимение;
- лексика и фразеология;
- морфология и синтаксис;
- состав слова и словообразование;
- способы связи предложений в тексте;
- строение типовых фрагментов текста.

Ученики не только заучивают правила, но учатся грамотно писать и говорить с их использованием. Оттого для закрепления пройденных тем важно ознакомиться с порядком выполнения упражнений, составления разборов и их оформлением.
Готовые домашние задания для 6 класса на сайте ГДЗ Путина помогают не только шестиклассникам, но и их родителям, которые на их основе проверяют успеваемость своих детей и могут вместо репетитора разъяснить им порядок выполнения того или иного задания.
Решебники от ГДЗ Путина по русскому языку 6 класса к Разумовской
Выполненные упражнения и разборы по русскому языку, уникальные сочинения и эссе портала ГДЗ от Путина имеют ряд достоинств по сравнению с вариантами ответов из решебников иных сайтов:
- здесь собраны подробные синтаксические и морфологические разборы с пояснениями;
- аналогов уникальных творческих работ и сочинений ресурса нет в сети;
- все упражнения оформлены по требованиям Минобразования России;
- все материалы доступны в круглосуточном режиме и без регистрации.

Сайт ГДЗ от Путина можно открывать без затруднений с компьютера, телефона или планшета в любое удобное время. Школьникам это позволяет обращаться к ресурсу каждый раз при подготовке домашнего задания, родителям – при проверке работ их детей.
ГДЗ по русскому языку 6 класса к учебнику Разумовской настолько детально раскрывают порядок выполнения упражнений, что школьники могут не посещать репетиторов и дополнительные занятия – разобрать и запомнить применение сложных правил они могут и без их помощи.
Популярные решебники
ГДЗ по Русскому языку 6 класс: Ладыженская Т.А.
Издатель: М. Т. Баранов, Т.А. Ладыженская, Л. А. Тростенцова, 2015г. / 2019г.
ГДЗ по Русскому языку 6 класс: Разумовская М.М.
Издатель: М.М. Разумовская, С.И. Львова, В.И. Капинос. 2013-2019г.
Формирование, разборка и состав стрессовых гранул регулируются активностью АДФ-рибозилгидролазы альфавируса
. 2021 9 февраля; 118 (6): e2021719118.
2021 9 февраля; 118 (6): e2021719118.
doi: 10.1073/pnas.2021719118.
Аравинт Кумар Джаябалан 1 , Шриватсан Адиварахан 2 , Аакаш Коппула 3 , Рэйчи Абрахам 4 , Мона Батиш 3 5 , Даниэль Зенклузен 2 , Дайан Э. Гриффин 4 , Энтони К.Л. Леунг 6 7 8
Принадлежности
- 1 Кафедра биохимии и молекулярной биологии, Школа общественного здравоохранения Блумберга, Университет Джона Хопкинса, Балтимор, Мэриленд, 21205.
- 2 Департамент биохимии и молекулярной медицины, Университет Монреаля, Монреаль, QC h4T 1J4, Канада.
- 3 Факультет биологических наук, Делавэрский университет, Ньюарк, Делавэр, DE 19716.
- 4 W. Harry Feinstone Кафедра молекулярной микробиологии и иммунологии, Школа общественного здравоохранения Bloomberg, Университет Джона Хопкинса, Балтимор, MD 21205.
- 5 Кафедра медицинских и молекулярных наук, Делавэрский университет, Ньюарк, Делавэр, DE 19716.
- 6 Кафедра биохимии и молекулярной биологии, Школа общественного здравоохранения Блумберга, Университет Джона Хопкинса, Балтимор, Мэриленд, 21205; anthony. [email protected].
- 7 Кафедра молекулярной биологии и генетики, Медицинский факультет Университета Джона Хопкинса, Балтимор, Мэриленд, 21205.
- 8 Кафедра онкологии, Медицинский факультет, Университет Джона Хопкинса, Балтимор, Мэриленд, 21205.

- PMID: 33547245
- PMCID: PMC8017970
- DOI: 10.1073/пнас.2021719118
Бесплатная статья ЧВК
Аравинт Кумар Джаябалан и др.
Proc Natl Acad Sci U S A. .
.
Бесплатная статья ЧВК
. 2021 9 февраля; 118 (6): e2021719118.
doi: 10.1073/pnas.2021719118.Авторы
Аравинт Кумар Джаябалан 1 , Шриватсан Адиварахан 2 , Аакаш Коппула 3 , Рэйчи Абрахам 4 , Мона Батиш 3 5 , Даниэль Зенклузен 2 , Дайан Э. Гриффин 4 , Энтони К.Л. Леунг 6 7 8
Принадлежности
- 1 Кафедра биохимии и молекулярной биологии, Школа общественного здравоохранения Блумберга, Университет Джона Хопкинса, Балтимор, Мэриленд, 21205.
- 2 Департамент биохимии и молекулярной медицины, Университет Монреаля, Монреаль, QC h4T 1J4, Канада.
- 3 Факультет биологических наук, Делавэрский университет, Ньюарк, Делавэр, DE 19716.
- 4 W. Harry Feinstone Кафедра молекулярной микробиологии и иммунологии, Школа общественного здравоохранения Bloomberg, Университет Джона Хопкинса, Балтимор, MD 21205.
- 5 Кафедра медицинских и молекулярных наук, Делавэрский университет, Ньюарк, Делавэр, DE 19716.
- 6 Кафедра биохимии и молекулярной биологии, Школа общественного здравоохранения Блумберга, Университет Джона Хопкинса, Балтимор, Мэриленд, 21205; anthony. [email protected].
- 7 Кафедра молекулярной биологии и генетики, Медицинский факультет Университета Джона Хопкинса, Балтимор, Мэриленд, 21205.
- 8 Кафедра онкологии, Медицинский факультет, Университет Джона Хопкинса, Балтимор, Мэриленд, 21205.


- PMID: 33547245
- PMCID: PMC8017970
- DOI: 10.1073/пнас.2021719118
Абстрактный
Хотя биомолекулярные конденсаты стали важным биологическим явлением, механизмы, регулирующие их состав, и способы, которыми вирусы захватывают эти механизмы, остаются неясными. Альфавирусы, переносимые комарами, вызывают целый ряд заболеваний, от сыпи и артрита до энцефалита, при этом отсутствуют лицензированные лекарства для лечения или вакцины для профилактики. Неструктурный белок 3 фактора вирулентности альфавируса (nsP3) подавляет образование стрессовых гранул (SG) — класса цитоплазматических конденсатов, обогащенных факторами инициации трансляции и образующихся на ранней стадии инфекции. nsP3 имеет консервативный N-концевой макродомен, который гидролизует АДФ-рибозу из АДФ-рибозилированных белков, и С-концевой гипервариабельный домен, который связывает основной компонент SG G3BP1. Здесь мы показываем, что активность макродоменной гидролазы снижает АДФ-рибозилирование G3BP1, разбирает индуцированные вирусом SG и подавляет образование SG. Экспрессия nsP3 приводит к образованию отдельного класса конденсатов, в которых отсутствуют факторы инициации трансляции, но которые содержат G3BP1 и другие SG-ассоциированные РНК-связывающие белки. Экспрессия дефицитного по АДФ-рибозилгидролазе nsP3 приводит к образованию конденсатов, которые сохраняют факторы инициации трансляции, а также РНК-связывающие белки, сходные с SG.
Альфавирусы, переносимые комарами, вызывают целый ряд заболеваний, от сыпи и артрита до энцефалита, при этом отсутствуют лицензированные лекарства для лечения или вакцины для профилактики. Неструктурный белок 3 фактора вирулентности альфавируса (nsP3) подавляет образование стрессовых гранул (SG) — класса цитоплазматических конденсатов, обогащенных факторами инициации трансляции и образующихся на ранней стадии инфекции. nsP3 имеет консервативный N-концевой макродомен, который гидролизует АДФ-рибозу из АДФ-рибозилированных белков, и С-концевой гипервариабельный домен, который связывает основной компонент SG G3BP1. Здесь мы показываем, что активность макродоменной гидролазы снижает АДФ-рибозилирование G3BP1, разбирает индуцированные вирусом SG и подавляет образование SG. Экспрессия nsP3 приводит к образованию отдельного класса конденсатов, в которых отсутствуют факторы инициации трансляции, но которые содержат G3BP1 и другие SG-ассоциированные РНК-связывающие белки. Экспрессия дефицитного по АДФ-рибозилгидролазе nsP3 приводит к образованию конденсатов, которые сохраняют факторы инициации трансляции, а также РНК-связывающие белки, сходные с SG.
Ключевые слова: АДФ-рибозилирование; альфавирус; биомолекулярные конденсаты; макродомен; стрессовые гранулы.
Заявление о конфликте интересов
Авторы заявляют об отсутствии конкурирующих интересов.
Цифры
Рис. 1.
Два различных класса биомолекулярных…
Рис. 1.
При инфицировании альфавирусом образуются два различных класса биомолекулярных конденсатов.
При инфицировании альфавирусом образуются два различных класса биомолекулярных конденсатов. ( A ) Клетки U2OS, стабильно экспрессирующие GFP-eIF3g, были инфицированы WT CHIKV mCherry при MOI 10, подвергались визуализации живых клеток с интервалом времени 12 минут в течение 14 часов. Показанные изображения представляют собой снимки различных стадий заражения. ( B ) Клетки U2OS были либо ложно инфицированы, либо инфицированы CHIKV. Через двенадцать часов после инфицирования клетки лизировали и блоттировали против указанных антител. ( C ) Скрипичный график показывает распределение временных точек (минут), представляющих появление, исчезновение SG и появление конденсата nsP3. Были включены 97 клеток из трех независимых экспериментов, и в каждой клетке были измерены моменты времени появления/исчезновения СГ и появления конденсатов nsP3. Столбики погрешностей соответствуют SD.
Рис. 2.
Макродомен nsP3 подавляет образование СГ. …
…
Рис. 2.
Макродомен nsP3 подавляет образование СГ. ( A ) Схематическое изображение nsP3…
Рис. 2. Макродомен nsP3 подавляет образование СГ. ( A ) Схематическое представление архитектуры домена nsP3. Клетки U2OS, трансфицированные GFP-мечеными nsP3 MD , nsP3 ZBD или nsP3 HVD , либо обрабатывали 0,2 мМ арсенита отдельно, либо совместно обрабатывали 100 мкг/мл циклогексимида в течение 30 мин. Затем клетки обрабатывали и иммуноокрашивали на маркеры SG G3BP1 (красный) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. ( B ) Гистограмма показывает процент GFP-позитивных клеток с SG. *** P < 0,005, двусторонний, непарный тест Стьюдента t . Столбики погрешностей соответствуют среднему значению ± стандартное отклонение для трех независимых экспериментов. ( C ) Клетки U2OS, трансфицированные вектором GFP, nsP3 WT с меткой GFP или nsP3 AGDA , обрабатывали 0,2 мМ арсенита в течение 30 минут и подвергали иммуноокрашиванию на G3BP1 (красный) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. ( D ) Гистограмма показывает процент GFP-позитивных клеток с SG. * P < 0,05, *** P < 0,001, двусторонний, непарный тест Стьюдента t . Планки погрешностей соответствуют среднему значению ± стандартное отклонение с n = 3. (Шкала шкалы, 10 мкм.)
( C ) Клетки U2OS, трансфицированные вектором GFP, nsP3 WT с меткой GFP или nsP3 AGDA , обрабатывали 0,2 мМ арсенита в течение 30 минут и подвергали иммуноокрашиванию на G3BP1 (красный) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. ( D ) Гистограмма показывает процент GFP-позитивных клеток с SG. * P < 0,05, *** P < 0,001, двусторонний, непарный тест Стьюдента t . Планки погрешностей соответствуют среднему значению ± стандартное отклонение с n = 3. (Шкала шкалы, 10 мкм.)
Рис. 3.
АДФ-рибозилгидролазная активность nsP3 подавляет…
Рис. 3.
АДФ-рибозилгидролазная активность nsP3 подавляет образование СГ. ( A ) Трансфецированные клетки U2OS…
Рис. 3.
3. АДФ-рибозилгидролазная активность nsP3 подавляет образование SG. ( A ) Клетки U2OS, трансфицированные GFP-меченым nsP3 WT или другими точечными мутантами nsP3 (D10A, G32E, G32S, G112E и Y114A), обрабатывали 0,2 мМ арсенита в течение 30 минут и подвергали иммуноокрашиванию на G3BP1 (красный) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. ( B ) Клетки 293F трансфицировали либо вектором GFP, либо меченным GFP nsP3 WT , nsP3 G32E , nsP3 G32S или nsP3 Y114A . После 24-часовой трансфекции клетки осаждали, лизировали, иммунопреципитировали с использованием антител к GFP и проводили блоттинг иммунопреципитатов против антител G3BP1 и GFP. ( C ) Гистограмма показывает процент GFP-положительных клеток с SG в A . ( Нижний ) Связывание АДФ-рибозы и гидролазная активность протестированных мутантов (31). ** P < 0,01, двусторонний, непарный Стьюдента т тест. Столбики погрешностей соответствуют среднему значению ± стандартное отклонение с n = 3. ( D ) Клетки, трансфицированные GFP-мечеными nsP3 WT или nsP3 G32E , совместно обрабатывали 0,2 мМ арсенита и 100 мкг/мл циклогексимида для получения 30-минимина. Затем клетки подвергали иммуноокрашиванию на G3BP1 (красный) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. ( E ) В таблице обобщена колокализация nsP3 (WT или G32E) с различными связывающими мРНК белками и факторами трансляции, как показано на Приложение SI , рис. S3 A . ( F ) Микроскопия со сверхвысоким разрешением показывает, как распределение сигнала поли(А)+ мРНК перекрывается с мутантным nsP3 WT или G32E. Клетки U2OS, трансфицированные меченым GFP nsP3 WT или nsP3 G32E , обрабатывали 0,2 мМ арсенита в течение 30 минут и гибридизовали с олиго(dT) зондами с последующим окрашиванием на GFP. Зеленые и красные прямоугольники обозначают конденсаты из nsP3-трансфицированных и нетрансфицированных клеток соответственно.
Столбики погрешностей соответствуют среднему значению ± стандартное отклонение с n = 3. ( D ) Клетки, трансфицированные GFP-мечеными nsP3 WT или nsP3 G32E , совместно обрабатывали 0,2 мМ арсенита и 100 мкг/мл циклогексимида для получения 30-минимина. Затем клетки подвергали иммуноокрашиванию на G3BP1 (красный) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. ( E ) В таблице обобщена колокализация nsP3 (WT или G32E) с различными связывающими мРНК белками и факторами трансляции, как показано на Приложение SI , рис. S3 A . ( F ) Микроскопия со сверхвысоким разрешением показывает, как распределение сигнала поли(А)+ мРНК перекрывается с мутантным nsP3 WT или G32E. Клетки U2OS, трансфицированные меченым GFP nsP3 WT или nsP3 G32E , обрабатывали 0,2 мМ арсенита в течение 30 минут и гибридизовали с олиго(dT) зондами с последующим окрашиванием на GFP. Зеленые и красные прямоугольники обозначают конденсаты из nsP3-трансфицированных и нетрансфицированных клеток соответственно. (Шкала баров, 10 мкм.)
(Шкала баров, 10 мкм.)
Рис. 4.
Дифференциальная способность связывания PAR и АДФ-рибозилирование…
Рис. 4.
Дифференциальная ФАР-связывающая способность и АДФ-рибозилирование компонентов СГ. ( A ) 293F ячейки…
Рис. 4. Дифференциальная ФАР-связывающая способность и АДФ-рибозилирование компонентов СГ. ( 9Клетки 0179 A ) 293F трансфицировали либо вектором GFP, либо меченными GFP G3BP1, G3BP2, eIF3d, eIF3g или eIF3i в течение 36 часов. Затем клетки осаждали, лизировали и инкубировали со 100 пмоль биотина-PAR и гранулами стрептавидина. Затем пулдаун стрептавидина блоттировали антителами против GFP. ( B ) Клетки 293F, трансфицированные либо вектором GFP, либо мечеными GFP G3BP1, G3BP2, eIF3d, eIF3g или eIF3i в течение 36 часов. Затем клетки осаждали, лизировали и подвергали иммунопреципитации с использованием антител против GFP. Затем иммунопреципитаты блоттировали реагентом pan-ADPr.
Затем клетки осаждали, лизировали и подвергали иммунопреципитации с использованием антител против GFP. Затем иммунопреципитаты блоттировали реагентом pan-ADPr.
Рис. 5.
nsP3 снижает АДФ-рибозилирование…
Рис. 5.
nsP3 снижает АДФ-рибозилирование основного компонента SG G3BP1. ( А ) 293F…
Рис. 5. nsP3 снижает АДФ-рибозилирование основного компонента SG G3BP1. ( 9Клетки 0179 A ) 293F трансфицировали либо GFP, G3BP1, меченым GFP, либо котрансфицировали G3BP1, меченым GFP, с возрастающей концентрацией меченого FLAG nsP3 WT . После 24-часовой трансфекции клетки осаждали, лизировали, иммунопреципитировали с использованием антител против GFP и иммуноблоттировали с реагентом pan-ADPr. ( B ) Клетки 293F трансфицировали GFP, G3BP1, меченным GFP отдельно, или котрансфицировали G3BP1, меченым GFP, либо с меченым FLAG nsP3 WT , либо с nsP3 G32E . Через 24 часа после трансфекции клетки осаждали, лизировали, подвергали иммунопреципитации с использованием антител против GFP и иммуноблотингу с реагентом pan-ADPr. ( C ) Клетки 293F, трансфицированные меченным GFP G3BP1, лизировали и подвергали иммунопреципитации с использованием антител против GFP. Иммунопреципитаты разделяли на две половины и инкубировали либо с буфером, либо с 5 мкг ЧИКВ МД в течение 1 ч при 37 °С. После инкубации шарики промывали и промокали реагентом pan-ADPr. ( D ) Схематическое изображение доменной структуры G3BP1. ( E – G ) Клетки с двойным нокаутом (dKO) U2OS G3BP1/2 трансфицировали либо вектором GFP, либо конструкциями G3BP1, меченными GFP, в течение 36 часов. ( E ) Затем клетки либо лизировали, иммунопреципитировали с использованием антител против GFP и подвергали блотингу с реагентом pan-ADPr (звездочки обозначают тяжелую и легкую цепи), либо ( F ) пермеабилизировали, фиксировали и иммуноокрашивали для PABP (красный) и eIF3b (синий).
( B ) Клетки 293F трансфицировали GFP, G3BP1, меченным GFP отдельно, или котрансфицировали G3BP1, меченым GFP, либо с меченым FLAG nsP3 WT , либо с nsP3 G32E . Через 24 часа после трансфекции клетки осаждали, лизировали, подвергали иммунопреципитации с использованием антител против GFP и иммуноблотингу с реагентом pan-ADPr. ( C ) Клетки 293F, трансфицированные меченным GFP G3BP1, лизировали и подвергали иммунопреципитации с использованием антител против GFP. Иммунопреципитаты разделяли на две половины и инкубировали либо с буфером, либо с 5 мкг ЧИКВ МД в течение 1 ч при 37 °С. После инкубации шарики промывали и промокали реагентом pan-ADPr. ( D ) Схематическое изображение доменной структуры G3BP1. ( E – G ) Клетки с двойным нокаутом (dKO) U2OS G3BP1/2 трансфицировали либо вектором GFP, либо конструкциями G3BP1, меченными GFP, в течение 36 часов. ( E ) Затем клетки либо лизировали, иммунопреципитировали с использованием антител против GFP и подвергали блотингу с реагентом pan-ADPr (звездочки обозначают тяжелую и легкую цепи), либо ( F ) пермеабилизировали, фиксировали и иммуноокрашивали для PABP (красный) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. ( G ) Гистограмма показывает процент GFP-позитивных клеток с SG. нс, незначащий, двусторонний, непарный Стьюдента т тест. Столбики погрешностей соответствуют среднему значению ± стандартное отклонение с n = 3. ( H и I ) Клетки U2OS G3BP1/2 dKO котрансфицировали в двух комбинациях: 1) FLAG-вектор с G3BP1 или G3BP1-меченым ∆1– 168, и 2) nsP3 с маркировкой FLAG с G3BP1 или G3BP1 с меткой GFP ∆1–168. Через 36 ч после трансфекции клетки фиксировали и окрашивали на FLAG (красный) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. Гистограмма показывает процент GFP-положительных клеток с SG. *** Р < 0,001, **** P < 0,0001, двусторонний, непарный Критерий Стьюдента t . Планки погрешностей соответствуют среднему значению ± стандартное отклонение с n = 3. (Шкала шкалы, 10 мкм.)
Звездочки указывают на нетрансфицированные клетки. ( G ) Гистограмма показывает процент GFP-позитивных клеток с SG. нс, незначащий, двусторонний, непарный Стьюдента т тест. Столбики погрешностей соответствуют среднему значению ± стандартное отклонение с n = 3. ( H и I ) Клетки U2OS G3BP1/2 dKO котрансфицировали в двух комбинациях: 1) FLAG-вектор с G3BP1 или G3BP1-меченым ∆1– 168, и 2) nsP3 с маркировкой FLAG с G3BP1 или G3BP1 с меткой GFP ∆1–168. Через 36 ч после трансфекции клетки фиксировали и окрашивали на FLAG (красный) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. Гистограмма показывает процент GFP-положительных клеток с SG. *** Р < 0,001, **** P < 0,0001, двусторонний, непарный Критерий Стьюдента t . Планки погрешностей соответствуют среднему значению ± стандартное отклонение с n = 3. (Шкала шкалы, 10 мкм.)
Рис. 6.
6.
АДФ-рибозилгидролазная активность nsP3 подавляет…
Рис. 6.
АДФ-рибозилгидролазная активность nsP3 подавляет образование СГ при альфавирусной инфекции. ( А )…
Рис. 6. АДФ-рибозилгидролазная активность nsP3 подавляет образование SG при альфавирусной инфекции. ( A ) Титры вируса WT и мутанта Y114A в клетках NSC34 (31) и U2OS проанализированы при 24 hpi. Данные представлены в виде единиц образования бляшек (БОЕ)/мл ± SEM. ( B ) Клетки 293T, трансфицированные вектором GFP или GFP-G3BP1 в течение 12 часов, были либо ложно инфицированы, либо инфицированы CHIKV WT или CHIKV Y114A при МВД 1 в течение 24 часов. При 24 HPI клетки лизировали, иммунопреципитировали с использованием антител против GFP и блоттировали реагентом pan-ADPr. ( C и D ) Клетки U2OS инфицировали либо CHIKV WT , либо CHIKV Y114A при множественности множественности 1. При 5,5 hpi клетки обрабатывали 0,2 мМ арсенита в течение 30 минут и окрашивали иммуноокрашиванием на nsP3 (зеленый) , G3BP1 (красный) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. Гистограмма показывает процент инфицированных вирусом клеток (nsP3-позитивных) с SG. ** P < 0,01, двусторонний, непарный тест Стьюдента t . Столбики погрешностей соответствуют среднему значению ± стандартное отклонение с n = 3. ( E ) Клетки U2OS были инфицированы либо CHIKV WT , либо CHIKV Y114A при MOI 5. При 7,5 HPI клетки обрабатывали 0,2 мМ арсенита. в течение 30 мин. Затем обработанные клетки фиксировали и гибридизовали с зондами вРНК (красные) с последующим иммуноокрашиванием на G3BP1 (зеленый) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. ( Ф ) Прямоугольная диаграмма показывает процент клеток с вРНК , совместно локализованных с eIF3b/G3BP1 , на рис. 6 E и SI Приложение , рис.
При 5,5 hpi клетки обрабатывали 0,2 мМ арсенита в течение 30 минут и окрашивали иммуноокрашиванием на nsP3 (зеленый) , G3BP1 (красный) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. Гистограмма показывает процент инфицированных вирусом клеток (nsP3-позитивных) с SG. ** P < 0,01, двусторонний, непарный тест Стьюдента t . Столбики погрешностей соответствуют среднему значению ± стандартное отклонение с n = 3. ( E ) Клетки U2OS были инфицированы либо CHIKV WT , либо CHIKV Y114A при MOI 5. При 7,5 HPI клетки обрабатывали 0,2 мМ арсенита. в течение 30 мин. Затем обработанные клетки фиксировали и гибридизовали с зондами вРНК (красные) с последующим иммуноокрашиванием на G3BP1 (зеленый) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. ( Ф ) Прямоугольная диаграмма показывает процент клеток с вРНК , совместно локализованных с eIF3b/G3BP1 , на рис. 6 E и SI Приложение , рис. S5 E . Около 20 клеток из каждой временной точки были количественно определены для колокализации. * P < 0,05, нс, недостоверный, двусторонний, непарный критерий Стьюдента t . (Шкала, 10 мкм.)
S5 E . Около 20 клеток из каждой временной точки были количественно определены для колокализации. * P < 0,05, нс, недостоверный, двусторонний, непарный критерий Стьюдента t . (Шкала, 10 мкм.)
Рис. 7.
АДФ-рибозилгидролазная активность nsP3 регулирует…
Рис. 7.
АДФ-рибозилгидролазная активность nsP3 регулирует разборку SG. ( A ) Схематическое изображение…
Рис. 7. АДФ-рибозилгидролазная активность nsP3 регулирует разборку SG. ( A ) Схематическое изображение установки эксперимента по визуализации живых клеток. ( B и C ) Клетки U2OS, стабильно экспрессирующие GFP-eIF3g, были инфицированы WT или Y114A CHIKV 9.0007 mCherry с MOI 10 и подвергали визуализации живых клеток с интервалом времени 12 минут в течение 14 часов. Показанные изображения представляют собой моменты времени, когда SG не наблюдались (t = -12 мин), за которыми следует момент времени, когда наблюдались SG (t = 0 ч). Белые стрелки указывают на СГ. (Масштабная линейка, 10 мкм.) Скрипичный график показывает распределение среднего времени пребывания SG (минуты) в клетках, инфицированных WT и Y114A CHIKV mCherry . *** P < 0,001, двусторонний, непарный Стьюдента т тест. WT ( n = 51) и Y114A ( n = 77) инфицированные клетки из трех независимых экспериментов были включены для измерения времени пребывания SG. Столбики погрешностей соответствуют SD. ( D ) Модель сборки и разборки SG, индуцированной вирусной инфекцией, регулируемой активностью nsP3 ADP-рибозилгидролазы.
Показанные изображения представляют собой моменты времени, когда SG не наблюдались (t = -12 мин), за которыми следует момент времени, когда наблюдались SG (t = 0 ч). Белые стрелки указывают на СГ. (Масштабная линейка, 10 мкм.) Скрипичный график показывает распределение среднего времени пребывания SG (минуты) в клетках, инфицированных WT и Y114A CHIKV mCherry . *** P < 0,001, двусторонний, непарный Стьюдента т тест. WT ( n = 51) и Y114A ( n = 77) инфицированные клетки из трех независимых экспериментов были включены для измерения времени пребывания SG. Столбики погрешностей соответствуют SD. ( D ) Модель сборки и разборки SG, индуцированной вирусной инфекцией, регулируемой активностью nsP3 ADP-рибозилгидролазы.
См. это изображение и информацию об авторских правах в PMC
Похожие статьи
Роль Chikungunya nsP3 в регуляции активности G3BP1, формировании стрессовых гранул и эффективности лекарств.
Лу Х, Алам У, Уиллис С, Кеннеди Д. Лу X и др. Арх Мед Рез. 2021 Январь; 52(1):48-57. doi: 10.1016/j.arcmed.2020.10.002. Epub 2020 31 октября. Арх Мед Рез. 2021. PMID: 33131924
Компоненты стрессовых гранул G3BP1 и G3BP2 играют провирусную роль на ранней стадии репликации вируса чикунгунья.
Шольте Ф.Е., Тас А., Албулеску И.С., Жусинайте Э., Меритс А., Снайдер Э.Дж., ван Хемерт М.Дж. Шольте Ф.Е. и соавт. Дж Вирол. 2015 г., апрель 89(8):4457-69. doi: 10.1128/ОВИ.03612-14. Epub 2015 4 февраля. Дж Вирол. 2015. PMID: 25653451 Бесплатная статья ЧВК.
Отдельные домены G3BP способствуют эффективной кластеризации комплексов репликации альфавируса и задействованию аппарата инициации трансляции.
Гётте Б.
, Панас М.Д., Хеллстрем К., Лю Л., Самрин Б., Ларссон О., Ахола Т., Макинерни Г.М.
Гётте Б. и др.
PLoS Патог. 14 июня 2019 г .; 15 (6): e1007842. doi: 10.1371/journal.ppat.1007842. Электронная коллекция 2019 июнь.
PLoS Патог. 2019.
PMID: 31199850
Бесплатная статья ЧВК.Прогресс исследований структуры и функции G3BP.
Кан В, Ван И, Ян В, Чжан Дж, Чжэн Х, Ли Д. Канг В. и др. Фронт Иммунол. 2021 30 авг;12:718548. дои: 10.3389/fimmu.2021.718548. Электронная коллекция 2021. Фронт Иммунол. 2021. PMID: 34526993 Бесплатная статья ЧВК. Обзор.
Распутин десять лет спустя, а распущенность больше, чем когда-либо? Обзор G3BP.
Алам Ю., Кеннеди Д. Алам У и др. Biochim Biophys Acta Mol Cell Res.
2019 март; 1866 (3): 360-370. doi: 10.1016/j.bbamcr.2018.09.001. Epub 2018 5 сентября.
Biochim Biophys Acta Mol Cell Res. 2019.
PMID: 30595162
Бесплатная статья ЧВК.
Обзор.

 , Панас М.Д., Хеллстрем К., Лю Л., Самрин Б., Ларссон О., Ахола Т., Макинерни Г.М.
Гётте Б. и др.
PLoS Патог. 14 июня 2019 г .; 15 (6): e1007842. doi: 10.1371/journal.ppat.1007842. Электронная коллекция 2019 июнь.
PLoS Патог. 2019.
PMID: 31199850
Бесплатная статья ЧВК.
, Панас М.Д., Хеллстрем К., Лю Л., Самрин Б., Ларссон О., Ахола Т., Макинерни Г.М.
Гётте Б. и др.
PLoS Патог. 14 июня 2019 г .; 15 (6): e1007842. doi: 10.1371/journal.ppat.1007842. Электронная коллекция 2019 июнь.
PLoS Патог. 2019.
PMID: 31199850
Бесплатная статья ЧВК. 2019 март; 1866 (3): 360-370. doi: 10.1016/j.bbamcr.2018.09.001. Epub 2018 5 сентября.
Biochim Biophys Acta Mol Cell Res. 2019.
PMID: 30595162
Бесплатная статья ЧВК.
Обзор.
2019 март; 1866 (3): 360-370. doi: 10.1016/j.bbamcr.2018.09.001. Epub 2018 5 сентября.
Biochim Biophys Acta Mol Cell Res. 2019.
PMID: 30595162
Бесплатная статья ЧВК.
Обзор.Посмотреть все похожие статьи
Цитируется
Ингибирование cGAS ослабляет индуцированные Alu РНК иммунные ответы и цитотоксичность в пигментированном эпителии сетчатки.
Li J, Zhang F, Bian W, Chen Y, Liu J, Liu Z, Xiong Y, Wan X. Ли Дж. и др. Клетка Биоски. 2022 25 июля; 12 (1): 116. doi: 10.1186/s13578-022-00854-y. Клетка Биоски. 2022. PMID: 35879806 Бесплатная статья ЧВК.
Стрессовые шаги: прогресс и проблемы в понимании вызванной стрессом конденсации и накопления мРНК в стрессовых гранулах.
Glauninger H, Wong Hickernell CJ, Bard JAM, Drummond DA. Глаунингер Х. и др. Мол Ячейка. 2022 21 июля; 82 (14): 2544-2556. doi: 10.1016/j.molcel.2022.05.014. Epub 2022 3 июня. Мол Ячейка. 2022. PMID: 35662398 Обзор.
Роль стресс-гранул в подавлении репликации вируса эндорибонуклеазой вируса инфекционного бронхита.
Чжао Дж., Фэн Д., Чжао Й., Хуан М., Чжан С., Чжан Г. Чжао Дж. и др. Дж Вирол. 22 июня 2022 г .;96(12):e0068622. doi: 10.1128/jvi.00686-22. Epub 2022 31 мая. Дж Вирол. 2022. PMID: 35638780
Внутриклеточные моно-АДФ-рибозилтрансферазы в интерфазе вирус-хозяин.
Люшер Б., Верхейрстратен М., Криг С., Корн П. Люшер Б. и соавт. Cell Mol Life Sci. 2022 10 мая; 79 (6): 288.
doi: 10.1007/s00018-022-04290-6.
Cell Mol Life Sci. 2022.
PMID: 35536484
Бесплатная статья ЧВК.
Обзор.Активность поли-АДФ-рибозогидролазы макродомена Sindbis важна для синтеза вирусной РНК.
Агилар Э.Г., Паничча Г., Адура С., Сингер З.С., Эшбрук А.В., Разуки Б.С., Райс К.М., Макдональд М.Р. Агилар Э.Г. и соавт. Дж Вирол. 2022 13 апр;96(7):e0151621. doi: 10.1128/jvi.01516-21. Epub 2022 17 марта. Дж Вирол. 2022. PMID: 35297669 Бесплатная статья ЧВК.

 doi: 10.1007/s00018-022-04290-6.
Cell Mol Life Sci. 2022.
PMID: 35536484
Бесплатная статья ЧВК.
Обзор.
doi: 10.1007/s00018-022-04290-6.
Cell Mol Life Sci. 2022.
PMID: 35536484
Бесплатная статья ЧВК.
Обзор.Просмотреть все статьи «Цитируется по»
Типы публикаций
термины MeSH
вещества
Грантовая поддержка
- R01 GM104135/GM/NIGMS NIH HHS/США
- R56 AI137264/AI/NIAID NIH HHS/США
- PJT-148932/CIHR/Канада
- UDRF_SI_19A00244/CIHR/Канада
Серия композиций — Наама Цабар
Композиция 20 , 2016 г. — по настоящее время
— по настоящее время
Исполнение с 20 музыкантами, 20 усилителями и 3 песнями.
Куратор Сесилия Алемани и презентация High Line Art
Композиция 20 создает поле из 20 выступающих музыкантов, каждый из которых работает на своем усилителе, уникальным образом создавая 20 разрозненных источников звука. Музыканты разделены на три группы, которые исполняют собственные композиции: две песни были написаны специально для High Line, а третья представляла собой левацкую панк-песню 80-х на иврите.
Все три песни имеют одинаковую музыкальную структуру, одни и те же четыре аккорда, музыкальную гамму и BPM. В какой-то момент выступления все три оркестра играют одновременно. Благодаря их структурному сходству будет составлено плотное, но не совсем какофоническое музыкальное поле. По мере того, как песни перетекают одна в другую, коллективные смещающиеся силуэты музыкантов образуют скульптурную композицию, дополняющую звуковую. Для публики звук становится физическим, поскольку их движение по полю будет разбирать и собирать песни в музыкальном миксе, определяемом их положением и местоположением.
В композиции 20 приняли участие музыканты Роуз Бланшеи (Прима), Розана Кабан (Psychic Twin), Женевьев Чапин, Эндрю Дойч (HOME, Time Walk, Disbelief St.), Ароне Дайер (Букэ и Газе), Кристина Файлз, Синди Гуден. (Very Fresh), Майкл Уайт, Моника Хайдеманн (Heidemann), Аланна Хиггинс (MOON, Gem Spa), Лола Джонсон (Fisty, Quartascope), Эми Кария (BETS, Evil Daughter), Лайла Ларсон, Кэти Лау (Painted Zeros), Марисса Маззотта (Круги/Волны), Линдси-Пейдж МакКлой (Патио), Ава Мендоса (Неестественные пути), Кристин Мюллер, Линдси А. Пауэлл (Поле), Сэм Блейклок, Сара Регистр (Нормальный разговор), Джессика Сегал, Нога Шефи, и Таль Зубальский.
Фото: Лиз Лигон
Композиция 21 Наамы Цабар была организована и заказана Художественным музеем Нашера в Университете Дьюка, Дарем, Северная Каролина, 2019 г. Видеодокументация и производство Kidethnic.com. Работа © Наама Цабар. Видео © Nasher Museum of Art в Университете Дьюка.
Композиция 21 , 2019, Художественный музей Нашера, Северная Каролина, Сочинено и исполнено в сотрудничестве с Марией Олбани, Ширлетт Аммонс, Блэр Коппейдж, Хейли Фель, Л. А. Фей, Лео Фицпатрик, Джен Фу, Элизабет Хаммонд, Лора Кинг, Сус Лонг, Элисон Мартлью, Х.К. Макинтайр, Риз МакГенри, Синклер Палмер, Ким Регистр, Нора Роджерс, Сара Штраус, Камара Томас, Мара Томас, Сара Уорд и Дженни Уотерс.
А. Фей, Лео Фицпатрик, Джен Фу, Элизабет Хаммонд, Лора Кинг, Сус Лонг, Элисон Мартлью, Х.К. Макинтайр, Риз МакГенри, Синклер Палмер, Ким Регистр, Нора Роджерс, Сара Штраус, Камара Томас, Мара Томас, Сара Уорд и Дженни Уотерс.
Фотографии Дж. Колдевла. Изображения предоставлены Nasher Museum of Art
Композиция 21 , 2017, Prospect .4 New Orleans, Композиторы и исполнители Дана Эбботт, KC St. Afford, Дженн Аттауэй, Джаз Батлер, Джой Кларк, Элоиза Дэвис, Линн Друри, Хелен Джиллет, Ванда Джозеф, Месхия Лейк, Джина Мари Лесли, Келси Мэй, Тиффани Моррис, Джули Оделл, Сара Кинтана, Таше де ла Роша, Кэтрин Роуз, Маюми Шара, Кей Слотер, Сара Штраус и Боанна Траянова.
Фотографии Дж. Колдевла. Изображения предоставлены Prospect 4 New Orleans
Композиция 24 , 2006 г. – продолжается
Выступление с участием 24 музыкантов, 20 усилителей и 4 песен
Исполняется в Art Focus 5 , куратор Ами Барак и Бернар Блистене в музее Artzene 90906 , 9055 , , куратор Далия Левин.