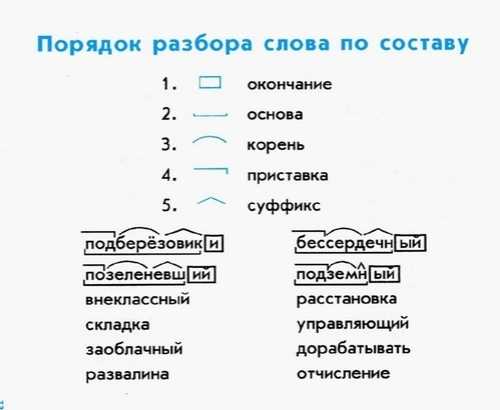
СУНЦ УрФУ
Расписание
Электронный журнал
Поступающим
Олимпиады, турниры, конкурсы
Планы работы
Подготовительные курсы
Новости:
05.05.2023
Заключительный этап. Успех!
Наши лицеисты достойно выступили на заключительном этапе Всероссийской олимпиады школьников.
04.05.2023
Успехи на международном форуме в Кыргызстане
Лицеисты привезли из солнечного Кыргызстана золотую и бронзовую медали международного форума «Мы — интеллектуалы XXI века!».
04.05.2023
Зарядись «Энергией будущего»!
Лицеисты СУНЦ с успехом выступили на всероссийском конкурсе научно-исследовательских работ и проектов.
25.04.2023
Время зарабатывать!
Соцэки СУНЦ совершенствуют свои практические навыки.
24.04.2023
На выборах победил лицей!
В СУНЦ УрФУ 20 апреля состоялись выборы в Совет лицеистов.
21.04.2023
Важный шаг на пути развития СУНЦев
В Москве состоялось заседание экспертного совета комитета по науке и высшему образованию Государственной Думы, посвященное развитию СУНЦев.
Больше новостей
Видеогалерея:
Мужчины СУНЦ о 8 Марта (2023)
Концерт к 8 Марта (2023)
Поздравление с Днем защитника Отечества (2023)
Больше видео
О нас:
Специализированный учебно-научный центр (СУНЦ) — структурное подразделение ФГАОУ ВО «УрФУ имени первого Президента России Б.Н. Ельцина», созданное в 1990 году как нетиповое структурное подразделение вуза, осуществляющее углубленное дифференцированное обучение по программам основного общего и среднего общего образования. Всего в России 10 СУНЦев. До мая 2011 года СУНЦ работал в составе Уральского государственного университета имени А. М. Горького (УрГУ).
В настоящее время СУНЦ имеет в своем составе 8 кафедр, укомплектованных профессорско-преподавательским составом УрФУ и учителями. Обучение производится по авторским программам, разработанным в соответствии с федеральными государственными образовательными стандартами; в составе СУНЦ — 8–11 классы различных профилей.
Иногородние обучающиеся проживают в уютном общежитии.
Прием производится в 8, 9, 10 и 11 классы. Работают подготовительные курсы.
Подробнее о правилах приема в СУНЦ можно узнать в отделе конкурсного отбора
по телефону +7 343 367-82-22 и в разделе нашего сайта «Поступающим».
Как нас найти:
Данилы Зверева ул., 30, Екатеринбург. N56°52´4˝ E60°39´16˝
Проезд:
- автобусами № 48, 52, 81 до остановки «Фирма Авангард»;
- автобусами № 28, 58 до остановки «Данилы Зверева», далее 7 минут пешком по улице Данилы Зверева;
- троллейбусом № 18 до остановки «Данилы Зверева», далее 14 минут пешком по улицам Сулимова, Данилы Зверева;
- троллейбусами № 4 до остановки «Сулимова», № 19, 32 до остановки «Боровая», далее 15 минут пешком по улицам Боровая, Вилонова, Данилы Зверева.
Ответов пока нет | ||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
Посмотреть всех экспертов из раздела Учеба и наука | ||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||


 2+3, x0=2;
2) f(x)=x(x+1), a=3.
Запишите уравнение касательной к графику функции в точке с
2+3, x0=2;
2) f(x)=x(x+1), a=3.
Запишите уравнение касательной к графику функции в точке сРазборка и повторная сборка кристаллов дифенилаланина путем выпаривания растворителя
Сохранить цитату в файл
Формат: Резюме (текст)PubMedPMIDAbstract (текст)CSV
Добавить в коллекции
- Создать новую коллекцию
- Добавить в существующую коллекцию
Назовите свою коллекцию:
Имя должно содержать менее 100 символов
Выберите коллекцию:
Не удалось загрузить вашу коллекцию из-за ошибки
Повторите попытку
Добавить в мою библиографию
- Моя библиография
Повторите попытку
Ваш сохраненный поиск
Название сохраненного поиска:
Условия поиска:
Тестовые условия поиска
Электронная почта: (изменить)
Который день? Первое воскресеньеПервый понедельникПервый вторникПервая средаПервый четвергПервая пятницаПервая субботаПервый деньПервый рабочий день
Который день? ВоскресеньеПонедельникВторникСредаЧетвергПятницаСуббота
Формат отчета: РезюмеРезюме (текст)АбстрактАбстракт (текст)PubMed
Отправить максимум:
1 шт. 5 шт. 10 шт. 20 шт. 50 шт. 100 шт. 200 шт.
5 шт. 10 шт. 20 шт. 50 шт. 100 шт. 200 шт.
Отправить, даже если нет новых результатов
Необязательный текст в электронном письме:
Создайте файл для внешнего программного обеспечения для управления цитированием
Полнотекстовые ссылки
Эльзевир Наука
Полнотекстовые ссылки
. 2021 Октябрь; 599: 661-666.
doi: 10.1016/j.jcis.2021.04.062. Epub 2021 17 апр.
Цзяруй Ся 1 , Бинбин Сан 1 , Ченлей Ван 1 , Нан Сун 1 , Хунцянь Цао 2 , И Цзя 1 , Ян Ян 3 , Джунбай Ли 4
Принадлежности
- 1 Пекинская национальная лаборатория молекулярных наук (BNLMS), Ключевая лаборатория CAS по коллоидной, межфазной и химической термодинамике, Институт химии Китайской академии наук, Пекин 100190, Китай; Университет Китайской академии наук, Пекин 100049, Китай.

- 2 Ключевая лаборатория CAS по биомедицинским эффектам наноматериалов и нанобезопасности, Национальный центр нанонауки и технологий, Пекин 100190 Китай; Школа общественного здравоохранения, Шаньдунский университет, Цзинань 250000, Китай.
- 3 Ключевая лаборатория CAS по биомедицинским эффектам наноматериалов и нанобезопасности, Национальный центр нанонауки и технологий, Пекин 100190 Китай. Электронный адрес: [email protected].
- 4 Пекинская национальная лаборатория молекулярных наук (BNLMS), Ключевая лаборатория CAS коллоидной, межфазной и химической термодинамики, Институт химии Китайской академии наук, Пекин 100190, Китай; Университет Китайской академии наук, Пекин 100049, Китай. Электронный адрес: jbli@iccas. ac.cn.

 ac.cn.
ac.cn.- PMID: 33979748
- DOI: 10.1016/j.jcis.2021.04.062
Цзяруи Ся и др. J Коллоидный интерфейс Sci. 2021 окт.
. 2021 Октябрь; 599: 661-666.
doi: 10.1016/j.jcis.2021.04.062. Epub 2021 17 апр.
Авторы
Цзяжуй Ся 1 , Бинбин Сан 1 , Ченлей Ван 1 , Нан Сун 1 , Хунцянь Цао 2 , И Цзя 1 , Ян Ян 3 , Джунбай Ли 4
Принадлежности
- 1 Пекинская национальная лаборатория молекулярных наук (BNLMS), Ключевая лаборатория CAS по коллоидной, межфазной и химической термодинамике, Институт химии Китайской академии наук, Пекин 100190, Китай; Университет Китайской академии наук, Пекин 100049, Китай.
- 2 Ключевая лаборатория CAS по биомедицинским эффектам наноматериалов и нанобезопасности, Национальный центр нанонауки и технологий, Пекин 100190 Китай; Школа общественного здравоохранения, Шаньдунский университет, Цзинань 250000, Китай.
- 3 Ключевая лаборатория CAS по биомедицинским эффектам наноматериалов и нанобезопасности, Национальный центр нанонауки и технологий, Пекин 100190 Китай. Электронный адрес: [email protected].
- 4 Пекинская национальная лаборатория молекулярных наук (BNLMS), Ключевая лаборатория CAS коллоидной, межфазной и химической термодинамики, Институт химии Китайской академии наук, Пекин 100190, Китай; Университет Китайской академии наук, Пекин 100049, Китай. Электронный адрес: jbli@iccas. ac.cn.

 ac.cn.
ac.cn.- PMID: 33979748
- DOI: 10.1016/j.jcis.2021.04.062
Абстрактный
Гипотеза: Кристаллические самосборки дифенилаланина (ФФ) уже давно считаются связанными с болезнью Альцгеймера. Лучшее понимание механизма формирования таких структур может привести к стратегиям исследования динамических процессов сборки и разборки FF.
Эксперимент: Сборка, разборка и повторная сборка кристаллов ФФ зависят от состава растворителя и могут быть вызваны испарением растворителя. В данной работе эти процессы непосредственно контролируются, а полученные структуры анализируются.
Выводы: Показана роль растворителя для сборки, разборки и повторной сборки кристаллов дифенилаланина. Исходная кристаллическая структура, образующаяся в результате самосборки мономеров ФФ, может быть преобразована в игольчатые кристаллы и далее в полые гексагональные микротрубки путем испарения растворителя. Показано, что все процессы сборки-разборки являются самопроизвольными и обусловлены термодинамикой. Обнаружено также, что некоторые кристаллические структуры обладают оптическими волноводными свойствами.
Ключевые слова: дифенилаланин; Разборка; Оптический волновод; Самостоятельная сборка; Термодинамика.
Copyright © 2021 Elsevier Inc. Все права защищены.
Заявление о конфликте интересов
 gov/pub-one»> Декларация о конкурирующих интересах Авторы заявляют, что у них нет известных конкурирующих финансовых интересов или личных отношений, которые могли бы повлиять на работу, представленную в этой статье.
gov/pub-one»> Декларация о конкурирующих интересах Авторы заявляют, что у них нет известных конкурирующих финансовых интересов или личных отношений, которые могли бы повлиять на работу, представленную в этой статье.Похожие статьи
Кинетика образования микро/нанотрубок L,L-дифенилаланина в зависимости от растворителя.
Bandeira CCS, Foiani LMC, Carlos GB, Ishikawa MS, Ferreira PMGL, da Silva Martinho H. Бандейра CCS и др. Phys Chem Chem Phys. 2023 8 февраля; 25 (6): 5107-5113. дои: 10.1039/d2cp03491c. Phys Chem Chem Phys. 2023. PMID: 36722992
Индуцированная температурой обратимая самосборка пептида дифенилаланина и структурный переход от органогеля к кристаллическим нанопроволокам.
Хуан Р, Ван И, Ци В, Су Р, Хэ З.
Хуанг Р. и др.
Nanoscale Res Lett. 3 декабря 2014 г .; 9 (1): 653. дои: 10.1186/1556-276X-9-653. Электронная коллекция 2014.
Nanoscale Res Lett. 2014.
PMID: 25520600
Бесплатная статья ЧВК.Направленная самосборка монокристалла дипептида в капилляре.
Сунь Б., Риглер Х., Дай Л., Эйкельманн С., Ли И., Ли Г., Ян И., Ли К., Фу М., Фей Дж., Ли Дж. Сан Б. и др. АКС Нано. 2018 27 февраля; 12 (2): 1934-1939. doi: 10.1021/acsnano.7b08925. Epub 2018 22 января. АКС Нано. 2018. PMID: 29337528
Кинетика, термодинамика и механизмы самосборки коротких ароматических пептидов.
Мейсон Т.О., Бьюэлл А.К. Мейсон Т.О. и соавт. Adv Exp Med Biol. 2019;1174:61-112.
doi: 10.1007/978-981-13-9791-2_3.
Adv Exp Med Biol. 2019.
PMID: 31713197
Обзор.Функциональные архитектуры, основанные на самосборке биоинспирированных дипептидов: структурная модуляция и ее фотоэлектронные приложения.
Чен С, Лю К, Ли Дж, Ян С. Чен С и др. Adv Коллоидный интерфейс Sci. 2015 ноябрь; 225: 177-93. doi: 10.1016/j.cis.2015.09.001. Epub 2015 5 сентября. Adv Коллоидный интерфейс Sci. 2015. PMID: 26365127 Обзор.
 Хуанг Р. и др.
Nanoscale Res Lett. 3 декабря 2014 г .; 9 (1): 653. дои: 10.1186/1556-276X-9-653. Электронная коллекция 2014.
Nanoscale Res Lett. 2014.
PMID: 25520600
Бесплатная статья ЧВК.
Хуанг Р. и др.
Nanoscale Res Lett. 3 декабря 2014 г .; 9 (1): 653. дои: 10.1186/1556-276X-9-653. Электронная коллекция 2014.
Nanoscale Res Lett. 2014.
PMID: 25520600
Бесплатная статья ЧВК. doi: 10.1007/978-981-13-9791-2_3.
Adv Exp Med Biol. 2019.
PMID: 31713197
Обзор.
doi: 10.1007/978-981-13-9791-2_3.
Adv Exp Med Biol. 2019.
PMID: 31713197
Обзор.Посмотреть все похожие статьи
Полнотекстовые ссылки
Эльзевир Наука
Укажите
Формат: ААД АПА МДА НЛМ
Отправить по номеру
Стресс-гранулы образование, разборка и состав регулируются активностью АДФ-рибозилгидролазы альфавируса
. 2021 9 февраля; 118 (6): e2021719118.
2021 9 февраля; 118 (6): e2021719118.
doi: 10.1073/pnas.2021719118.
Аравинт Кумар Джаябалан 1 , Шриватсан Адиварахан 2 , Аакаш Коппула 3 , Рэйчи Абрахам 4 , Мона Батиш 3 5 , Даниэль Зенклузен 2 , Дайан Э. Гриффин 4 , Энтони К.Л. Леунг 6 7 8
Принадлежности
- 1 Кафедра биохимии и молекулярной биологии, Школа общественного здравоохранения Блумберга, Университет Джона Хопкинса, Балтимор, Мэриленд, 21205.
- 2 Департамент биохимии и молекулярной медицины, Университет Монреаля, Монреаль, QC h4T 1J4, Канада.
- 3 Факультет биологических наук, Делавэрский университет, Ньюарк, Делавэр, 19716.
- 4 W. Harry Feinstone Кафедра молекулярной микробиологии и иммунологии, Школа общественного здравоохранения Bloomberg, Университет Джона Хопкинса, Балтимор, MD 21205.
- 5 Кафедра медицинских и молекулярных наук, Делавэрский университет, Ньюарк, Делавэр, DE 19716.
- 6 Кафедра биохимии и молекулярной биологии, Школа общественного здравоохранения Блумберга, Университет Джона Хопкинса, Балтимор, Мэриленд, 21205; anthony. [email protected].
- 7 Кафедра молекулярной биологии и генетики, Медицинский факультет Университета Джона Хопкинса, Балтимор, Мэриленд, 21205.
- 8 Кафедра онкологии, Медицинский факультет, Университет Джона Хопкинса, Балтимор, Мэриленд, 21205.


- PMID: 33547245
- PMCID: PMC8017970
- DOI: 10.1073/пнас.2021719118
Бесплатная статья ЧВК
Аравинт Кумар Джаябалан и др.
Proc Natl Acad Sci U S A. .
.
Бесплатная статья ЧВК
. 2021 9 февраля; 118 (6): e2021719118.
doi: 10.1073/pnas.2021719118.
Авторы
Аравинт Кумар Джаябалан 1 , Шриватсан Адиварахан 2 , Аакаш Коппула 3 , Рэйчи Абрахам 4 , Мона Батиш 3 5 , Даниэль Зенклузен 2 , Дайан Э. Гриффин 4 , Энтони К.Л. Леунг 6 7 8
Принадлежности
- 1 Кафедра биохимии и молекулярной биологии, Школа общественного здравоохранения Блумберга, Университет Джона Хопкинса, Балтимор, Мэриленд, 21205.
- 2 Департамент биохимии и молекулярной медицины, Университет Монреаля, Монреаль, QC h4T 1J4, Канада.
- 3 Факультет биологических наук, Делавэрский университет, Ньюарк, Делавэр, 19716.
- 4 W. Harry Feinstone Кафедра молекулярной микробиологии и иммунологии, Школа общественного здравоохранения Bloomberg, Университет Джона Хопкинса, Балтимор, MD 21205.
- 5 Кафедра медицинских и молекулярных наук, Делавэрский университет, Ньюарк, Делавэр, 19716.
- 6 Кафедра биохимии и молекулярной биологии, Школа общественного здравоохранения Блумберга, Университет Джона Хопкинса, Балтимор, Мэриленд, 21205; anthony. [email protected].
- 7 Кафедра молекулярной биологии и генетики, Медицинский факультет Университета Джона Хопкинса, Балтимор, Мэриленд, 21205.
- 8 Кафедра онкологии, Медицинский факультет, Университет Джона Хопкинса, Балтимор, Мэриленд, 21205.


- PMID: 33547245
- PMCID: PMC8017970
- DOI: 10.1073/пнас.2021719118
Абстрактный
Хотя биомолекулярные конденсаты стали важным биологическим явлением, механизмы, регулирующие их состав, и способы, которыми вирусы захватывают эти механизмы, остаются неясными. Альфавирусы, переносимые комарами, вызывают целый ряд заболеваний, от сыпи и артрита до энцефалита, при этом отсутствуют лицензированные лекарства для лечения или вакцины для профилактики. Неструктурный белок 3 фактора вирулентности альфавируса (nsP3) подавляет образование стрессовых гранул (SG) — класса цитоплазматических конденсатов, обогащенных факторами инициации трансляции и образующихся на ранней стадии инфекции. nsP3 имеет консервативный N-концевой макродомен, который гидролизует АДФ-рибозу из АДФ-рибозилированных белков, и С-концевой гипервариабельный домен, который связывает основной компонент SG G3BP1. Здесь мы показываем, что активность макродоменной гидролазы снижает АДФ-рибозилирование G3BP1, разбирает индуцированные вирусом SG и подавляет образование SG. Экспрессия nsP3 приводит к образованию отдельного класса конденсатов, в которых отсутствуют факторы инициации трансляции, но которые содержат G3BP1 и другие SG-ассоциированные РНК-связывающие белки. Экспрессия дефицитного по АДФ-рибозилгидролазе nsP3 приводит к образованию конденсатов, которые сохраняют факторы инициации трансляции, а также РНК-связывающие белки, сходные с SG.
Альфавирусы, переносимые комарами, вызывают целый ряд заболеваний, от сыпи и артрита до энцефалита, при этом отсутствуют лицензированные лекарства для лечения или вакцины для профилактики. Неструктурный белок 3 фактора вирулентности альфавируса (nsP3) подавляет образование стрессовых гранул (SG) — класса цитоплазматических конденсатов, обогащенных факторами инициации трансляции и образующихся на ранней стадии инфекции. nsP3 имеет консервативный N-концевой макродомен, который гидролизует АДФ-рибозу из АДФ-рибозилированных белков, и С-концевой гипервариабельный домен, который связывает основной компонент SG G3BP1. Здесь мы показываем, что активность макродоменной гидролазы снижает АДФ-рибозилирование G3BP1, разбирает индуцированные вирусом SG и подавляет образование SG. Экспрессия nsP3 приводит к образованию отдельного класса конденсатов, в которых отсутствуют факторы инициации трансляции, но которые содержат G3BP1 и другие SG-ассоциированные РНК-связывающие белки. Экспрессия дефицитного по АДФ-рибозилгидролазе nsP3 приводит к образованию конденсатов, которые сохраняют факторы инициации трансляции, а также РНК-связывающие белки, сходные с SG. Таким образом, наши данные показывают, что АДФ-рибозилирование контролирует состав биомолекулярных конденсатов, особенно локализацию факторов инициации трансляции, во время инфекции альфавирусом.
Таким образом, наши данные показывают, что АДФ-рибозилирование контролирует состав биомолекулярных конденсатов, особенно локализацию факторов инициации трансляции, во время инфекции альфавирусом.
Ключевые слова: АДФ-рибозилирование; альфавирус; биомолекулярные конденсаты; макродомен; стрессовые гранулы.
Заявление о конфликте интересов
Авторы заявляют об отсутствии конкурирующих интересов.
Цифры
Рис. 1.
Два различных класса биомолекулярных…
Рис. 1.
При инфицировании альфавирусом образуются два различных класса биомолекулярных конденсатов. ( А )…
( А )…
Во время заражения альфавирусом образуются два различных класса биомолекулярных конденсатов. ( A ) Клетки U2OS, стабильно экспрессирующие GFP-eIF3g, были инфицированы вирусом WT CHIKV mCherry при MOI 10, подвергались визуализации живых клеток с интервалом времени 12 минут в течение 14 часов. Показанные изображения представляют собой снимки различных стадий заражения. ( B ) Клетки U2OS были либо ложно инфицированы, либо инфицированы CHIKV. Через двенадцать часов после инфицирования клетки лизировали и блоттировали против указанных антител. ( C ) Скрипичный график показывает распределение временных точек (минут), представляющих появление, исчезновение SG и появление конденсата nsP3. Были включены 97 клеток из трех независимых экспериментов, и в каждой клетке были измерены моменты времени появления/исчезновения СГ и появления конденсатов nsP3. Столбики погрешностей соответствуют SD. ( D ) Клетки U2OS инфицировали CHIKV WT при множественности заражения 1. Через 6 ч после заражения (hpi) клетки фиксировали и иммуноокрашивали на nsP3 и определяли белки SG. ( E ) Клетки U2OS, трансфицированные GFP-меченым nsP3 WT , либо не подвергались стрессу, либо обрабатывались 100 мкг/мл циклогексимида (CHX), 0,2 мМ арсенита отдельно (ARS), либо совместно обрабатывались 0,2 мМ арсенита и 100 мкг/мл. циклогексимид (ARS+CHX) в течение 30 мин. Затем клетки фиксировали и иммуноокрашивали на G3BP1 (красный) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. ( F ) Гистограмма показывает процент GFP-положительных клеток с SG из трех независимых экспериментов. Столбики погрешностей соответствуют среднему значению ± стандартное отклонение. (Шкала, 10 мкм.)
( D ) Клетки U2OS инфицировали CHIKV WT при множественности заражения 1. Через 6 ч после заражения (hpi) клетки фиксировали и иммуноокрашивали на nsP3 и определяли белки SG. ( E ) Клетки U2OS, трансфицированные GFP-меченым nsP3 WT , либо не подвергались стрессу, либо обрабатывались 100 мкг/мл циклогексимида (CHX), 0,2 мМ арсенита отдельно (ARS), либо совместно обрабатывались 0,2 мМ арсенита и 100 мкг/мл. циклогексимид (ARS+CHX) в течение 30 мин. Затем клетки фиксировали и иммуноокрашивали на G3BP1 (красный) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. ( F ) Гистограмма показывает процент GFP-положительных клеток с SG из трех независимых экспериментов. Столбики погрешностей соответствуют среднему значению ± стандартное отклонение. (Шкала, 10 мкм.)
Рис. 2.
Макродомен nsP3 подавляет образование СГ. …
…
Рис. 2.
Макродомен nsP3 подавляет образование СГ. ( A ) Схематическое изображение nsP3…
Рис. 2. Макродомен nsP3 подавляет образование СГ. ( A ) Схематическое представление архитектуры домена nsP3. Клетки U2OS, трансфицированные GFP-мечеными nsP3 MD , nsP3 ZBD или nsP3 HVD , либо обрабатывали 0,2 мМ арсенита отдельно, либо совместно обрабатывали 100 мкг/мл циклогексимида в течение 30 мин. Затем клетки обрабатывали и иммуноокрашивали на маркеры SG G3BP1 (красный) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. ( B ) Гистограмма показывает процент GFP-позитивных клеток с SG. *** P < 0,005, двусторонний, непарный критерий Стьюдента t . Столбики погрешностей соответствуют среднему значению ± стандартное отклонение для трех независимых экспериментов. ( C ) Клетки U2OS, трансфицированные вектором GFP, nsP3 WT с меткой GFP или nsP3 AGDA , обрабатывали 0,2 мМ арсенита в течение 30 минут и подвергали иммуноокрашиванию на G3BP1 (красный) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. ( D ) Гистограмма показывает процент GFP-положительных клеток с SG. * P < 0,05, *** P < 0,001, двусторонний, непарный критерий Стьюдента t . Планки погрешностей соответствуют среднему значению ± стандартное отклонение с n = 3. (Шкала шкалы, 10 мкм.)
( C ) Клетки U2OS, трансфицированные вектором GFP, nsP3 WT с меткой GFP или nsP3 AGDA , обрабатывали 0,2 мМ арсенита в течение 30 минут и подвергали иммуноокрашиванию на G3BP1 (красный) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. ( D ) Гистограмма показывает процент GFP-положительных клеток с SG. * P < 0,05, *** P < 0,001, двусторонний, непарный критерий Стьюдента t . Планки погрешностей соответствуют среднему значению ± стандартное отклонение с n = 3. (Шкала шкалы, 10 мкм.)
Рис. 3.
АДФ-рибозилгидролазная активность nsP3 подавляет…
Рис. 3.
АДФ-рибозилгидролазная активность nsP3 подавляет образование SG. ( A ) Трансфецированные клетки U2OS…
Рис. 3.
3. АДФ-рибозилгидролазная активность nsP3 подавляет образование SG. ( A ) Клетки U2OS, трансфицированные GFP-меченым nsP3 WT или другими точечными мутантами nsP3 (D10A, G32E, G32S, G112E и Y114A), обрабатывали 0,2 мМ арсенита в течение 30 минут и подвергали иммуноокрашиванию на G3BP1 (красный) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. ( B ) Клетки 293F трансфицировали либо вектором GFP, либо меченным GFP nsP3 WT , nsP3 G32E , nsP3 G32S или nsP3 Y114A . После 24-часовой трансфекции клетки осаждали, лизировали, иммунопреципитировали с использованием антител к GFP и проводили блоттинг иммунопреципитатов против антител G3BP1 и GFP. ( C ) Гистограмма показывает процент GFP-положительных клеток с SG в A . ( Нижний ) Связывание АДФ-рибозы и гидролазная активность протестированных мутантов (31). ** P < 0,01, двусторонний, непарный Стьюдента т тест. Столбики ошибок соответствуют среднему значению ± стандартное отклонение с n = 3. ( D ) Клетки, трансфицированные GFP-меченым nsP3 WT или nsP3 G32E , совместно обрабатывали 0,2 мМ арсенита и 100 мкг/мл циклогема. ксимид в течение 30 мин. Затем клетки подвергали иммуноокрашиванию на G3BP1 (красный) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. ( E ) В таблице обобщена колокализация nsP3 (WT или G32E) с различными белками, связывающими мРНК, и факторами трансляции, как показано на Приложение SI , рис. S3 A . ( F ) Микроскопия со сверхвысоким разрешением показывает, как распределение сигнала поли(А)+ мРНК перекрывается с мутантным nsP3 WT или G32E. Клетки U2OS, трансфицированные GFP-мечеными nsP3 WT или nsP3 G32E , обрабатывали 0,2 мМ арсенита в течение 30 минут и гибридизовали с олиго(dT) зондами с последующим окрашиванием на GFP. Зеленые и красные прямоугольники обозначают конденсаты из nsP3-трансфицированных и нетрансфицированных клеток соответственно.
Столбики ошибок соответствуют среднему значению ± стандартное отклонение с n = 3. ( D ) Клетки, трансфицированные GFP-меченым nsP3 WT или nsP3 G32E , совместно обрабатывали 0,2 мМ арсенита и 100 мкг/мл циклогема. ксимид в течение 30 мин. Затем клетки подвергали иммуноокрашиванию на G3BP1 (красный) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. ( E ) В таблице обобщена колокализация nsP3 (WT или G32E) с различными белками, связывающими мРНК, и факторами трансляции, как показано на Приложение SI , рис. S3 A . ( F ) Микроскопия со сверхвысоким разрешением показывает, как распределение сигнала поли(А)+ мРНК перекрывается с мутантным nsP3 WT или G32E. Клетки U2OS, трансфицированные GFP-мечеными nsP3 WT или nsP3 G32E , обрабатывали 0,2 мМ арсенита в течение 30 минут и гибридизовали с олиго(dT) зондами с последующим окрашиванием на GFP. Зеленые и красные прямоугольники обозначают конденсаты из nsP3-трансфицированных и нетрансфицированных клеток соответственно. (Шкала баров, 10 мкм.)
(Шкала баров, 10 мкм.)
Рис. 4.
Дифференциальная способность связывания PAR и АДФ-рибозилирование…
Рис. 4.
Дифференциальная ФАР-связывающая способность и АДФ-рибозилирование компонентов СГ. ( A ) Элементы 293F…
Рис. 4. Дифференциальная ФАР-связывающая способность и АДФ-рибозилирование компонентов СГ. ( 9Клетки 0421 A ) 293F трансфицировали либо вектором GFP, либо меченными GFP G3BP1, G3BP2, eIF3d, eIF3g или eIF3i в течение 36 часов. Затем клетки осаждали, лизировали и инкубировали со 100 пмоль биотина-PAR и гранулами стрептавидина. Затем пулдаун стрептавидина блоттировали антителами против GFP. ( B ) Клетки 293F, трансфицированные либо вектором GFP, либо меченными GFP G3BP1, G3BP2, eIF3d, eIF3g или eIF3i в течение 36 часов. Затем клетки осаждали, лизировали и подвергали иммунопреципитации с использованием антител против GFP. Затем иммунопреципитаты блоттировали реагентом pan-ADPr.
Затем клетки осаждали, лизировали и подвергали иммунопреципитации с использованием антител против GFP. Затем иммунопреципитаты блоттировали реагентом pan-ADPr.
Рис. 5.
nsP3 снижает АДФ-рибозилирование…
Рис. 5.
nsP3 снижает АДФ-рибозилирование основного компонента SG G3BP1. ( А ) 293F…
Рис. 5. nsP3 снижает АДФ-рибозилирование основного компонента SG G3BP1. ( 9Клетки 0421 A ) 293F трансфицировали либо GFP, G3BP1, меченым GFP, либо котрансфицировали G3BP1, меченым GFP, с возрастающей концентрацией меченого FLAG nsP3 WT . После 24-часовой трансфекции клетки осаждали, лизировали, иммунопреципитировали с использованием антител против GFP и иммуноблоттировали с реагентом pan-ADPr. ( B ) Клетки 293F трансфицировали GFP, G3BP1, меченным GFP отдельно, или котрансфицировали G3BP1, меченым GFP, либо с FLAG-меченым nsP3 WT , либо с nsP3 G32E . Через 24 часа после трансфекции клетки осаждали, лизировали, подвергали иммунопреципитации с использованием антител против GFP и иммуноблотингу с реагентом pan-ADPr. ( C ) Клетки 293F, трансфицированные меченным GFP G3BP1, лизировали и подвергали иммунопреципитации с использованием антител против GFP. Иммунопреципитаты разделяли на две половины и инкубировали либо с буфером, либо с 5 мкг ЧИКВ МД в течение 1 ч при 37 °С. После инкубации шарики промывали и промокали реагентом pan-ADPr. ( D ) Схематическое изображение доменной структуры G3BP1. ( E – G ) Клетки U2OS G3BP1/2 с двойным нокаутом (dKO) трансфицировали либо вектором GFP, либо конструкциями G3BP1, меченными GFP, в течение 36 часов. ( E ) Затем клетки либо лизировали, иммунопреципитировали с использованием антител против GFP и блоттировали реагентом pan-ADPr (звездочки обозначают тяжелую и легкую цепи), либо ( F ) пермеабилизировали, фиксировали и иммуноокрашивали на PABP (красный) и eIF3b (синий).
( B ) Клетки 293F трансфицировали GFP, G3BP1, меченным GFP отдельно, или котрансфицировали G3BP1, меченым GFP, либо с FLAG-меченым nsP3 WT , либо с nsP3 G32E . Через 24 часа после трансфекции клетки осаждали, лизировали, подвергали иммунопреципитации с использованием антител против GFP и иммуноблотингу с реагентом pan-ADPr. ( C ) Клетки 293F, трансфицированные меченным GFP G3BP1, лизировали и подвергали иммунопреципитации с использованием антител против GFP. Иммунопреципитаты разделяли на две половины и инкубировали либо с буфером, либо с 5 мкг ЧИКВ МД в течение 1 ч при 37 °С. После инкубации шарики промывали и промокали реагентом pan-ADPr. ( D ) Схематическое изображение доменной структуры G3BP1. ( E – G ) Клетки U2OS G3BP1/2 с двойным нокаутом (dKO) трансфицировали либо вектором GFP, либо конструкциями G3BP1, меченными GFP, в течение 36 часов. ( E ) Затем клетки либо лизировали, иммунопреципитировали с использованием антител против GFP и блоттировали реагентом pan-ADPr (звездочки обозначают тяжелую и легкую цепи), либо ( F ) пермеабилизировали, фиксировали и иммуноокрашивали на PABP (красный) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. ( G ) Гистограмма показывает процент GFP-положительных клеток с SG. нс, незначащий, двусторонний, непарный Стьюдента т тест. Столбики погрешностей соответствуют среднему значению ± стандартное отклонение с n = 3. ( H и I ) Клетки U2OS G3BP1/2 dKO котрансфицировали в двух комбинациях: 1) FLAG-вектор с G3BP1 или G3BP1-меченым ∆1– 168, и 2) nsP3 с маркировкой FLAG с G3BP1 или G3BP1 с меткой GFP ∆1–168. Через 36 ч после трансфекции клетки фиксировали и окрашивали на FLAG (красный) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. Гистограмма показывает процент GFP-положительных клеток с SG. *** Р < 0,001, **** P < 0,0001, двусторонний, непарный Критерий Стьюдента t . Планки погрешностей соответствуют среднему значению ± стандартное отклонение с n = 3. (Шкала шкалы, 10 мкм.)
Звездочки указывают на нетрансфицированные клетки. ( G ) Гистограмма показывает процент GFP-положительных клеток с SG. нс, незначащий, двусторонний, непарный Стьюдента т тест. Столбики погрешностей соответствуют среднему значению ± стандартное отклонение с n = 3. ( H и I ) Клетки U2OS G3BP1/2 dKO котрансфицировали в двух комбинациях: 1) FLAG-вектор с G3BP1 или G3BP1-меченым ∆1– 168, и 2) nsP3 с маркировкой FLAG с G3BP1 или G3BP1 с меткой GFP ∆1–168. Через 36 ч после трансфекции клетки фиксировали и окрашивали на FLAG (красный) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. Гистограмма показывает процент GFP-положительных клеток с SG. *** Р < 0,001, **** P < 0,0001, двусторонний, непарный Критерий Стьюдента t . Планки погрешностей соответствуют среднему значению ± стандартное отклонение с n = 3. (Шкала шкалы, 10 мкм.)
Рис. 6.
6.
АДФ-рибозилгидролазная активность nsP3 подавляет…
Рис. 6.
АДФ-рибозилгидролазная активность nsP3 подавляет образование СГ при альфавирусной инфекции. ( А )…
Рис. 6. АДФ-рибозилгидролазная активность nsP3 подавляет образование SG при альфавирусной инфекции. ( A ) Титры вируса WT и мутанта Y114A в клетках NSC34 (31) и U2OS проанализированы при 24 hpi. Данные представлены в виде единиц образования бляшек (БОЕ)/мл ± SEM. ( B ) Клетки 293T, трансфицированные вектором GFP или GFP-G3BP1 в течение 12 часов, были либо ложно инфицированы, либо инфицированы CHIKV WT или CHIKV Y114A при МВД 1 в течение 24 часов. При 24 HPI клетки лизировали, иммунопреципитировали с использованием антител против GFP и блоттировали реагентом pan-ADPr. ( C и D ) Клетки U2OS инфицировали либо CHIKV WT , либо CHIKV Y114A при множественности множественности 1. При 5,5 hpi клетки обрабатывали 0,2 мМ арсенита в течение 30 минут и окрашивали иммуноокрашиванием на nsP3 ( зеленый) , G3BP1 (красный) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. Гистограмма показывает процент инфицированных вирусом клеток (nsP3-позитивных) с SG. ** P < 0,01, двусторонний, непарный критерий Стьюдента t . Столбики погрешностей соответствуют среднему значению ± стандартное отклонение с n = 3. ( E ) Клетки U2OS были инфицированы либо CHIKV WT , либо CHIKV Y114A при MOI 5. При 7,5 HPI клетки обрабатывали 0,2 мМ арсенита. в течение 30 мин. Затем обработанные клетки фиксировали и гибридизовали с зондами вРНК (красные) с последующим иммуноокрашиванием на G3BP1 (зеленый) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. ( Ф ) Прямоугольная диаграмма показывает процент клеток с вРНК , совместно локализованных с eIF3b/G3BP1 , на рис. 6 E и SI Приложение , рис.
При 5,5 hpi клетки обрабатывали 0,2 мМ арсенита в течение 30 минут и окрашивали иммуноокрашиванием на nsP3 ( зеленый) , G3BP1 (красный) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. Гистограмма показывает процент инфицированных вирусом клеток (nsP3-позитивных) с SG. ** P < 0,01, двусторонний, непарный критерий Стьюдента t . Столбики погрешностей соответствуют среднему значению ± стандартное отклонение с n = 3. ( E ) Клетки U2OS были инфицированы либо CHIKV WT , либо CHIKV Y114A при MOI 5. При 7,5 HPI клетки обрабатывали 0,2 мМ арсенита. в течение 30 мин. Затем обработанные клетки фиксировали и гибридизовали с зондами вРНК (красные) с последующим иммуноокрашиванием на G3BP1 (зеленый) и eIF3b (синий). Звездочки указывают на нетрансфицированные клетки. ( Ф ) Прямоугольная диаграмма показывает процент клеток с вРНК , совместно локализованных с eIF3b/G3BP1 , на рис. 6 E и SI Приложение , рис.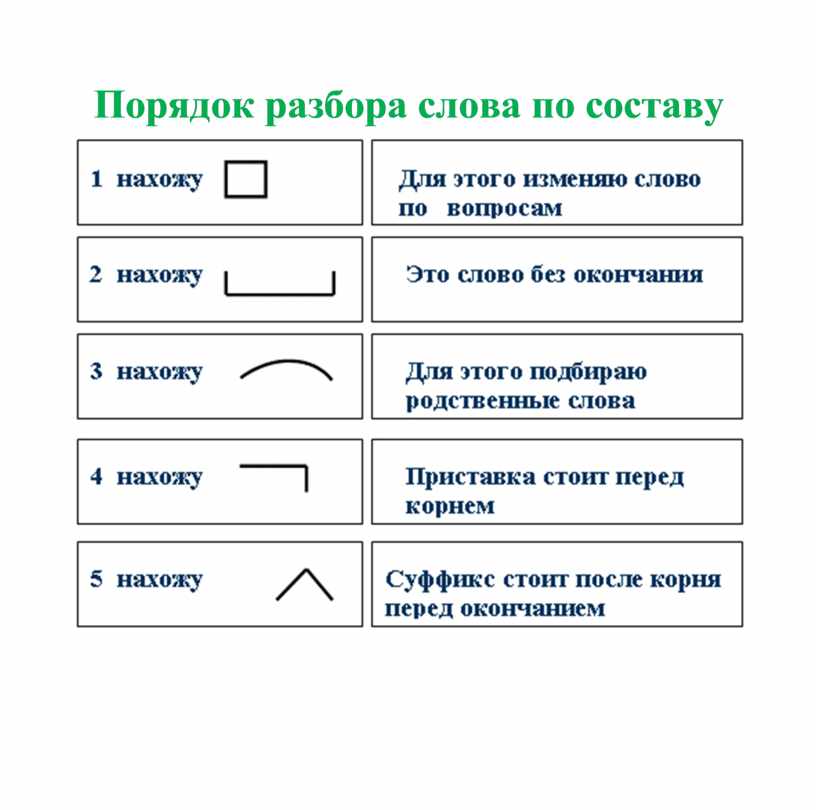 S5 E . Около 20 клеток из каждой временной точки были количественно определены для колокализации. * P < 0,05, нс, недостоверный, двусторонний, непарный критерий Стьюдента t . (Шкала, 10 мкм.)
S5 E . Около 20 клеток из каждой временной точки были количественно определены для колокализации. * P < 0,05, нс, недостоверный, двусторонний, непарный критерий Стьюдента t . (Шкала, 10 мкм.)
Рис. 7.
АДФ-рибозилгидролазная активность nsP3 регулирует…
Рис. 7.
АДФ-рибозилгидролазная активность nsP3 регулирует разборку SG. ( A ) Схематическое изображение…
Рис. 7. АДФ-рибозилгидролазная активность nsP3 регулирует разборку SG. ( A ) Схематическое изображение экспериментальной установки для визуализации живых клеток. ( B и C ) Клетки U2OS, стабильно экспрессирующие GFP-eIF3g, были инфицированы WT или Y114A CHIKV mCherry с MOI 10 и подвергали визуализации живых клеток с интервалом времени 12 минут в течение 14 часов.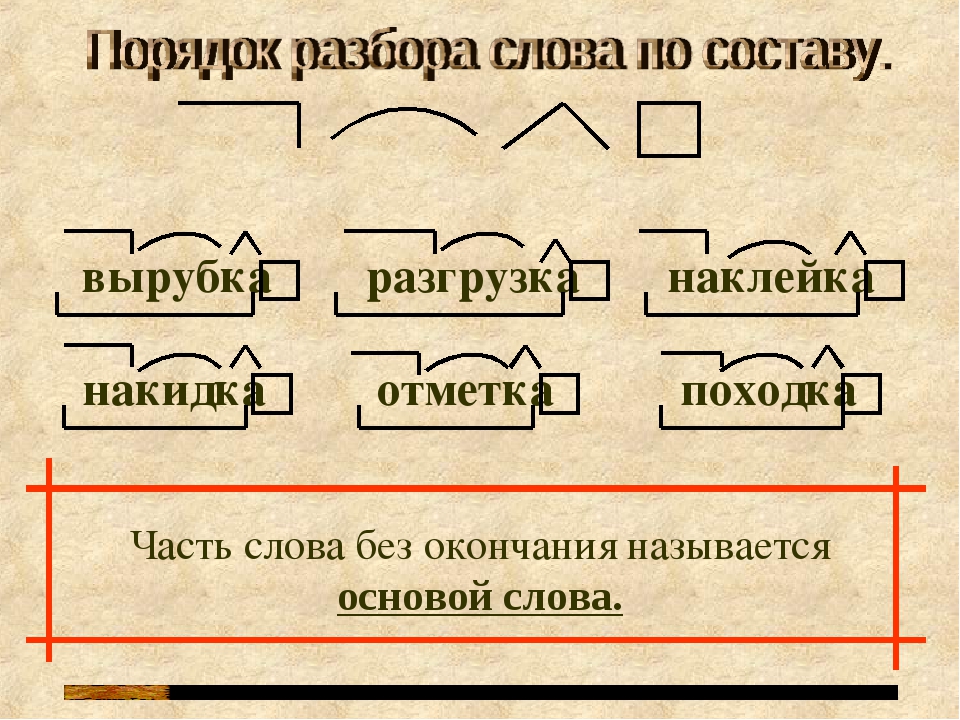 Показанные изображения представляют собой моменты времени, когда SG не наблюдались (t = -12 мин), за которыми следует момент времени, когда наблюдались SG (t = 0 ч). Белые стрелки указывают на СГ. (Масштабная линейка, 10 мкм.) Скрипичный график показывает распределение среднего времени пребывания SG (минуты) в клетках, инфицированных WT и Y114A CHIKV mCherry . *** P < 0,001, двусторонний, непарный Стьюдента т тест. WT ( n = 51) и Y114A ( n = 77) инфицированные клетки из трех независимых экспериментов были включены для измерения времени пребывания SG. Столбики погрешностей соответствуют SD. ( D ) Модель сборки и разборки SG, индуцированной вирусной инфекцией, регулируемой активностью nsP3 ADP-рибозилгидролазы.
Показанные изображения представляют собой моменты времени, когда SG не наблюдались (t = -12 мин), за которыми следует момент времени, когда наблюдались SG (t = 0 ч). Белые стрелки указывают на СГ. (Масштабная линейка, 10 мкм.) Скрипичный график показывает распределение среднего времени пребывания SG (минуты) в клетках, инфицированных WT и Y114A CHIKV mCherry . *** P < 0,001, двусторонний, непарный Стьюдента т тест. WT ( n = 51) и Y114A ( n = 77) инфицированные клетки из трех независимых экспериментов были включены для измерения времени пребывания SG. Столбики погрешностей соответствуют SD. ( D ) Модель сборки и разборки SG, индуцированной вирусной инфекцией, регулируемой активностью nsP3 ADP-рибозилгидролазы.
См. это изображение и информацию об авторских правах в PMC
Похожие статьи
Провирусная и противовирусная роль РНК-связывающего белка G3BP1.
Джаябалан А.К., Гриффин Д.Э., Леунг А.К.Л. Джаябалан А.К. и др. Вирусы. 2023 6 февраля; 15 (2): 449. дои: 10.3390/v15020449. Вирусы. 2023. PMID: 36851663 Бесплатная статья ЧВК. Обзор.
Роль Chikungunya nsP3 в регуляции активности G3BP1, формировании стрессовых гранул и эффективности лекарств.
Лу X, Алам Ю, Уиллис С, Кеннеди Д. Лу X и др. Арх Мед Рез. 2021 Январь; 52(1):48-57. doi: 10.1016/j.arcmed.2020.10.002. Epub 2020 31 октября. Арх Мед Рез. 2021. PMID: 33131924
Компоненты стрессовых гранул G3BP1 и G3BP2 играют провирусную роль на ранней стадии репликации вируса чикунгунья.
Шольте Ф.Е., Тас А., Албулеску И.С., Жусинайте Э.
, Меритс А., Снайдер Э.Дж., ван Хемерт М.Дж.
Шольте Ф.Е. и соавт.
Дж Вирол. 2015 апр;89(8):4457-69. doi: 10.1128/ОВИ.03612-14. Epub 2015 4 февраля.
Дж Вирол. 2015.
PMID: 25653451
Бесплатная статья ЧВК.Отдельные домены G3BP способствуют эффективной кластеризации комплексов репликации альфавируса и задействованию аппарата инициации трансляции.
Гётте Б., Панас М.Д., Хеллстрем К., Лю Л., Самрин Б., Ларссон О., Ахола Т., Макинерни Г.М. Гётте Б. и др. PLoS Патог. 201914 июня; 15 (6): e1007842. doi: 10.1371/journal.ppat.1007842. Электронная коллекция 2019 июнь. PLoS Патог. 2019. PMID: 31199850 Бесплатная статья ЧВК.
Прогресс исследований структуры и функции G3BP.
Кан В, Ван И, Ян В, Чжан Дж, Чжэн Х, Ли Д.
Канг В. и др.
Фронт Иммунол. 2021 30 авг;12:718548. doi: 10.3389/fimmu.2021.718548. Электронная коллекция 2021.
Фронт Иммунол. 2021.
PMID: 34526993
Бесплатная статья ЧВК.
Обзор.
 , Меритс А., Снайдер Э.Дж., ван Хемерт М.Дж.
Шольте Ф.Е. и соавт.
Дж Вирол. 2015 апр;89(8):4457-69. doi: 10.1128/ОВИ.03612-14. Epub 2015 4 февраля.
Дж Вирол. 2015.
PMID: 25653451
Бесплатная статья ЧВК.
, Меритс А., Снайдер Э.Дж., ван Хемерт М.Дж.
Шольте Ф.Е. и соавт.
Дж Вирол. 2015 апр;89(8):4457-69. doi: 10.1128/ОВИ.03612-14. Epub 2015 4 февраля.
Дж Вирол. 2015.
PMID: 25653451
Бесплатная статья ЧВК. Канг В. и др.
Фронт Иммунол. 2021 30 авг;12:718548. doi: 10.3389/fimmu.2021.718548. Электронная коллекция 2021.
Фронт Иммунол. 2021.
PMID: 34526993
Бесплатная статья ЧВК.
Обзор.
Канг В. и др.
Фронт Иммунол. 2021 30 авг;12:718548. doi: 10.3389/fimmu.2021.718548. Электронная коллекция 2021.
Фронт Иммунол. 2021.
PMID: 34526993
Бесплатная статья ЧВК.
Обзор.Посмотреть все похожие статьи
Цитируется
Провирусная и противовирусная роль РНК-связывающего белка G3BP1.
Джаябалан А.К., Гриффин Д.Э., Леунг А.К.Л. Джаябалан А.К. и др. Вирусы. 2023 6 февраля; 15 (2): 449. дои: 10.3390/v15020449. Вирусы. 2023. PMID: 36851663 Бесплатная статья ЧВК. Обзор.
Идентификация белков комаров, которые по-разному взаимодействуют с неструктурным белком 3 альфавируса, определяющим специфичность вектора.
Байерс Н.М., Бернс П.Л., Стучлик О., Рид М.С., Ледерманн Д.П., Пол Дж., Пауэрс А.М. Байерс Н.М. и соавт. PLoS Negl Trop Dis. 2023 25 января; 17 (1): e0011028. doi: 10.1371/journal.pntd.0011028. Электронная коллекция 2023 янв. PLoS Negl Trop Dis. 2023. PMID: 36696390 Бесплатная статья ЧВК.
Роль макродомена nsP3, зависящая от типа клеток, для связывания АДФ-рибозы и активности гидролазы при заражении вирусом чикунгунья.
Ким Т., Абрахам Р., Питерс Л., Йе Дж. Х., Гриффин Д. Э. Ким Т и др. Вирусы. 2022 9 декабря; 14 (12): 2744. дои: 10.3390/v14122744. Вирусы. 2022. PMID: 36560748 Бесплатная статья ЧВК.
Отдельная фаза? Поли(АДФ-рибоза) против РНК в организации биомолекулярных конденсатов.

