Определение, фонетический (звуко-буквенный) разбор и разбор слова по составу
На данной странице представлено лексическое значение слова «мягкий», а также сделан звуко-буквенный разбор и разбор слова по составу с транскрипцией и ударениями.Оглавление:
- Значение слова
- Звуко-буквенный разбор
- Разбор по составу
Значение слова
МЯГКИЙ [хк], ая, ое; гок, гка, гко, гки и гки; мягче; мягчайший.
1. Легко поддающийся давлению, сжатию, малоупругий, эластичный. м. хлеб. М. диван. Мягкая шерсть. Мягкие волосы. Мягкая обувь.
2. Приятный при ощущении, не раздражающий. м. свет. М. голос.
3. Плавный, размеренный. Мягкие движения. Мягко (нареч.) ступать. Мягкая посадка (о летательном аппарате: при минимальной скорости).
4. Кроткий, лишённый грубости, резкости. м. характер. Сделать замечание в мягкой форме.
5. Не очень строгий, снисходительный, не суровый. м. приговор.
6. Тёплый, приятный. м. климат.
7. О воде: содержащий мало солей кальция и магния, легко смывающий мыло, не жёсткий (в 5 знач.).
8. полн. ф. О транспортных средствах: с мягкими сидениями или предназначенный для использования мягких сидений. м. вагон. Мягкая плацкарта.
9. полн. ф. О согласных звуках: произносимый с приближением средней части языка к твёрдому нёбу; противоп. твёрдый (в 5 знач.) (спец.).
• Мягкий знак название буквы «ь».
| уменьш. мяконький, ая, ое (к 1 знач.) и мягонький, ая, ое (к 1 знач.).
| сущ. мягкость, и, ж.
Фонетический (звуко-буквенный) разбор
мя́гкий
мягкий — слово из 2 слогов: мя-гкий. Ударение падает на 1-й слог.
Транскрипция слова: [м’ахк’ий’]
м — [м’] — согласный, звонкий непарный, сонорный (всегда звонкий), мягкий (парный)
я — [а] — гласный, ударный
г — [х] — согласный, глухой непарный, твёрдый (парный)
к — [к’] — согласный, глухой парный, мягкий (парный)
и — [и] — гласный, безударный
й — [й’] — согласный, звонкий непарный, сонорный (всегда звонкий), мягкий (непарный, всегда произносится мягко)
В слове 6 букв и 6 звуков.
Цветовая схема: мягкий
Ударение в слове проверено администраторами сайта и не может быть изменено.
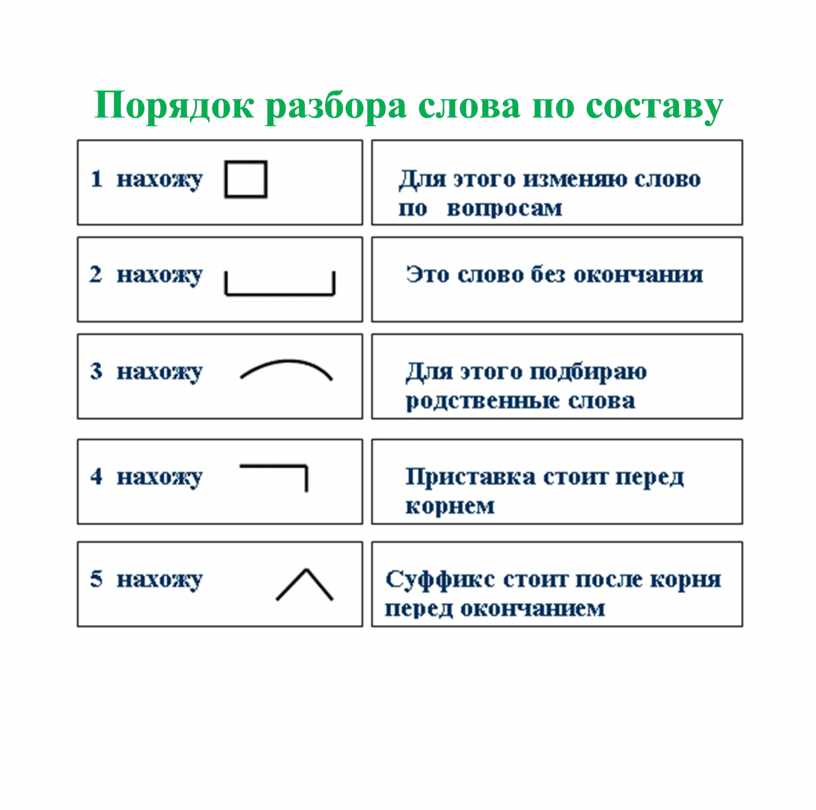
Разбор слова «мягкий» по составу
мягкий
Части слова «мягкий»: мягк/ий
Состав слова:
мягк — корень,
ий — окончание,
мягк — основа слова.
Онлайн тест по Русскому языку по теме Разбор наречия по составу
Тест состоит из семи заданий с выбором ответа, предполагающих не только теоретические знания, но также наличие практических навыков у учащегося. Для того, чтобы ответить на первый вопрос, нужно вспомнить общее определение разложения слова по составу. Исходя из определения, это выделение его морфем, которое выполняется согласно установленной схеме. Склоняемые части речи можно изменять по падежам и формам, чтобы выделить основу и окончание, но наречие остается без изменений, а значит, чтобы выделить в нем морфемы, нужно понять, от какого слова оно образовано. Далее необходимо вспомнить, какие формообразующие суффиксы они могут иметь.
Обучающийся должен уметь правильно выделять в предложенных словах в следующем порядке: сначала формообразующий суффикс, затем основу, приставку, корень, и, наконец, обычный суффикс, если он есть. Следует помнить, что окончание у любого наречия всегда будет нулевым. При владении всеми перечисленными навыками, верно решить все задания теста не составит никакой сложности.
Пройти тест онлайн
Может быть интересно
Ещё никто не оставил комментария, вы будете первым.
Написать комментарий
Спасибо за комментарий, он будет опубликован после проверки
Фонетический разбор слова «Привлекательнейшей». Сколько звуков и букв в слове?
Транскрипция слова: [ пр’ивл’иќат’ил’н’ий’шый’ ]
В слове «Привлекательнейшей»: 18 букв, 17 звуков (6 гласных, 11 согласных).
| Буква | Звук | Характеристика звука |
|---|---|---|
| п | [ п ] | согласный, глухой парный, твёрдый парный |
| р | [ р’ ] | согласный, звонкий непарный (сонорный), мягкий парный |
| и | [ и ] | гласный, безударный |
| в | [ в ] | согласный, звонкий парный, твёрдый парный |
| л | [ л’ ] | согласный, звонкий непарный (сонорный), мягкий парный |
| е | [ и ] | гласный, безударный |
| к | [ к ] | согласный, глухой парный, твёрдый парный |
| а | [ ́а ] | гласный, ударный |
| т | [ т’ ] | согласный, глухой парный, мягкий парный |
| е | [ и ] | гласный, безударный |
| л | [ л’ ] | согласный, звонкий непарный (сонорный), мягкий парный |
| ь | ||
| н | [ н’ ] | согласный, звонкий непарный (сонорный), мягкий парный |
| е | [ и ] | гласный, безударный |
| й | [ й’ ] | согласный, звонкий непарный (сонорный), мягкий непарный |
| ш | [ ш ] | согласный, глухой парный, твёрдый непарный |
| е | [ ы ] | гласный, безударный |
| й | [ й’ ] | согласный, звонкий непарный (сонорный), мягкий непарный |
Цветовая схема: п р и в л е к а т е л ь н е й ш е й
Наноструктурный вид процесса разборки клеточной стенки во время созревания плодов и послеуборочного хранения с помощью атомно-силовой микроскопии
Основные моменты
- •
Во время размягчения плодов происходит разрушение клеточной стенки и потеря средней ламели.

- •
Наноструктурные свойства стеновых компонентов можно охарактеризовать с помощью АСМ.
- •
Утрата пектиновой сложности, длины и разветвленности коррелирует со смягчением.
- •
Послеуборочные обработки, уменьшающие размягчение, сохраняют структуру пектина.
- •
АСМ-визуализация целлюлозы и гемицеллюлозы сильно зависит от процедуры экстракции.
Реферат
Предпосылки
Механические свойства клеточных стенок паренхимы, а также прочность и протяженность областей адгезии между соседними клетками вместе с клеточным тургором являются основными определяющими факторами прочности мясистых плодов.Эти свойства изменяются во время созревания, что приводит к размягчению плодов. Модификации клеточной стенки включают деполимеризацию гликанов и пектинов матрикса, солюбилизацию пектинов и потерю нейтральных сахаров из боковых цепей пектина. Эти изменения ослабляют клеточные стенки и увеличивают разделение клеток, что в сочетании со снижением тургора клеток вызывает структурные изменения. Атомно-силовая микроскопия (АСМ) использовалась для характеристики наноструктуры полисахаридов клеточной стенки во время созревания и хранения нескольких плодов после сбора урожая.Этот метод позволяет получать изображения отдельных полимеров при большом увеличении с минимальной пробоподготовкой.
Объем и подход
В этой статье рассматриваются основные особенности процесса разборки клеточной стенки, связанные с размягчением плодов, с наноструктурной точки зрения, как это было предоставлено исследованиями АСМ.
Основные результаты и выводы
Исследования AFM показывают, что размер, разветвленность и сложность пектина уменьшаются во время созревания и хранения плодов, и в большинстве случаев эти изменения коррелируют с размягчением.Доказано, что послеуборочные обработки, улучшающие качество плодов, сохраняют структуру пектина, что указывает на четкую связь между размягчением и метаболизмом пектина. Наноструктурные характеристики целлюлозы и гемицеллюлозы во время созревания мало изучены АСМ, и имеющиеся скудные результаты не являются окончательными. В глобальном масштабе AFM может стать мощным инструментом для понимания основ текстурного качества свежих и хранящихся фруктов.
Наноструктурные характеристики целлюлозы и гемицеллюлозы во время созревания мало изучены АСМ, и имеющиеся скудные результаты не являются окончательными. В глобальном масштабе AFM может стать мощным инструментом для понимания основ текстурного качества свежих и хранящихся фруктов.
Ключевые слова
Атомно-силовая микроскопия
Клеточная стенка
Гемицеллюлоза
Смягчение плодов
Текстура фруктов
Пектины
Послеуборочные
Рекомендуемые статьиЦитирующие статьи (0)
Просмотр аннотации.Опубликовано Elsevier Ltd.
Рекомендуемые статьи
Цитирующие статьи
Frontiers | Состав полимеров клеточной стенки и пространственное распределение в спелых бананах и плодах манго: влияние на адгезию клеток и восприятие текстуры
Введение
Банан ( Musa acuminata ) и манго ( Mangifera indica ) — две важные тропические культуры, потребляемые во всем мире из-за их сенсорных и пищевых свойств. Однако их текстура на стадии созревания заметно отличается.Восприятие текстуры фруктов определяется комплексными сигналами, включая физические и химические реакции на компоненты пищи. Текстура является вторым по важности аспектом сенсорной приемлемости мясистых плодов помимо внешнего вида (Contador et al., 2015). Хотя сенсорный анализ и реологическое тестирование являются классическими подходами к определению текстурного восприятия (Colin-Henrion et al., 2007; Charles et al., 2017), в последнее время утверждается, что оральная обработка включает не только объемную реологию (например.g., вязкость), но также и трибологические явления с преобладанием поверхности (например, трение и смазывание), особенно на более поздних стадиях обработки полости рта (Chen and Stokes, 2012; Stokes et al., 2013; Sarkar et al., 2019). Недавно трибология была успешно использована для понимания орального восприятия с преобладанием поверхности с использованием эмпирических корреляций между коэффициентами трения (μ) и характеристиками вкусовых ощущений, такими как скользкость и пастообразность биополимерных гидрогелей (Krop et al.
Как банан, так и манго были описаны как имеющие «тающую текстуру», при которой ткань распадается в полости рта без жевания (Contador et al., 2015). Спелые плоды банана вызывают сложную текстурную реакцию, описываемую как мучнистая и слегка вяжущая текстура (Valente et al., 2011), которая контрастирует с мясистой, скользкой и сочной текстурой плодов манго (Suwonsichon et al., 2012). Оба типа фруктов подвергаются климактерическому созреванию с быстрыми биохимическими и биофизическими изменениями, в результате которых плоды размягчаются в течение нескольких дней после начала созревания (Ali et al., 2004). Несколько скоординированных процессов приводят к разборке клеточной стенки и средних ламелл, что приводит к потере тургора и разделению клеток (Brummell and Harpster, 2001).Разборка клеточной стенки широко изучалась на помидорах ( Solanum esculentum ) в качестве модельной системы созревания климактерических плодов (Rose and Bennett, 1999; Wang et al., 2018). Хотя банан был предложен в качестве модельной системы для созревания однодольных растений (D’Hont et al., 2012), мало что известно о том, как разрушается клеточная стенка банана. Сильное усиление (до 12 раз) генов, кодирующих пектинлиазы (PL), эндотрансгликозилазу / гидролазы ксилоглюкана (XTH) и экспансины, наблюдалось в спелых фруктах по сравнению с незрелыми фруктами, в то время как некоторые изоформы эндо-полигалактуроназы (PG), пектина Метилэстераза (PME) и целлюлаза также активировались в меньшей степени (Asif et al.
Более того, роль клеточной адгезии и специфических полимеров клеточной стенки в обработке полости рта и восприятии текстуры все еще плохо изучена. Стоит отметить, что некоторые ферменты клеточной стенки продолжают быть активными в оральной фазе, и их активность может влиять на текстуру.В томатах активность PME была обнаружена в имитируемых условиях пероральной обработки и была связана со снижением вязкости в течение 1 минуты после пероральной обработки (Rabiti et al., 2018). Кроме того, неповрежденность клеточных стенок плодов является сильным положительным фактором, определяющим вязкость фруктовых продуктов (Chu et al., 2017), и отрицательно связана с ферментационным потенциалом микробиоты (Low et al., 2015). Оба эти свойства важны для пользы для здоровья, связанной с употреблением фруктов (Dreher, 2018).
Визуализация полимеров клеточной стенки в muro с использованием зондов антител может дать представление о функции полимера (Lee et al., 2011), и этот подход предполагает потенциальную роль различных доменов пектина и ксилоглюкана в опосредовании клеточной адгезии в созревающих плодах томатов ( Орфила и др. , 2001; Ордаз-Ортис и др., 2009). Антитела также являются полезными инструментами для профилирования полисахаридных эпитопов в популяциях полисахаридов, извлеченных из клеточных стенок (Pattathil et al., 2010; Cornuault et al., 2014), хотя этот метод ранее не использовался для оценки полимеров, солюбилизированных во время разделения клеток. Атомно-силовая микроскопия использовалась для визуализации структуры фракций клеточной стенки плодов (Paniagua et al., 2014; Cárdenas-Pérez et al., 2018; Posé et al., 2018) и интактной клеточной поверхности луковых клеток (Zhang et al., 2018). др., 2016). АСМ предоставляет дополнительную структурную информацию для иммунофлуоресцентной микроскопии.
, 2001; Ордаз-Ортис и др., 2009). Антитела также являются полезными инструментами для профилирования полисахаридных эпитопов в популяциях полисахаридов, извлеченных из клеточных стенок (Pattathil et al., 2010; Cornuault et al., 2014), хотя этот метод ранее не использовался для оценки полимеров, солюбилизированных во время разделения клеток. Атомно-силовая микроскопия использовалась для визуализации структуры фракций клеточной стенки плодов (Paniagua et al., 2014; Cárdenas-Pérez et al., 2018; Posé et al., 2018) и интактной клеточной поверхности луковых клеток (Zhang et al., 2018). др., 2016). АСМ предоставляет дополнительную структурную информацию для иммунофлуоресцентной микроскопии.
Это исследование было направлено на выяснение молекулярного механизма, лежащего в основе текстурных различий между бананом и манго.Мы использовали новую комбинацию методов в различных масштабах для анализа свойств отделенных ячеек плодов и их потенциального вклада в обработку полости рта и восприятие текстуры.
Материалы и методы
Растительные материалы
плодов банана ( Musa acuminata, var Cavendish) и манго ( Mangifera indica, var Kesar) были куплены на рынке в Лидсе, Англия. Плоды манго были отнесены к пятой стадии, были мягкими и полностью созревшими без каких-либо признаков гниения (Nambi et al., 2015). Плоды банана были на седьмой стадии с желтым цветом, мягкой текстурой и коричневыми пятнами (Soltani et al., 2010). Фрукты очищали и ткань паренхимы осторожно соскребали с помощью металлического шпателя, пропускали через сито с крупными ячейками (250 мкм) и переносили в пробирку, содержащую воду MiIliQ, до конечной суспензии 9,0 мас.%. Образец супернатанта собирали для анализа гликома солюбилизированных полимеров. Два фрукта каждого вида обрабатывали как биологические копии для каждого эксперимента.Репрезентативные фотографии были выбраны для маркировки и экспериментов АСМ.
Объемная реология
Реологические характеристики суспензий клеток манго или банана (9,0 мас. % Клеток в воде MiIliQ) проводили с использованием реометра с контролируемым напряжением (Kinexus Ultra, Malvern Instruments Ltd, Вустершир, Соединенное Королевство). Температуру контролировали на уровне 37 ° C, чтобы имитировать физиологические условия. Геометрия конуса на пластине (40 мм, 4 °) использовалась для измерения поведения стационарного потока в зависимости от скорости сдвига в диапазоне от 0.От 1 до 1000 с –1 . Результаты представлены в виде средних значений и стандартных отклонений не менее трех измерений каждого образца фруктовой суспензии. Два фрукта каждого вида обрабатывали как биологические копии.
% Клеток в воде MiIliQ) проводили с использованием реометра с контролируемым напряжением (Kinexus Ultra, Malvern Instruments Ltd, Вустершир, Соединенное Королевство). Температуру контролировали на уровне 37 ° C, чтобы имитировать физиологические условия. Геометрия конуса на пластине (40 мм, 4 °) использовалась для измерения поведения стационарного потока в зависимости от скорости сдвига в диапазоне от 0.От 1 до 1000 с –1 . Результаты представлены в виде средних значений и стандартных отклонений не менее трех измерений каждого образца фруктовой суспензии. Два фрукта каждого вида обрабатывали как биологические копии.
Мягкая трибология
Измерения трения были выполнены в присутствии клеточных суспензий (9,0 мас.% Клеток манго или банана в воде MilliQ) с использованием Mini Traction Machine 2 (MTM2, PCS instruments, Лондон, Соединенное Королевство) с установкой из мягкого полимерного шарика на диске. используя небольшую модификацию ранее описанного метода (Laguna et al., 2017; Кроп и др., 2019). Трибологическая установка включала гидрофобные контактные поверхности (угол контакта с водой 108 ∘ (Sarkar et al., 2017) с участием гладкого полидиметилсилоксанового (PDMS) шарика (радиус 6,35 мм) на гладком диске PDMS (радиус 13 мм, толщина 4 мм). ) в камере мини-горшка. Для каждого отдельного измерения использовались свежий шар и диск, и все измерения трения проводились при 37 ° C для имитации условий полости рта. Нормальная нагрузка ( F n ) 2 Во всех экспериментах использовался N, а скорость уноса варьировалась от 300 до 3 мм с –1 .Скорость уноса ( U ) рассчитывалась по уравнению (1):
U = 12 (UB + UD) (1)
Где U B и U D — скорости шара и диска соответственно. Отношение скольжения к ролику, определяемое как | U B — U D | / U было зафиксировано на уровне 50%. Сила трения ( F f = μ. F n ) измеряли как функцию скорости уноса, а безразмерный коэффициент трения (μ) сообщали как средние значения и стандартные отклонения по меньшей мере трех измерений каждого образца фруктовой суспензии. Два фрукта каждого вида обрабатывали как биологические копии.
Сила трения ( F f = μ. F n ) измеряли как функцию скорости уноса, а безразмерный коэффициент трения (μ) сообщали как средние значения и стандартные отклонения по меньшей мере трех измерений каждого образца фруктовой суспензии. Два фрукта каждого вида обрабатывали как биологические копии.
Цитохимическое окрашивание клеточной поверхности
Для неспецифического окрашивания клеточной мембраны и содержимого 0,05% (мас. / Об.) Толуидинового синего O (T3260, Sigma-Aldrich) в 0,1 М фосфатном буфере, pH 6.8 добавляли к фруктовой ткани в пробирке. После окрашивания в течение 5 мин окрашенные образцы помещали на предметные стекла, покрытые поли-L-лизином (Polysine, J2800AMNZ, Thermo-Scientific). Для окрашивания крахмалом ткань плода диспергировали в дистиллированной воде и помещали на предметное стекло, покрытое полизином, затем добавляли одну каплю раствора йода по Граму (
, Sigma-Aldrich) и смешивали непосредственно на предметном стекле. Для окрашивания целлюлозы 0,1% (мас. / Об.) Краситель Calcofluor White [Fluorescent Brightener 28 (319945), Sigma-Aldrich] добавляли к фруктовой ткани в пробирке.Одну каплю окрашенного образца помещали на предметное стекло, покрытое полизином, затем подщелачивали одной каплей 10% (об. / Об.) NaOH. Образец исследовали с использованием инвертированного светового микроскопа для окрашивания толуидиновым синим O и йодом и УФ-флуоресцентного микроскопа для окрашивания Calcofluor White (Olympus, модель Bh3, Япония). Изображения были получены с помощью цифровой камеры (Sony, модель sCMEX-3). Все окрашивание проводили при комнатной температуре.Маркировка иммунофлуоресценции клеточной поверхности
Ткань плода собирали, как описано выше.Поверхность плодовых клеток иммуно метили крысиными моноклональными антителами к эпитопам полисахаридов стенок растительных клеток. Для этого эксперимента было выбрано семь антител: LM28 (Cornuault et al., 2015), LM25 (Pedersen et al. , 2012), LM21 (Marcus et al., 2010), JIM5 и JIM7 (Clausen et al., 2003), LM5 (Jones et al., 1997), LM6-M (Cornuault et al., 2017). Список антител и эпитопов доступен на http://www.plants.leeds.ac.uk/pk/pdf/JPKab05.pdf. Супернатанты гибридомы с антителами перед использованием разводили 10 раз в 3% (мас. / Об.) Обезжиренном сухом молоке (Marvel) в 10 мМ фосфатно-солевом буфере (PBS).Во-первых, препараты силана (Thermo-fisher) активировали с использованием 2,5% (об. / Об.) Глутарового альдегида (A17876, Sigma-Aldrich, Сент-Луис, Миссури, США) в PBS pH 7,45. Взвешенные фруктовые клетки (50 мкл) добавляли на активированное предметное стекло для подготовки силана с последующей быстрой сушкой в течение 10 минут на горячей пластине. Поверхностные неспецифические эпитопы блокировали 50 мкл 3% (мас. / Об.) Обезжиренного сухого молока в 10 мМ PBS в течение 30 мин. Затем клетки плодов метили отобранными моноклональными антителами в течение 1 часа. После трехкратной промывки PBS по 5 мин каждая плодовые клетки инкубировали со 100-кратным разведением антикрысиного IgG-FITC (F1763, Sigma-Aldrich, St.Луис, Миссури, США) в 3% (мас. / Об.) Обезжиренном сухом молоке в 10 мМ PBS в течение 1 ч с последующими тремя 5-минутными промывками PBS. Реагент против тушения Citifluor AF1 (AGR1320, Agar Scientific) был добавлен на предметное стекло перед исследованием под флуоресцентным микроскопом (Olympus, модель Bh3), снабженным синей эпифлуоресценцией. Что касается отрицательного контроля, образец обрабатывали в соответствии с описанными выше этапами, исключая первичные моноклональные антитела. Все этапы маркировки проводились при комнатной температуре.
, 2012), LM21 (Marcus et al., 2010), JIM5 и JIM7 (Clausen et al., 2003), LM5 (Jones et al., 1997), LM6-M (Cornuault et al., 2017). Список антител и эпитопов доступен на http://www.plants.leeds.ac.uk/pk/pdf/JPKab05.pdf. Супернатанты гибридомы с антителами перед использованием разводили 10 раз в 3% (мас. / Об.) Обезжиренном сухом молоке (Marvel) в 10 мМ фосфатно-солевом буфере (PBS).Во-первых, препараты силана (Thermo-fisher) активировали с использованием 2,5% (об. / Об.) Глутарового альдегида (A17876, Sigma-Aldrich, Сент-Луис, Миссури, США) в PBS pH 7,45. Взвешенные фруктовые клетки (50 мкл) добавляли на активированное предметное стекло для подготовки силана с последующей быстрой сушкой в течение 10 минут на горячей пластине. Поверхностные неспецифические эпитопы блокировали 50 мкл 3% (мас. / Об.) Обезжиренного сухого молока в 10 мМ PBS в течение 30 мин. Затем клетки плодов метили отобранными моноклональными антителами в течение 1 часа. После трехкратной промывки PBS по 5 мин каждая плодовые клетки инкубировали со 100-кратным разведением антикрысиного IgG-FITC (F1763, Sigma-Aldrich, St.Луис, Миссури, США) в 3% (мас. / Об.) Обезжиренном сухом молоке в 10 мМ PBS в течение 1 ч с последующими тремя 5-минутными промывками PBS. Реагент против тушения Citifluor AF1 (AGR1320, Agar Scientific) был добавлен на предметное стекло перед исследованием под флуоресцентным микроскопом (Olympus, модель Bh3), снабженным синей эпифлуоресценцией. Что касается отрицательного контроля, образец обрабатывали в соответствии с описанными выше этапами, исключая первичные моноклональные антитела. Все этапы маркировки проводились при комнатной температуре.
Атомно-силовая микроскопия клеточной поверхности (АСМ)
Ткань плода собирали, как описано выше.Суспензии клеток далее пропускали через металлическое сито со средним размером ячеек (150 мкм) для удаления рыхлого крахмала, при этом ретентат промывали водой MilliQ (3 × 50 мл) и ресуспендировали в воде MilliQ. 200 мкл клеточной суспензии наносили на покровное стекло и давали ему высохнуть в течение не менее 48 часов (при комнатной температуре) перед формированием изображений с помощью АСМ. Высушенные образцы получали с помощью Multimode ® AFM со сканером J (Bruker, Калифорния, США) с PF QNM (количественное картирование наномеханических свойств PeakForce).Изображения были сглажены, чтобы удалить изгиб в каждой строке сканирования, и экспортированы в формат TIFF. Для каждого образца сканировали не менее пяти различных клеток при 0,8–0,9 Гц. Для визуализации были выбраны только целые отдельные клетки (т.е. клетки, которые не были прикреплены к другим клеткам), что сводит к минимуму вероятность того, что внутренняя поверхность будет отображена. Были выбраны пять областей на каждой ячейке в областях, которые не пересекали очевидную складку или морщину, вызванную высыханием круглых клеток на плоской поверхности. Затем для бумаги были выбраны репрезентативные изображения.
Высушенные образцы получали с помощью Multimode ® AFM со сканером J (Bruker, Калифорния, США) с PF QNM (количественное картирование наномеханических свойств PeakForce).Изображения были сглажены, чтобы удалить изгиб в каждой строке сканирования, и экспортированы в формат TIFF. Для каждого образца сканировали не менее пяти различных клеток при 0,8–0,9 Гц. Для визуализации были выбраны только целые отдельные клетки (т.е. клетки, которые не были прикреплены к другим клеткам), что сводит к минимуму вероятность того, что внутренняя поверхность будет отображена. Были выбраны пять областей на каждой ячейке в областях, которые не пересекали очевидную складку или морщину, вызванную высыханием круглых клеток на плоской поверхности. Затем для бумаги были выбраны репрезентативные изображения.
Приготовление нерастворимого в спирте остатка (AIR)
Готовили нерастворимый в спирте остаток из каждого плода. Ткань плода (3 г ) гомогенизировали при 13000 г (Polytron, модель 2500 E, Швейцария) с 7 г 100% этанола в течение примерно 1 мин до получения гомогенного образца, что дало конечную концентрацию этанола 70%. . Затем образец центрифугировали при 3500 g в течение 20 мин (центрифуга Heraeus Megafuge 16R, Германия) при комнатной температуре.Супернатант удаляли, а остаток ресуспендировали в 70% (об. / Об.) Этаноле, гомогенизировали при 13000 g в течение 30 с и центрифугировали при 5000 об / мин в течение 20 минут. Остаток многократно промывали серией растворителей: 80% (об. / Об.) Этанол, 90% (об. / Об.) Этанол, 100% (об. / Об.) Этанол, 100% (об. / Об.) Ацетон и метанол: хлороформ. (2: 3). Эти шаги были направлены на осаждение растворимых волокон, удаление компонентов с малой молекулярной массой и инактивацию ферментов. Полученный AIR сушили в течение ночи в вытяжном шкафу перед экстракцией для определения профиля иммунного гликома.
Гликометрический профиль клеточной стенки
Гликом-анализ — это метод на основе ELISA, который позволяет быстро анализировать полисахаридные эпитопы, обнаруженные во фракциях солюбилизированных клеточных стенок (Pattathil et al. , 2010). AIR последовательно экстрагировали 50 мМ CDTA, 4 М КОН и 1 мкг / мл целлюлазы 5a (NZYTech). AIR (4 мг) помещали в пробирки на 2 мл и в образец добавляли шариковые подшипники перед измельчением в Tissue Lyser при 50 Гц в течение 2 мин. Затем добавляли 50 мМ CDTA и измельчали в течение 20 минут в анализаторе тканей с последующим покачиванием пробирки в течение 40 минут и центрифугированием при 3500 g в течение 15 минут.Супернатант сохраняли как фракцию CDTA, а остатки затем подвергали воздействию следующего экстрагирующего реагента. Остатки экстрагировали 4 М КОН с 1% NaBH 4, , получая фракцию КОН. Затем остатки обрабатывали 1 мкг / мл целлюлазы в 20 мМ Трис-буфере pH 8,8 и инкубировали в течение 2 часов при 37 ° C перед центрифугированием при 14000 об / мин в течение 15 минут. Супернатант сохраняли как фракцию целлюлазы. Фракции экстрагированных клеточных стенок или супернатанты от образцов, разделенных клетками, разбавляли в 10 раз перед нанесением покрытия на планшеты с иммуносорбентом (Nunc) в течение ночи при 4 ° C.Затем планшеты промывали водопроводной водой 9 раз и блокировали 5% (мас. / Об.) Обезжиренным сухим молоком в 10 мМ PBS (M / PBS) в течение 2 часов. После промывания водопроводной водой еще девять раз добавляли разведение 1:10 моноклональных антител в M / PBS (только разведение 1: 300 для антител к каллозе) и инкубировали в течение 1,5 часов. Каждая лунка планшета содержала один тип антител, и каждое антитело было взято в дубликатах лунок. В анализе использовали 40 антител. Большинство из них были моноклональными антителами крысы, за исключением антикаллозы, которая была повышена у мышей (BioSupplies, Австралия).После инкубации с первичными антителами лунки промывали девять раз водопроводной водой, затем разбавляли 1: 1000 вторичных антител в M / PBS (антитела против мышиного IgG-HRP для антител к каллозе и анти-крысиные IgG-HRP для всех остальных. , оба получены от Invitrogen) наносили на 1 час. Планшеты промывали девять раз водопроводной водой с последующим добавлением субстрата для генерации сигнала.
, 2010). AIR последовательно экстрагировали 50 мМ CDTA, 4 М КОН и 1 мкг / мл целлюлазы 5a (NZYTech). AIR (4 мг) помещали в пробирки на 2 мл и в образец добавляли шариковые подшипники перед измельчением в Tissue Lyser при 50 Гц в течение 2 мин. Затем добавляли 50 мМ CDTA и измельчали в течение 20 минут в анализаторе тканей с последующим покачиванием пробирки в течение 40 минут и центрифугированием при 3500 g в течение 15 минут.Супернатант сохраняли как фракцию CDTA, а остатки затем подвергали воздействию следующего экстрагирующего реагента. Остатки экстрагировали 4 М КОН с 1% NaBH 4, , получая фракцию КОН. Затем остатки обрабатывали 1 мкг / мл целлюлазы в 20 мМ Трис-буфере pH 8,8 и инкубировали в течение 2 часов при 37 ° C перед центрифугированием при 14000 об / мин в течение 15 минут. Супернатант сохраняли как фракцию целлюлазы. Фракции экстрагированных клеточных стенок или супернатанты от образцов, разделенных клетками, разбавляли в 10 раз перед нанесением покрытия на планшеты с иммуносорбентом (Nunc) в течение ночи при 4 ° C.Затем планшеты промывали водопроводной водой 9 раз и блокировали 5% (мас. / Об.) Обезжиренным сухим молоком в 10 мМ PBS (M / PBS) в течение 2 часов. После промывания водопроводной водой еще девять раз добавляли разведение 1:10 моноклональных антител в M / PBS (только разведение 1: 300 для антител к каллозе) и инкубировали в течение 1,5 часов. Каждая лунка планшета содержала один тип антител, и каждое антитело было взято в дубликатах лунок. В анализе использовали 40 антител. Большинство из них были моноклональными антителами крысы, за исключением антикаллозы, которая была повышена у мышей (BioSupplies, Австралия).После инкубации с первичными антителами лунки промывали девять раз водопроводной водой, затем разбавляли 1: 1000 вторичных антител в M / PBS (антитела против мышиного IgG-HRP для антител к каллозе и анти-крысиные IgG-HRP для всех остальных. , оба получены от Invitrogen) наносили на 1 час. Планшеты промывали девять раз водопроводной водой с последующим добавлением субстрата для генерации сигнала.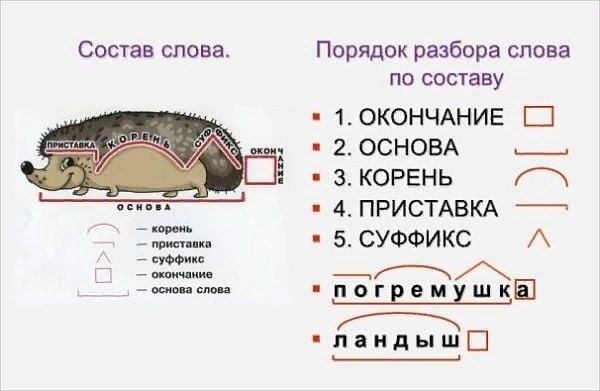 Субстрат содержал 1 М натрий-ацетатный буфер pH 6,0, тетраметилбензидин, 6% (об. / Об.) Перекись водорода и дистиллированную воду в соотношении 100: 10: 1: 1000.Реакцию останавливали добавлением 2,5 М серной кислоты, дающей желтый цвет. Сила связывания каждого антитела определялась по поглощению при 450 нм с помощью планшет-ридера для ELISA (считыватели для микропланшетов Multiskan Fc, Финляндия). Два плода каждого вида обрабатывали как биологические копии, и каждый экстракт или супернатант анализировали в лунках для повторений.
Субстрат содержал 1 М натрий-ацетатный буфер pH 6,0, тетраметилбензидин, 6% (об. / Об.) Перекись водорода и дистиллированную воду в соотношении 100: 10: 1: 1000.Реакцию останавливали добавлением 2,5 М серной кислоты, дающей желтый цвет. Сила связывания каждого антитела определялась по поглощению при 450 нм с помощью планшет-ридера для ELISA (считыватели для микропланшетов Multiskan Fc, Финляндия). Два плода каждого вида обрабатывали как биологические копии, и каждый экстракт или супернатант анализировали в лунках для повторений.
Анализ данных
Для окрашивания клеток и иммунофлуоресцентного мечения (качественный анализ) было выбрано одно микроскопическое изображение как репрезентативное из пяти захваченных изображений.Для профилирования гликома клеточной стенки стандартное отклонение было рассчитано с использованием Microsoft Excel из двух повторных экспериментов, и коэффициент вариации <15% был установлен в качестве приемлемого предела.
Результаты
Разделение клеток и окрашивание клеточной поверхности клеток банана и манго
Ткань паренхимы обоих плодов была спелой, мягкой, и клетки легко отделялись при небольшом стрессе. Окрашивание тканей выявило некоторые заметные различия в морфологии изолированных клеток (рис. 1).Ткань банана показала удлиненные, в основном неповрежденные клетки, которые оставались прикрепленными своими апикальными кончиками, очертания клеток четко визуализировались при окраске толуидиновым синим (рис. 1а). Они содержали несколько гранул крахмала, сильно окрашенных йодом (рис. 1b). Неповрежденность клеточной стенки была подтверждена окрашиванием Calcofluor White (рис. 1c), которое также выявило наличие небольших отверстий, напоминающих поля ямок (обозначенных желтой стрелкой на рис. 1c), организованных на узкой полосе по длине клетки.Этот образец предполагает, что клетки когда-то прикреплялись вдоль этой полоски, но адгезия была легко нарушена незначительным стрессом (например, осторожным соскабливанием шпателем). Напротив, клетки манго были более округлой формы (рис. 1d) и содержали несколько гранул крахмала (рис. 1e). Окрашивание толуидиновым синим не позволило выделить клетки так четко, как в случае банана. Окрашивание выявило овальные структуры на поверхности клеток. Нам не ясно, что это такое, но это могут быть очертания больших участков, содержащих поля ям. Окрашивание Calcofluor White показало большие участки клеточной стенки, которые, по-видимому, были разорваны (обозначены * на рисунке 1f), а также ярко окрашенные овальные области, которые содержали обильные поля ямок (обозначены стрелкой на рисунке 1f).Локализация ямок как у банана, так и у манго предполагает, что они могут способствовать адгезии клеток в этих фруктах.
Напротив, клетки манго были более округлой формы (рис. 1d) и содержали несколько гранул крахмала (рис. 1e). Окрашивание толуидиновым синим не позволило выделить клетки так четко, как в случае банана. Окрашивание выявило овальные структуры на поверхности клеток. Нам не ясно, что это такое, но это могут быть очертания больших участков, содержащих поля ям. Окрашивание Calcofluor White показало большие участки клеточной стенки, которые, по-видимому, были разорваны (обозначены * на рисунке 1f), а также ярко окрашенные овальные области, которые содержали обильные поля ямок (обозначены стрелкой на рисунке 1f).Локализация ямок как у банана, так и у манго предполагает, что они могут способствовать адгезии клеток в этих фруктах.
Рис. 1. Микрофотографии клеток банана, окрашенных толуидиновым синим (a) , йодом (b) и калькофлуором белым (c) ; и клетки манго, окрашенные толуидиновым синим (d) , йодом (e) и калькофлуором белым (f) . Масштабная линейка = 100 мкм. Красные стрелки указывают на гранулы крахмала, четко видимые в банане, желтые стрелки указывают на расположение ямок в полосах на банане и круглых косточках в манго. * Указывает на разрыв клеточной стенки плода манго.
Для более подробного изучения распределения полимеров клеточной стенки на поверхности клеток ткань плода метили семью моноклональными антителами, которые распознают различные эпитопы пектина и гемицеллюлозы. Как показано на фиг. 2, в клеточных стенках бананов обнаружено сильное и равномерное распределение эпитопов гемицеллюлозы, меченных антителами LM28 (анти-ксилан) и LM25 (анти-ксилоглюкан). LM21 (антиманнан) и JIM7 (антиметилэтерифицированный HG) показали точечное мечение по всей клеточной стенке.Яркая флуоресценция была обнаружена с помощью маркировки JIM5 (анти-гомогалактуронан), с самой яркой маркировкой на вершине клеток, где наблюдалась клеточная адгезия. Мечение доменов рамногалактуронана-I (RG-I) с помощью LM5 (антигалактан) и LM6 (антилинейный арабинан) было менее интенсивным, хотя полосатый рисунок можно было различить с помощью мечения LM5. Маркировка ткани манго показала иную схему маркировки. Наиболее сильное мечение наблюдалось с LM25 (антиксилоглюкан), за которым следовали LM5 (антигалактан) и LM8 (антиксилан).Не наблюдалось точечного мечения JIM7 (антиметилэтерифицированный HG) или LM21 (антиманнан). Мечение антителом JIM5 было слабым, но более сильное окрашивание наблюдалось на овальных участках, напоминающих поля ямок. Подобно банану, маркировка LM5 и LM6 не была интенсивной. Таким образом, схемы маркировки предполагают изменение поверхностных свойств клеток манго и банана.
Маркировка ткани манго показала иную схему маркировки. Наиболее сильное мечение наблюдалось с LM25 (антиксилоглюкан), за которым следовали LM5 (антигалактан) и LM8 (антиксилан).Не наблюдалось точечного мечения JIM7 (антиметилэтерифицированный HG) или LM21 (антиманнан). Мечение антителом JIM5 было слабым, но более сильное окрашивание наблюдалось на овальных участках, напоминающих поля ямок. Подобно банану, маркировка LM5 и LM6 не была интенсивной. Таким образом, схемы маркировки предполагают изменение поверхностных свойств клеток манго и банана.
Рис. 2. Банан (a – h) и манго (i – p) Клетки , меченные LM28, LM25, LM21, JIM5, JIM7, LM5 и LM6-M антителами, наблюдаемые под флуоресцентным микроскопом, оснащенным синей эпифлуоресценцией .Масштабная линейка = 50 мкм. Стрелки указывают на маркировку на концах банановых ячеек.
Атомно-силовая микроскопия
Поверхностные свойства клеток банана и манго, разделенных сдвигом, оценивали с помощью АСМ. Сканировались только клетки, которые четко разделены (а не разорваны), чтобы избежать наблюдения за внутренними поверхностями. На рис. 3 представлены репрезентативные изображения клеточных поверхностей с заметными различиями в свойствах поверхности (высоте), при этом банановые клетки демонстрируют аморфную структуру с агрегатами на поверхности, которые маскируют волокнистые структуры.Такая текстура объясняется остатками средних ламелл, которые не растворялись во время разделения клеток.
Рис. 3. Изображения высоты АСМ банана (a, b) и манго (c, d) клеток при размерах сканирования 1 мкм (слева) и 2 мкм (справа). Крупные агрегаты на поверхности клеток банана обозначены белыми стрелками. Напротив, фибриллярные структуры, приписываемые целлюлозе / гемицеллюлозе, четко видны в клеточной стенке манго.
С другой стороны, поверхность клеток манго выглядела более чистой, показывая четкую сеть микрофибрилл, встроенных в более темные области матрицы. Этот вид предполагает, что у манго произошло более глубокое растворение средней ламели.
Этот вид предполагает, что у манго произошло более глубокое растворение средней ламели.
Гликометрический анализ супернатантов разделения клеток и экстрактов клеточных стенок
Мы провели анализ супернатанта, собранного из разделенных клеток, а также полимеров, экстрагированных из AIR (Таблица 1).
Таблица 1. Профиль гликома клеточной стенки супернатантов разделения клеток и фракций, экстрагированных из банана и манго AIR, представленных на тепловой карте.
Если посмотреть на эпитопы, солюбилизированные во время разделения клеток, профили гликома оказались похожими в образцах банана и манго. В обоих случаях эпитопы пектина, обнаруженные с помощью LM18, LM19, LM20, JIM5 и JIM7, имели наивысшее относительное содержание, что указывает на солюбилизацию как метилированного, так и неметилированного HG в супернатанте разделения клеток. Пектин арабинан, но не галактан, также был обнаружен в растворимой фракции обоих плодов. Замещенный эпитоп ксилоглюкана, распознаваемый LM25 (ксилоглюкан с мотивом XLLG, XXLG и XXXG, где L и G показывают разные замены в основной цепи ксилоглюкана), также был обнаружен в супернатантах обоих плодов.Ключевым различием между двумя фруктами было присутствие маннана (распознаваемого антителом LM21) и ферулированного ксилана (распознаваемого антителом LM12) в супернатанте для отделения клеток банана, но не в манго. Этот анализ подтверждает присутствие маннана на поверхности клеток банана, некоторые из которых растворяются во время разделения клеток. Следует отметить, что не были предприняты шаги для инактивации ферментов во время экспериментов по разделению клеток, поскольку большинство процедур, используемых для инактивации ферментов, вероятно, повлияют на разделение клеток и солюбилизацию полимера.Роль эндогенных ферментов в восприятии текстуры требует дальнейшего изучения. Недавно наблюдалась активность PME во время пероральной обработки томатов (Rabiti et al., 2018).
Последовательная экстракция полимеров клеточной стенки CDTA, KOH и целлюлазы из AIR. В целом уровень растворимых эпитопов в манго был выше, чем в банане. В частности, CDTA солюбилизировал больше эпитопов HG и ксилоглюкана из AIR манго по сравнению с бананом. Маннан был солюбилизирован из обоих плодов с помощью CDTA, что позволяет предположить, что он легко экстрагируется.Эпитоп LM5 был очень распространен во всех фракциях манго, но обнаружил лишь незначительные уровни, обнаруженные в банане. Эпитопы разветвленных галактанов, обнаруженные LM26, были обнаружены на низких уровнях во всех фракциях манго, но не в банане. Относительное содержание AGP и экстенсинов было выше в манго по сравнению с бананом для большинства используемых антител. Гликом-анализ позволяет быстро анализировать полисахаридные эпитопы, обнаруженные во фракциях солюбилизированных клеточных стенок (Pattathil et al., 2010). Однако он не позволяет количественно определять полимеры.
В целом уровень растворимых эпитопов в манго был выше, чем в банане. В частности, CDTA солюбилизировал больше эпитопов HG и ксилоглюкана из AIR манго по сравнению с бананом. Маннан был солюбилизирован из обоих плодов с помощью CDTA, что позволяет предположить, что он легко экстрагируется.Эпитоп LM5 был очень распространен во всех фракциях манго, но обнаружил лишь незначительные уровни, обнаруженные в банане. Эпитопы разветвленных галактанов, обнаруженные LM26, были обнаружены на низких уровнях во всех фракциях манго, но не в банане. Относительное содержание AGP и экстенсинов было выше в манго по сравнению с бананом для большинства используемых антител. Гликом-анализ позволяет быстро анализировать полисахаридные эпитопы, обнаруженные во фракциях солюбилизированных клеточных стенок (Pattathil et al., 2010). Однако он не позволяет количественно определять полимеры.
Объемная реология
На рис. 4А показано, что водные суспензии клеток манго и банана демонстрируют явное истончение при сдвиге с кажущейся вязкостью, демонстрирующей снижение на три порядка величины как функцию скорости сдвига в пределах экспериментального окна. Наблюдаемое поведение истончения при сдвиге этих клеточных суспензий можно отнести к вызванному сдвиговым потоком разрушению тех агрегатов клеток банана или манго на отдельные клетки, которые выстраивались в направлении потока, как показано на схеме (рис. 4С).
Рис. 4. Кажущаяся вязкость как функция скорости сдвига (A) и коэффициенты трения как функция скорости уноса (B) для суспензий клеток манго и банана с соответствующими схемами, отображенными для реологии (C) и трибологические явления (BL, режим граничной смазки; ML, режим смешанной смазки) (D) . Вода MilliQ использовалась в качестве контроля как в реологических, так и в трибологических экспериментах. Планки погрешностей представляют собой стандартное отклонение как минимум трех измерений.
Примечательно, что суспензия клеток банана показала определенную предельную вязкость при нулевой скорости сдвига при 10 Па · с и второе плато Ньютона при 3 × 10 –3 Па · с. С другой стороны, суспензия клеток манго проявляла экстремальное истончение при сдвиге, причем значения плато не наблюдались до тех пор, пока скорость сдвига не составила 100 с -1 . Более важным здесь является тот факт, что обе системы показали очень похожие вязкости (0,05 Па · с) ( p > 0,05) при орально значимой скорости сдвига 50 с –1 (Ong et al., 2018), а также близкие конечные вязкости при 100 с и выше –1 . Модуль Юнга растительных клеток, измеренный с помощью зонда AFM, может варьироваться от 100 кПа до 1 МПа (Radotić et al., 2012; Zdunek and Kurenda, 2013). Даже при самых высоких скоростях сдвига (1000 с –1 ), используемых в этом исследовании, напряжение сдвига на клетки, создаваемое жидкостью-носителем, не может превышать 10 Па. Следовательно, можно предположить, что обе системы сохранят неповрежденные клетки после сдвиг, как схематично показано на рисунке 4C, обеспечивая структурные аспекты с более высоким сопротивлением потоку по сравнению с водой.
С другой стороны, суспензия клеток манго проявляла экстремальное истончение при сдвиге, причем значения плато не наблюдались до тех пор, пока скорость сдвига не составила 100 с -1 . Более важным здесь является тот факт, что обе системы показали очень похожие вязкости (0,05 Па · с) ( p > 0,05) при орально значимой скорости сдвига 50 с –1 (Ong et al., 2018), а также близкие конечные вязкости при 100 с и выше –1 . Модуль Юнга растительных клеток, измеренный с помощью зонда AFM, может варьироваться от 100 кПа до 1 МПа (Radotić et al., 2012; Zdunek and Kurenda, 2013). Даже при самых высоких скоростях сдвига (1000 с –1 ), используемых в этом исследовании, напряжение сдвига на клетки, создаваемое жидкостью-носителем, не может превышать 10 Па. Следовательно, можно предположить, что обе системы сохранят неповрежденные клетки после сдвиг, как схематично показано на рисунке 4C, обеспечивая структурные аспекты с более высоким сопротивлением потоку по сравнению с водой.
Мягкая трибология
Смазывающие характеристики суспензий клеток манго и банана показаны на Рисунке 4B, где коэффициент трения (μ) отложен в зависимости от скорости уноса. График μ в зависимости от скорости уноса для воды Milli-Q также показан для сравнения. Граничный режим смазки обычно наблюдается при самых низких скоростях уноса (≥10 мм с — 1) и характеризуется относительно высокими значениями μ, которые не зависят от скорости (сухое трение).На рисунке 4B граничный режим четко наблюдается как для суспензий клеток манго, так и для банановых клеток. Независимо от типа плода, обе клеточные суспензии показали одинаковые значения μ ( p > 0,05) в режиме граничной смазки, которые были значительно ниже, чем у воды (рис. 4В). Это указывает на смазывающие свойства.
Учитывая размер клеток (100–150 мкм в диаметре), маловероятно, что любой из типов клеток попадет в зону контакта. Даже если бы они вошли в зону контакта, они бы расплющились (Саркар и др. , 2017; Torres et al., 2018) или разорвался из-за высокого давления внутри замкнутого пространства. Следовательно, такое уменьшение значений μ в граничном режиме предполагает, что это произошло не из-за уноса неповрежденных клеток (если таковые остались), а из-за растворимых полимеров в непрерывной фазе.
, 2017; Torres et al., 2018) или разорвался из-за высокого давления внутри замкнутого пространства. Следовательно, такое уменьшение значений μ в граничном режиме предполагает, что это произошло не из-за уноса неповрежденных клеток (если таковые остались), а из-за растворимых полимеров в непрерывной фазе.
Эти растворимые полимеры достоверно адсорбировались на поверхности и образовывали пленки толщиной в несколько молекул (схематично показано на рисунке 4D) и уменьшали μ по сравнению с водой ( p <0.05). Это замечательное поведение граничной смазки отличается от поведения призрачных гранул крахмала, наблюдавшихся в предыдущем отчете (Zhang et al., 2017), где их граничные смазочные профили были близки к воде из-за присутствия неадсорбирующихся полимеров крахмала в непрерывной фазе.
При увеличении скорости увлечения (≥10 мм с –1 ) кривые показывают смешанный режим смазки с уменьшением значений μ. Уменьшение μ в этом режиме связано с частичным разделением контактных поверхностей прерывистым слоем смазки (Sarkar et al., 2019), где давление испытывает как смазка, так и поверхности. Как видно на рисунке 4B, он находится в смешанном режиме, где тип ячеек показал отличительные смазочные свойства. В частности, клетки манго с почти сферическим внешним видом (размером около 150 мкм) показали гораздо более быстрое начало смешанного режима смазки (≥10 мм с –1 ) с резким снижением μ (μ <0,05) орально значимых скоростей (50 мм с –1 ). В случае банановых клеток эллипсоидальной формы граничный режим был расширен до 100 мм с –1 (Рисунок 4B), что предполагает ограниченную вероятность того, что банановые клетки вступали в контакт с орально релевантными скоростями.В этом случае углеводные полимеры, солюбилизированные во время разделения клеток, могут влиять на реологические и трибологические свойства суспензий клеток.
Обсуждение
Предполагается, что разделение клеток из-за солюбилизации полимеров средней ламеллы, а также разборка первичной клеточной стенки способствуют текстурному восприятию спелых плодов. Результаты этого исследования показывают, что стенки клеток банана разбираются иначе, чем стенки клеток манго, во время размягчения, связанного со созреванием.Клетки банана очень легко отделяются при стрессе, но остаются неповрежденными, что свидетельствует о слабой средней ламелле, но более сильных первичных стенках. Согласно данным АСМ, клетки банана, по-видимому, удерживают агрегированный материал на поверхности, который, как здесь предполагается, является остатками средней ламеллы. Эти агрегированные структуры напоминают структуры, наблюдаемые с помощью АСМ извлеченных пектинов из незрелой клубники (Paniagua et al., 2014), но это первый раз, когда они наблюдаются непосредственно в muro . Иммунофлуоресцентная микроскопия показала, что эти агрегаты представляют собой метилэтерифицированный HG или маннан, что проявляется в виде точечных меток на поверхности клеток.Галактан также, по-видимому, имеет отчетливый паттерн мечения на поверхности, который предполагает агрегацию на поверхности клетки. Кроме того, профилирование гликома подтвердило присутствие пектинов и маннанов в супернатанте отделенных банановых клеток. Было показано, что маннаны являются основными компонентами стенок банановых клеток с относительно хорошей растворимостью (Shiga et al., 2017). Изолированные маннаны образуют слабые гели, которые легко ломаются и деформируются при деформации (Ben-Zion and Nussinovitch, 1997). Это свойство может быть очень полезным для банана для сохранения слабой адгезии между клетками, которая легко разрушается с помощью механической силы без необходимости ферментативного разрушения.Неясно, связано ли это поведение разделения клеток каким-либо образом с рассредоточением семян или оно было выбрано путем селекции людей. Было высказано предположение, что маннаны и другие гемицеллюлозы играют роль в адгезии клеток при созревании плодов томатов (Ordaz-Ortiz et al., 2009). Присутствие ферулированного ксилана в надосадочной жидкости для разделения клеток является неожиданным, поскольку они обычно извлекаются из нерастворимых фракций клеточной стенки (Schendel et al.
Результаты этого исследования показывают, что стенки клеток банана разбираются иначе, чем стенки клеток манго, во время размягчения, связанного со созреванием.Клетки банана очень легко отделяются при стрессе, но остаются неповрежденными, что свидетельствует о слабой средней ламелле, но более сильных первичных стенках. Согласно данным АСМ, клетки банана, по-видимому, удерживают агрегированный материал на поверхности, который, как здесь предполагается, является остатками средней ламеллы. Эти агрегированные структуры напоминают структуры, наблюдаемые с помощью АСМ извлеченных пектинов из незрелой клубники (Paniagua et al., 2014), но это первый раз, когда они наблюдаются непосредственно в muro . Иммунофлуоресцентная микроскопия показала, что эти агрегаты представляют собой метилэтерифицированный HG или маннан, что проявляется в виде точечных меток на поверхности клеток.Галактан также, по-видимому, имеет отчетливый паттерн мечения на поверхности, который предполагает агрегацию на поверхности клетки. Кроме того, профилирование гликома подтвердило присутствие пектинов и маннанов в супернатанте отделенных банановых клеток. Было показано, что маннаны являются основными компонентами стенок банановых клеток с относительно хорошей растворимостью (Shiga et al., 2017). Изолированные маннаны образуют слабые гели, которые легко ломаются и деформируются при деформации (Ben-Zion and Nussinovitch, 1997). Это свойство может быть очень полезным для банана для сохранения слабой адгезии между клетками, которая легко разрушается с помощью механической силы без необходимости ферментативного разрушения.Неясно, связано ли это поведение разделения клеток каким-либо образом с рассредоточением семян или оно было выбрано путем селекции людей. Было высказано предположение, что маннаны и другие гемицеллюлозы играют роль в адгезии клеток при созревании плодов томатов (Ordaz-Ortiz et al., 2009). Присутствие ферулированного ксилана в надосадочной жидкости для разделения клеток является неожиданным, поскольку они обычно извлекаются из нерастворимых фракций клеточной стенки (Schendel et al. , 2016; Ruthes et al., 2017) и локализуются в перикарпийном и алейроновом слоях твердеющих клеточных стенок в развитие зерна кукурузы (Chateigner-Boutin et al., 2016). Их присутствие не было обнаружено в банановых фруктах, и их роль требует дальнейшего изучения.
, 2016; Ruthes et al., 2017) и локализуются в перикарпийном и алейроновом слоях твердеющих клеточных стенок в развитие зерна кукурузы (Chateigner-Boutin et al., 2016). Их присутствие не было обнаружено в банановых фруктах, и их роль требует дальнейшего изучения.
Целостность банановых клеток, их размер и форма (высокое соотношение сторон (длина / диаметр), т. Е. 2-4: 1) уменьшают вероятность увлечения между поверхностями ротовой полости, то есть языком и небом, что приводит к возможному восприятию терпкости. Действительно, банановые клетки были исключены из зоны контакта, как схематично показано на фиг. 4D, что привело к некоторой степени шероховатости, поскольку клетки не уменьшали трение.Агрегаты клеток манго или банана, наблюдаемые на рисунке 1, скорее всего, представляли большую эффективную объемную долю, чем составляющие их отдельные клетки, и, следовательно, генерировали повышенные значения вязкости при низких скоростях сдвига (10 -1 с -1 ) ( Genovese, 2012; Moelants et al., 2014). Клетки банана также остаются нетронутыми при жевании и пищеварении в желудочно-кишечном тракте (Low et al., 2015; Chu et al., 2017), и эта устойчивость была очевидна в экспериментах по трению, в которых клетки банана не разрушались при более высоких скоростях сдвига.Эластичность можно объяснить более высокой деформируемостью или более высокой механической прочностью. Оба могут привести к меньшему разрыву. Дальнейшие эксперименты с АСМ, которые измеряют механическую прочность клеточных стенок, необходимы для оценки свойств интактных банановых клеток. Появляются последствия для здоровья неповрежденных клеточных стенок. Было показано, что клетки банана менее восприимчивы к ферментации микробиоты по сравнению с клетками манго (Low et al., 2015). Между тем было показано, что полисахариды, солюбилизированные из мякоти банана, включая маннаны, пектины и AGP, вызывают иммуномодулирующие реакции, благоприятные для здоровья кишечника (Shiga et al., 2017). Пектины и маннаны были обнаружены в супернатантах разделения клеток, что подтверждает их легкую растворимость.
Клетки манго, с другой стороны, как разделенные, так и разорванные. Поверхности разделенных клеток, наблюдаемые с помощью АСМ, предполагали более выраженную разборку средней ламеллы и клеточных стенок в этих областях. Однако более высокая склонность к разрыву клеток манго предполагает сильную адгезию клеток в других регионах, что, вероятно, связано с полями ямок. Физические, химические и биологические изменения клеточных стенок манго во время созревания были элегантно изучены с использованием ряда методов (Cárdenas-Pérez et al., 2018). Высокая активность PME и эндо-PG на более поздних стадиях созревания привела к повышенной растворимости пектина, более коротким и менее организованным полимерам (как видно с помощью АСМ) и механически более слабым клеточным стенкам. Эти молекулярные изменения коррелировали с более мягкой текстурой в масштабе ткани (Cárdenas-Pérez et al., 2018). Эти наблюдения подтверждаются здесь, так как стенки клеток манго казались деформируемыми при низком сдвиге, что приводило к образованию слоя, который уменьшал трение в экспериментах по трибологии. Основными полимерами, солюбилизированными во время разделения клеток манго, были в основном пектины и ксилоглюканы, в то время как маннан солюбилизировался только при химической обработке.Их солюбилизация и разборка клеточной стенки в целом объясняется активностью эндогенных ферментов клеточной стенки во время созревания, включая PME, endo-PG, PL и XTH (Chourasia et al., 2006, 2008). Солюбилизированный материал, который также может способствовать более быстрому наступлению смешанного режима смазки в клетках манго, что можно интерпретировать как гладкое и скользкое ощущение во рту.
Результаты объемной реологии предполагают, что суспензии клеток манго и банана имеют схожую объемную вязкость при перорально значимых скоростях сдвига и, следовательно, могут быть экстраполированы для получения аналогичного восприятия «толщины полости рта» на начальных этапах пероральной обработки.Но существенные различия в их биосмазывающем поведении могут объяснить их различные текстурные характеристики на более поздних этапах обработки, которые включают трение между поверхностями полости рта (например, языком и небом). Например, более низкое трение между мягкими контактными поверхностями в этих трибологических экспериментах (имитация языка и нёба) в случае клеток манго связано с включением клеток манго между этими контактными поверхностями на орально значимых скоростях (Рисунок 4). Такое более низкое трение может быть отражено как «гладкое» сенсорное восприятие после оральной обработки манго, поскольку можно предположить, что язык отделен от ротового неба тонким слоем клеток манго и не трется о ротовое небо.С другой стороны, в случае банановых клеток, они не вступали в контакт (рис. 4D), что можно интерпретировать в реальной жизни, поскольку язык трется о ротовое нёбо в отсутствие каких-либо клеток, что приводит к повышенному трению, что может быть отражено как «грубое» или «терпкое восприятие». Комбинация реологии и трибологии с анализом клеточной стенки, использованная в этом исследовании, впервые предлагает уникальный подход для получения механистического понимания вклада клеток и полимеров клеточной стенки в восприятие текстуры спелых фруктов.Кроме того, такие знания также могут быть использованы для количественного понимания механизмов, лежащих в основе сенсорного ощущения во рту во фруктах, а также в полутвердых продуктах питания, таких как фруктовые пюре и детское питание, богатое фруктами, где одной реологии массы недостаточно для механистического объяснения поверхностных взаимодействий. возникающие на более поздних этапах устной обработки. Будущие исследования необходимо провести с различными концентрациями клеточных суспензий, чтобы четко изучить влияние объемной доли клеток, модуля упругости клеток, роли слюны и взаимодействия слюны как с клетками, так и с полимерами клеточной стенки.Инструментальные исследования должны быть подкреплены количественным сенсорным анализом для изучения корреляции между инструментом и ощущением во рту.
Например, более низкое трение между мягкими контактными поверхностями в этих трибологических экспериментах (имитация языка и нёба) в случае клеток манго связано с включением клеток манго между этими контактными поверхностями на орально значимых скоростях (Рисунок 4). Такое более низкое трение может быть отражено как «гладкое» сенсорное восприятие после оральной обработки манго, поскольку можно предположить, что язык отделен от ротового неба тонким слоем клеток манго и не трется о ротовое небо.С другой стороны, в случае банановых клеток, они не вступали в контакт (рис. 4D), что можно интерпретировать в реальной жизни, поскольку язык трется о ротовое нёбо в отсутствие каких-либо клеток, что приводит к повышенному трению, что может быть отражено как «грубое» или «терпкое восприятие». Комбинация реологии и трибологии с анализом клеточной стенки, использованная в этом исследовании, впервые предлагает уникальный подход для получения механистического понимания вклада клеток и полимеров клеточной стенки в восприятие текстуры спелых фруктов.Кроме того, такие знания также могут быть использованы для количественного понимания механизмов, лежащих в основе сенсорного ощущения во рту во фруктах, а также в полутвердых продуктах питания, таких как фруктовые пюре и детское питание, богатое фруктами, где одной реологии массы недостаточно для механистического объяснения поверхностных взаимодействий. возникающие на более поздних этапах устной обработки. Будущие исследования необходимо провести с различными концентрациями клеточных суспензий, чтобы четко изучить влияние объемной доли клеток, модуля упругости клеток, роли слюны и взаимодействия слюны как с клетками, так и с полимерами клеточной стенки.Инструментальные исследования должны быть подкреплены количественным сенсорным анализом для изучения корреляции между инструментом и ощущением во рту.
Доступность данных
Необработанные данные, подтверждающие выводы этой рукописи, будут предоставлены авторами без излишних оговорок любому квалифицированному исследователю.
Авторские взносы
CO и YB-A разработали исследовательский проект. GR и SA провели микроскопию и эксперименты с гликомом. EA-R выполнила эксперименты с объемной реологией и трибологией.HL проводил эксперименты с АСМ под наблюдением CO и SC. GR, SA, EA-R, AS и CO проанализировали данные. Рукопись написали GR, CO и AS. YB-A и JPK критически рассмотрели и доработали рукопись.
Финансирование
HL финансировалась доктором философии EPSRC-SOFI CDT. студенчество при поддержке PepsiCo Inc. «Взгляды и мнения, выраженные в этой презентации, принадлежат автору и не обязательно отражают позицию или политику PepsiCo Inc.» SA финансировалась грантом Leverhume Trust Grant RPG-2016-136.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Сноски
Список литературы
Али, З. М., Чин, Л. Х., и Лазан, Х. (2004). Сравнительное исследование ферментов, разрушающих стенки, модификаций пектина и размягчения во время созревания отобранных тропических фруктов. Plant Sci. 167, 317–327.
Google Scholar
Асиф, М. Х., Лахвани, Д., Патак, С., Гупта, П., Баг, С. К., Нат, П. и др. (2014). Транскриптомный анализ ткани спелых и незрелых плодов банана позволяет определить основные метаболические сети, участвующие в процессе созревания плодов. BMC Plant Biol. 14:15. DOI: 10.1186 / s12870-014-0316-1
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бен-Цион, О., и Нусинович, А. (1997). Прогнозирование деформируемости при сжатии многослойных гелей и текстурированных фруктов, склеенных тремя различными методами склеивания. Food Hydrocoll. 11, 253–260.
Google Scholar
Браммелл, Д. А., и Харпстер, М. Х. (2001). Метаболизм клеточной стенки при смягчении и качестве плодов и его манипуляции у трансгенных растений. Завод Мол. Биол. 47, 311–340.
Завод Мол. Биол. 47, 311–340.
PubMed Аннотация | Google Scholar
Карденас-Перес, С., Чанона-Перес, Дж. Дж., Гуэмес-Вера, Н., Цибульска, Дж., Шиманска-Чарго, М., Чилинска, М., и др. (2018). Структурные, механические и ферментативные исследования пектина и целлюлозы при созревании манго. Carbohydr. Polym. 196, 313–321. DOI: 10.1016 / j.carbpol.2018.05.044
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чарльз М., Эндриззи И., Апреа Э., Замбанини Дж., Бетта Э. и Гаспери Ф. (2017). Динамические и статические сенсорные методы для изучения роли аромата на вкус и текстуру: мультисенсорный подход к восприятию яблока. Food Qual. Предпочитать. 62, 17–30.
Google Scholar
Шатеньер-Бутин, А.-Л., Ордас-Ортис, Дж.J., Alvarado, C., Bouchet, B., Durand, S., Verhertbruggen, Y., et al. (2016). Развитие околоплодника кукурузы: модель для изучения синтеза арабиноксилана и ферулоилирования. Перед. Plant Sci. 7: 1476. DOI: 10.3389 / fpls.2016.01476
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чен Дж. И Стокс Дж. Р. (2012). Реология и трибология: два различных режима ощущения текстуры пищи. Trends Food Sci. Technol. 25, 4–12.
Google Scholar
Чурасия, А., Разум, В. А., и Нат, П. (2006). Дифференциальная экспрессия пектатлиазы во время этилен-индуцированного размягчения манго после сбора урожая (Mangifera indica var. Dashehari) . Physiol. Растение. 128, 546–555.
Google Scholar
Чурасия, А., Сане, В. А., Сингх, Р. К., и Нат, П. (2008). Выделение и характеристика гена MiCel1 из манго: экспрессия, связанная со созреванием, и повышенная активность эндоглюканазы во время размягчения. Регул роста растений. 56, 117–127.
Google Scholar
Чу, Дж., Игбетар, Б. Д., и Орфила, К. (2017). Волокнистые клеточные структуры обнаруживаются в коммерческом фруктовом смузи и остаются нетронутыми во время имитации пищеварения. J. Nutr. Food Sci. 7: 576.
J. Nutr. Food Sci. 7: 576.
Google Scholar
Клаузен, М. Х., Уиллатс, В. Г., и Нокс, Дж. П. (2003). Синтетические метилгексагалактуронат гаптен-ингибиторы моноклональных антител против гомогалактуронана LM7, JIM5 и JIM7. Carbohydr.Res. 338, 1797–1800.
PubMed Аннотация | Google Scholar
Колин-Генрион, М., Кувелье, Г., и Ренар, К.М.Г.С. (2007). Текстура пюре из фруктов и овощей. Steward Postharvest Ред. 3, 1–14.
Google Scholar
Контадор, Л., Шинья, П., и Инфанте, Р. (2015). Фенотипирование текстуры свежих мясистых фруктов. Sci. Hortic. 193, 40–46.
Google Scholar
Корнуо, В., Баффетто, Ф., Маркус, С.E., Crépeau, M.-J., Guillon, F., Ralet, M.-C., et al. (2017). LM6-M: крысиное моноклональное антитело с высокой авидностью к пектиновому αα-1,5-L-арабинану. bioRxiv 161604.
Google Scholar
Cornuault, V., Buffetto, F., Rydahl, M.G., Marcus, S.E., Torode, T.A., Xue, J., et al. (2015). Моноклональные антитела указывают на низкое содержание связей между гетероксиланом и другими гликанами стенок растительных клеток. Планта 242, 1321–1334. DOI: 10.1007 / s00425-015-2375-4
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Корнуо, В., Мэнфилд, И. В., Ралет, М. К., и Нокс, Дж. П. (2014). Хроматография с обнаружением эпитопа: метод анализа структурной неоднородности и взаимосвязей гликанов матрикса клеточной стенки растений. Завод Дж. 78, 715–722. DOI: 10.1111 / tpj.12504
PubMed Аннотация | CrossRef Полный текст | Google Scholar
D’Hont, A., Denoeud, F., Aury, J.M., Baurens, F.C, Carreel, F., Garsmeur, O., et al. (2012). Геном банана (Musa acuminata) и эволюция однодольных растений. Природа 488, 213–217. DOI: 10.1038 / природа11241
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Эспиноза, Л., То, Н., Симоно, Р., Ренар, К. М. Г. К., Биау, Н. , и Кувелье, Г. (2011). Влияние обработки на реологические, структурные и сенсорные свойства яблочного пюре. Proc. Food Sci. 1, 513–520.
, и Кувелье, Г. (2011). Влияние обработки на реологические, структурные и сенсорные свойства яблочного пюре. Proc. Food Sci. 1, 513–520.
Google Scholar
Дженовезе, Д. Б. (2012). Реология сдвига твердых сфер, дисперсных и агрегатных суспензий и композитов наполнитель-матрица. Adv. Coll. Интерфейс Sci. 17, 1–16. DOI: 10.1016 / j.cis.2011.12.005
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хань Ю., Чжу, К., Чжан, З., Мэн, К., Хоу, Ю., Бан, К., и др. (2015). Анализ генов ксилоглюкан-эндотрансгликозилазы / гидролазы (XTH) и различных ролей изоферментов во время развития плодов хурмы и послеуборочного размягчения. PLoS One 10: e0123668. DOI: 10.1371 / journal.pone.0123668
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Джонс, Л., Сеймур, Г. Б., и Нокс, Дж. П. (1997). Локализация пектинового галактана в клеточных стенках томата с использованием моноклональных антител, специфичных к (1-> 4) -бета-D-галактану. Plant Physiol. 113, 1405–1412.
PubMed Аннотация | Google Scholar
Кроп, Э. М., Хетерингтон, М., М., Холмс, М., Микель, С., и Саркар, А. (2019). О связи реологии и оральной трибологии с сенсорными свойствами гидрогелей. Food Hydrocoll. 88, 101–113.
Google Scholar
Лагуна, Л., Фаррелл, Г., Брайант, М., Морина, А., Саркар, А. (2017). Связь реологии и трибологии товарных молочных коллоидов с сенсорным восприятием. Food Funct. 8, 563–573.
Google Scholar
Лоу Д. Ю., Уильямс Б. А., Дарси Б. Р., Фланаган Б. М. и Гидли М. Дж. (2015). Ферментация жевательных манго и бананов in vitro: размер частиц, крахмал и сосудистые волокна. Food Funct. 6, 2464–2474. DOI: 10.1039 / c5fo00363f
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Маркус, С.Э., Блейк, А. В., Бенианс, Т. А., Ли, К. Дж., Пойзер, К., Дональдсон, Л. и др. (2010). Ограниченный доступ белков к полисахаридам маннана в стенках интактных растительных клеток. Плант Дж. 64, 191–203. DOI: 10.1111 / j.1365-313X.2010.04319.x
Плант Дж. 64, 191–203. DOI: 10.1111 / j.1365-313X.2010.04319.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Моелантс, К. Р. Н., Кардинаэлс, Р., Ван Буггенхаут, С., Ван Лой, А. М., Молденерс, П., и Хендрикс, М. Е. (2014). Обзор взаимосвязи между обработкой, структурой пищи и реологическими свойствами пищевых суспензий на основе тканей растений. Компр. Rev. Food Sci. Food Saf. 13, 241–260.
Google Scholar
Муда П., Сеймур Г. Б., Эррингтон Н. и Такер Г. А. (1995). Изменения состава полимеров клеточной стенки во время созревания плодов манго. Carbohydr. Polym. 26, 255–260.
Google Scholar
Намби В. Э., Тангавел К. и Йесудас Д. М. (2015). Научная классификация периода созревания и разработка шкалы цветности индийского манго ( Mangifera indica L.) с помощью многомерного кластерного анализа. Sci. Hortic. 193, 90–98.
Google Scholar
Онг, Дж. Дж., Стил, К. М. и Дуйзер, Л. М. (2018). Опровержение предположений относительно скорости сдвига в полости рта во время обработки полости рта и глотания, основанных на сенсорном тестировании с загущенными жидкостями. Food Hydrocoll. 84, 173–180. DOI: 10.1016 / j.foodhyd.2018.05.043
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ордас-Ортис, Дж. Дж., Маркус, С.Э., и Нокс, Дж. П. (2009). Анализ микроструктуры клеточной стенки указывает на участие полисахаридов гемицеллюлозы в адгезии клеток в паренхиме перикарпия плодов томата. Мол. Завод 2, 910–921. DOI: 10.1093 / mp / ssp049
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Орфила, К., Сеймур, Г. Б., Уиллатс, В. Г. Т., Хаксэм, И. М., Джарвис, М. К., Довер, К. Дж. И др. (2001). Изменен гомогалактуронан средней ламели и нарушено отложение (1 -> 5) -альфа-L-арабинан в околоплоднике Cnr, созревающего мутанта томата. Plant Physiol. 126, 210–221.
PubMed Аннотация | Google Scholar
Паниагуа К. , Поза С., Моррис В. Дж., Кирби А. Р., Кесада М. А. и Меркадо Дж. А. (2014). Смягчение плодов и разборка пектина: обзор наноструктурных модификаций пектина, оцененных с помощью атомно-силовой микроскопии. Ann. Бот. 114, 1375–1383. DOI: 10.1093 / aob / mcu149
, Поза С., Моррис В. Дж., Кирби А. Р., Кесада М. А. и Меркадо Дж. А. (2014). Смягчение плодов и разборка пектина: обзор наноструктурных модификаций пектина, оцененных с помощью атомно-силовой микроскопии. Ann. Бот. 114, 1375–1383. DOI: 10.1093 / aob / mcu149
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Pattathil, S., Avci, U., Болдуин, Д., Свеннес, А.Г., Макгилл, Дж. А., Поппер, З., и др. (2010). Комплексный набор моноклональных антител, направленных на гликаны клеточной стенки растений. Plant Physiol. 153, 514–525. DOI: 10.1104 / стр.109.151985
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Педерсен, Х. Л., Фангель, Дж. У., Макклири, Б., Рузански, К., Райдал, М. Г., Ралет, М.-К., и др. (2012). Универсальные микроматрицы олигосахаридов высокого разрешения для гликобиологии растений и исследования клеточных стенок. J. Biol. Chem. 287, 39429–39438. DOI: 10.1074 / jbc.M112.396598
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Посе, С., Паниагуа, К., Матас, А. Дж., Ганнинг, А. П., Моррис, В. Дж., Кесада, М. А. и др. (2018). Наноструктурный вид процесса разборки клеточной стенки во время созревания плодов и послеуборочного хранения с помощью атомно-силовой микроскопии. Trends Food Sci. Technol. 87, 47–58.
Google Scholar
Прадо, С. Б.Р. Д., Мелфи, П. Р., Кастро-Алвес, В. К., Броетто, С. Г., Араужо, Э. С., Насименто, Дж. Р. О. Д. и др. (2016). Физиологическая деградация пектина в клеточных стенках папайи: высвобождение длинноцепочечных галактуронанов, полученных из нерастворимых фракций, во время послеуборочного созревания плодов. Перед. Plant Sci. 7: 1120. DOI: 10.3389 / fpls.2016.01120
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Рабити Д., Орфила К., Холмс М., Бордони А. и Саркар А. (2018).Пероральная обработка сырых томатов in vitro: новое понимание роли эндогенных ферментов плодов. J. Текстурный стержень. 49, 351–358. DOI: 10.1111 / jtxs.12338
J. Текстурный стержень. 49, 351–358. DOI: 10.1111 / jtxs.12338
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Radotić, K., Roduit, C., Simonovi, J., Hornitschek, P., Fankhauser, C., Mutavdži, D., et al. (2012). Томография жесткости с помощью атомно-силовой микроскопии на живых клетках арабидопсиса thaliana выявляет механические свойства поверхностных и глубоких слоев клеточной стенки во время роста. Biophys. J. 103, 386–394. DOI: 10.1016 / j.bpj.2012.06.046
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Роуз, Дж. К. С., и Беннетт, А. Б. (1999). Совместная разборка целлюлозно-ксилоглюкановой сети стенок растительных клеток: параллели между размножением клеток и созреванием плодов. Trends Plant Sci. 4, 176–183.
PubMed Аннотация | Google Scholar
Ruthes, A.C, Martínez-Abad, A., Tan, H.-T., Bulone, V., and Vilaplana, F.(2017). Последовательное фракционирование ферулоилированных гемицеллюлоз и олигосахаридов из пшеничных отрубей с использованием субкритической воды и ксиланолитических ферментов. Green Chem. 19, 1919–1931.
Google Scholar
Саркар, А., Андабло-Рейес, Э., Брайант, М., Доусон, Д., и Невилл, А. (2019). Смазка мягких поверхностей полости рта. Curr. Opin. Coll. Интерфейс Sci. 39, 61–75.
Google Scholar
Саркар, А., Канти, Ф., Гулотта, А., Мюррей, Б.С., и Чжан, С. (2017). Водная смазка, структура и реологические свойства частиц микрогеля сывороточного протеина. Langmuir 33, 14699–14708. DOI: 10.1021 / acs.langmuir.7b03627
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Шендель Р. Р., Мейер М. Р. и Бунзель М. (2016). Количественное профилирование боковых цепей ферулоилированного арабиноксилана из клеточных стенок злаков. Перед. Plant Sci. 6: 1249. DOI: 10.3389 / fpls.2015.01249
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Шига Т. М., Карпита, Н. К., Лайоло, Ф. М., и Корденунси-Лысенко, Б. Р. (2017). Два сорта банана различаются по составу потенциально иммуномодулирующего маннана и арабиногалактана. Carbohydr. Polym. 164, 31–41. DOI: 10.1016 / j.carbpol.2017.01.079
М., Карпита, Н. К., Лайоло, Ф. М., и Корденунси-Лысенко, Б. Р. (2017). Два сорта банана различаются по составу потенциально иммуномодулирующего маннана и арабиногалактана. Carbohydr. Polym. 164, 31–41. DOI: 10.1016 / j.carbpol.2017.01.079
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Солтани М., Алимардани Р. и Омид М. (2010). Прогноз качества бананов на стадии созревания с помощью системы измерения емкости. Aust. J. Crop Sci. 4, 443–447.
Google Scholar
Стокс, Дж. Р., Бём, М. В., и Байер, С. К. (2013). Обработка полости рта, текстура и ощущение во рту: от реологии до трибологии и не только. Curr. Opin. Coll. Интерфейс Sci. 18, 349–359.
Google Scholar
Сувонсихон, С., Чемберс, И., Конгпенсук, В., и Упадиссакун, К. (2012). Сенсорная лексика манго в зависимости от сорта и стадии спелости. J. Sens. Stud. 27, 148–160.
Google Scholar
Торрес, О., Андабло-Рейес, Э., Мюррей, Б.С., Саркар, А. (2018). Частицы эмульсионного микрогеля как высокоэффективные био-смазки. ACS Appl. Матер. Интерфейсы 10, 26893–26905. DOI: 10.1021 / acsami.8b07883
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Валенте, М., Рибейр, Ф., Селф, Г., Бертио, Л., и Ассемат, С. (2011). Инструментальная и сенсорная характеристика текстуры плодов манго. J. Food Qual. 34, 413–424.
Google Scholar
Ван, Д., Йейтс, Т. Х., Улуисик, С., Роуз, Дж. К. К., Сеймур, Г. Б. (2018). Смягчение фруктов: новый взгляд на роль пектина. Trends Plant Sci. 23, 302–310. DOI: 10.1016 / j.tplants.2018.01.006
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Здунек А., Куренда А. (2013). Определение упругих свойств клеток плодов томата с помощью атомно-силового микроскопа. Датчики 13, 12175–12191. DOI: 10.3390 / s1305
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чжан, Б. , Селуэй, Н., Шелат, К. Дж., Дхитал, С., Стокс, Дж. Р. и Гидли, М. Дж. (2017). Трибология суспензий набухших крахмальных гранул кукурузы и картофеля. Carbohydr. Polym. 155, 128–135. DOI: 10.1016 / j.carbpol.2016.08.064
, Селуэй, Н., Шелат, К. Дж., Дхитал, С., Стокс, Дж. Р. и Гидли, М. Дж. (2017). Трибология суспензий набухших крахмальных гранул кукурузы и картофеля. Carbohydr. Polym. 155, 128–135. DOI: 10.1016 / j.carbpol.2016.08.064
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чжан Т., Чжэн Ю. и Косгроув Д. Дж. (2016). Пространственная организация микрофибрилл целлюлозы и матричных полисахаридов в первичных клеточных стенках растений, как показано с помощью многоканальной атомно-силовой микроскопии. Plant J. 85, 179–192. DOI: 10.1111 / tpj.13102
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Самосборка нанодисков термодинамически обратима и управляема
Многие высокоупорядоченные сложные системы образуются в результате спонтанной самосборки более простых субъединиц. Важным биофизическим инструментом, который полагается на самосборку, является система Nanodisc, которая находит широкое применение в качестве нативной среды для изучения мембранных белков.Нанодиски самоорганизуются из растворенных в детергенте смесей фосфолипидов и сконструированных спиральных белков, называемых белками мембранного каркаса (MSP). Удаление детергента приводит к образованию наноразмерных бислоев, стабилизированных двумя «поясами» MSP. Несмотря на их многочисленные приложения в биологии и вклад многих лабораторий по всему миру, мало что известно о процессе самосборки, например о том, когда образуется бислой или когда MSP связывается с липидами. Мы используем флуоресценцию и оптическую спектроскопию для исследования самосборки при различных равновесиях, определяемых концентрацией детергента.Мы показываем, что бислой начинает формироваться ниже критической мицеллярной концентрации детергента (10 мМ), а ассоциация MSP и липидов начинается при более низких уровнях детергента, показывая зависимость от концентраций MSP и липидов. Следование процессу растворения путем добавления детергента к очищенным нанодискам демонстрирует, что самосборка обратима. Наши данные демонстрируют, что самосборка Nanodisc экспериментально доступна, и что контроль концентрации детергента позволяет точно контролировать реакцию самосборки.Это улучшенное понимание самосборки может привести к лучшему функциональному включению ранее не поддающихся лечению мембранных белков-мишеней.
Наши данные демонстрируют, что самосборка Nanodisc экспериментально доступна, и что контроль концентрации детергента позволяет точно контролировать реакцию самосборки.Это улучшенное понимание самосборки может привести к лучшему функциональному включению ранее не поддающихся лечению мембранных белков-мишеней.
У вас есть доступ к этой статье
Подождите, пока мы загрузим ваш контент… Что-то пошло не так. Попробуй еще раз?Невязанное микророботизированное кодирование трехмерного состава материала
Изготовление микрогелей
Раствор предшественника ПЭГ был приготовлен растворением (20%, мас. / Мас.) Диметакрилата полиэтиленгликоля (ПЭГДМА; МВ 1000; Polysciences) в лаборатории Дульбекко. фосфатно-солевой буфер (DPBS, GIBCO).Раствор предшественника GelMA получали растворением (5%, мас. / Мас.) Пенообразного порошка GelMA в DPBS (GIBCO). Затем к раствору форполимера добавляли 2-гидрокси-1- (4- (гидроксиэтокси) фенил) -2-метил-1-пропанон фотоинициатор (1%, мас. / Мас., Irgacure 2959; CIBA Chemicals). В данном исследовании мы использовали фотошаблоны нескольких геометрических форм (рис. 2). Каплю 50 мкл раствора фото-сшиваемого форполимера пипеткой наносили на предметное стекло, покрытое покровным стеклом и разделенное разделителями (покровное стекло 25 × 25 мм 2 , толщина: 150 мкм). Фотошаблоны помещали на покровное стекло между УФ-светом и форполимером. На каплю помещали еще одно покровное стекло размером 25 × 25 мм 2 . Микрогели изготавливали, подвергая раствор гелевого форполимера УФ-свету (500 мВт; на высоте 50 мм над микрогелями) в течение 30 с, чтобы полимеризация происходила на поверхности предметного стекла. Затем фотошаблон и покровное стекло были удалены. С помощью скальпеля удаляли микрогели и оставляли для пропитывания в растворе DPBS в стандартной чашке Петри диаметром 60 мм (Fisher Scientific).
Фотошаблоны помещали на покровное стекло между УФ-светом и форполимером. На каплю помещали еще одно покровное стекло размером 25 × 25 мм 2 . Микрогели изготавливали, подвергая раствор гелевого форполимера УФ-свету (500 мВт; на высоте 50 мм над микрогелями) в течение 30 с, чтобы полимеризация происходила на поверхности предметного стекла. Затем фотошаблон и покровное стекло были удалены. С помощью скальпеля удаляли микрогели и оставляли для пропитывания в растворе DPBS в стандартной чашке Петри диаметром 60 мм (Fisher Scientific).
Изготовление кремниевых чиплетов
Кремниевые чиплеты (1 мм × 1 мм × 0,1 мм) были изготовлены из 2-дюймовых кремниевых пластин (University Wafer, MA) с помощью автоматической пилы для резки кубиками (модель DAD321, Disco Corp., Токио, Япония) .
Изготовление строительных блоков PDMS
Строительные блоки PDMS были изготовлены с использованием фотолитографии и быстрого прототипирования с небольшими модификациями 26 (дополнительный рис. 2). Вкратце, мастер-форма SU-8 толщиной 500 мкм была изготовлена на 4-дюймовой кремниевой пластине с использованием стандартной фотолитографии.Перед использованием эталонная форма была покрыта слоем трихлор (1H, 1H, 2H, 2H-перфтороктил) силана (Sigma) для облегчения высвобождения структуры PDMS. Форполимер ПДМС получали смешиванием предшественника ПДМС и отвердителя в соотношении 11: 1. Форма из PDMS была изготовлена путем отверждения форполимера на мастер-форме при 80 ° C в течение 1 часа. Перед использованием форма PDMS была покрыта слоем трихлор (1H, 1H, 2H, 2H-перфтороктил) силана для облегчения высвобождения блоков PDMS. Форполимер ПДМС для строительных блоков был приготовлен путем смешивания предшественника ПДМС и отвердителя в соотношении 9: 1.Sudan Red G (Sigma) смешивали с форполимером с конечной концентрацией 0,5% (мас. / Мас.) Для визуализации блоков PDMS. Форполимер выливали в форму из ПДМС с последующим удалением избытка форполимера с помощью лезвия бритвы. Форполимер отверждали при 80 ° C в течение 1 ч в печи. Окончательные блоки PDMS были освобождены путем сгибания пресс-формы PDMS. Блоки PDMS были сделаны гидрофильными обработкой кислородной плазмой перед экспериментами по кодированию.
Форполимер отверждали при 80 ° C в течение 1 ч в печи. Окончательные блоки PDMS были освобождены путем сгибания пресс-формы PDMS. Блоки PDMS были сделаны гидрофильными обработкой кислородной плазмой перед экспериментами по кодированию.
Экспериментальная установка
Изготовленные строительные блоки (например, микрогели, силиконовые чиплеты, блоки PDMS) суспендировали в растворе DPBS в чашках Петри.Они были закодированы в различные сложные формы с помощью системы из восьми электромагнитов (рис. 1c). Строительные блоки по очереди направлялись в заранее определенное место путем прямого толкания магнитного непривязанного микроробота. Магнитные микророботы приводятся в действие набором независимых электромагнитных катушек, выровненных по направлению к общей центральной точке с открытым пространством ~ 10,4 см. Катушки работают с воздушным или железным сердечником, в зависимости от желаемых магнитных полей и градиентов. Максимальные поля, создаваемые системой, работающей при максимальном токе (19 А каждое), равны 8.3 мТл с использованием воздушных сердечников и 24,4 мТл с использованием железных сердечников. Точно так же максимальные пространственные градиенты поля составляют 0,34 Тл м -1 с использованием воздушных сердечников и 1,02 Тл м -1 с использованием железных сердечников. Поля и градиенты поля линейно связаны с токами катушки и измеряются с помощью датчика Холла (Allegro A1321) с погрешностью около 0,1 мТл. Управление токами, управляющими электромагнитными катушками, осуществляется ПК с системой сбора данных с полосой управления 20 кГц, а катушки питаются от линейных электронных усилителей (SyRen 25).Магнитными микророботами можно управлять с помощью магнитных катушек, окружающих рабочее пространство. Общий магнитный момент T и сила F , которые управляют этими взаимодействиями, составляют:
для микроробота с намагниченностью M , где B — полное магнитное поле от катушек. Приведение в действие микроробота осуществляется комбинацией магнитных сил, которые непосредственно притягивают плоскость подложки, и магнитных моментов, которые действуют, чтобы разрушить поверхностное трение и сцепление с поверхностью. Эти крутящие моменты применяются в виде пилообразной волны с частотой 10–50 Гц и приводят к дискретным шагам микророботов размером около 10–300 мкм в зависимости от величины и частоты импульса.
Эти крутящие моменты применяются в виде пилообразной волны с частотой 10–50 Гц и приводят к дискретным шагам микророботов размером около 10–300 мкм в зависимости от величины и частоты импульса.
Изготовление магнитных микророботов
В этой работе были изготовлены отдельные микророботы с магнитной твердостью, сохраняющей свою внутреннюю намагниченность в отсутствие внешнего магнитного поля. Микророботы были изготовлены в периодическом процессе с использованием мягкой фотолитографии и методов формования 27 .Микророботы были составлены из частиц NdFeB в полиуретановой (BJB Enterprise, ST-1087) матрице, детали изготовления приведены в ссылке. 28. Короче говоря, формы из мягкой резины были изготовлены из точных копий элементов SU-8, нанесенных с помощью фотолитографии на кремниевой подложке (дополнительный рис. 2). Микророботы были созданы из формы путем заливки суспензии полиуретана NdFeB в форму. Из-за высокой магнитной коэрцитивности NdFeB (то есть для размагничивания NdFeB требуются поля более 600 мТл) эти микророботы не подвергались размагничиванию из-за относительно слабых полей, примененных в этой работе.Номинальная объемная намагниченность M микроробота составляла около 50 кА м -1 , как измерено в магнитометре с переменным градиентом силы (Princeton Measurements MicroMag 2900). Процесс формования всех микророботов был подвержен отклонениям в геометрии микророботов (примерно до 10% от номинала), но метод управления движением не был чувствителен к небольшим геометрическим изменениям.
Вторичное сшивание для стабилизации кодированной конструкции
Микрогели были закодированы на нижней поверхности резервуара, и 10 мкл раствора форполимера были добавлены к конечной конструкции.Микрогели подвергали вторичному сшиванию УФ-излучением в течение 20 с для стабилизации формы структуры.
Инкапсуляция клеток
Фибробласты NIH 3T3 культивировали в среде Игла, модифицированной Дульбекко (DMEM; Sigma-Aldrich) с добавлением 10% фетальной бычьей сыворотки (FBS; GIBCO), в инкубаторе с 5% CO 2 -увлажненным при 37 ° C. . Чтобы собрать и инкапсулировать клетки, клетки сначала трипсинизировали 1% трипсином (GIBCO) и центрифугировали при 1000 об / мин. на 5 мин. Клетки суспендировали в 5% мас. / Об. GelMA, растворенном в PBS (Gibco, Invitrogen) при плотности 1 × 10 7 на мл, и смешивали с 0.5% мас. / Об. Фотоинициатора (Irgacure, Ciba). Затем были изготовлены инкапсулирующие клетки микрогели путем фотошивки с использованием УФ-света мощностью 500 мВт в течение 20 с (рис. 1а). Инкапсулирующие клетки микрогели сначала промывали DPBS, а затем инкубировали с живыми / мертвыми красителями в течение 15 мин. Живые / мертвые красители получали разбавлением 2 мл кальцеина AM и 0,5 мл гомодимера этидия-1 (Molecular Probes) в 1 мл DPBS. Клетки были помечены зеленым с помощью кальцеина AM и помечены красным с помощью красного флуоресцентного клеточного линкера PKh36.Флуоресцентные изображения получали с помощью инвертированного флуоресцентного микроскопа (Nikon, TE2000). Иммуноцитохимические изображения были получены после 4 дней культивирования при 37 ° C, 5% CO 2 в увлажненном инкубаторе.
. Чтобы собрать и инкапсулировать клетки, клетки сначала трипсинизировали 1% трипсином (GIBCO) и центрифугировали при 1000 об / мин. на 5 мин. Клетки суспендировали в 5% мас. / Об. GelMA, растворенном в PBS (Gibco, Invitrogen) при плотности 1 × 10 7 на мл, и смешивали с 0.5% мас. / Об. Фотоинициатора (Irgacure, Ciba). Затем были изготовлены инкапсулирующие клетки микрогели путем фотошивки с использованием УФ-света мощностью 500 мВт в течение 20 с (рис. 1а). Инкапсулирующие клетки микрогели сначала промывали DPBS, а затем инкубировали с живыми / мертвыми красителями в течение 15 мин. Живые / мертвые красители получали разбавлением 2 мл кальцеина AM и 0,5 мл гомодимера этидия-1 (Molecular Probes) в 1 мл DPBS. Клетки были помечены зеленым с помощью кальцеина AM и помечены красным с помощью красного флуоресцентного клеточного линкера PKh36.Флуоресцентные изображения получали с помощью инвертированного флуоресцентного микроскопа (Nikon, TE2000). Иммуноцитохимические изображения были получены после 4 дней культивирования при 37 ° C, 5% CO 2 в увлажненном инкубаторе.
Окрашивающие гидрогели
Для улучшения видимости во время видеозаписи гидрогели окрашивали пищевым красителем (краситель Procion Mx), который состоит из небольших молекул и имеет незначительное взаимодействие с полимером. Краситель продемонстрировал достаточное взаимодействие, чтобы оставаться в гидрогеле в течение достаточно длительного времени для процесса кодирования, и его можно было увидеть во время видеозаписи.
Запись и обработка изображений
Видеозаписи были записаны цифровой камерой (Foculus F0134SB), подключенной к линзам микроскопа с переменным увеличением, обеспечивающим поле обзора до 26 мм × 20 мм при виде сверху. Жизнеспособность клеток оценивалась количественно путем анализа изображений с использованием общедоступной программы NIH ImageJ (разработанной Национальным институтом здравоохранения США и доступной по адресу http://rsb.info.nih. gov/nih/image/). Для расчета скорости микроробота по видео использовался алгоритм отслеживания, основанный на фильтре частиц.Этот алгоритм находит наиболее вероятное положение микроробота и фильтрует его с помощью фильтра Баттерворта нижних частот для уменьшения шума отслеживания.
gov/nih/image/). Для расчета скорости микроробота по видео использовался алгоритм отслеживания, основанный на фильтре частиц.Этот алгоритм находит наиболее вероятное положение микроробота и фильтрует его с помощью фильтра Баттерворта нижних частот для уменьшения шума отслеживания.
Иммуноцитохимическое окрашивание гелей, сшитых УФ-светом.
Пролиферирующие клетки, инкапсулированные в гидрогели, выявляли с помощью иммуноцитохимии Ki67. Собранные гели, инкапсулирующие клетки 3T3 (рис. 5a – g), фиксировали 1% параформальдегидом в течение 1 ч при комнатной температуре и промывали. Гели были проницаемы с 0,3% Triton X-100 (Sigma) в 1% BSA (Sigma) в течение минимум 1 часа при комнатной температуре.Гели окрашивали Ki67 (Ab16667, Abcam) в течение ночи при 4 ° C. После отмывки гели инкубировали с вторичными антителами козы против кроличьего Alexa Fluor 564 (A11011, Invitrogen) в течение 2 часов при комнатной температуре. Актиновый цитоскелет окрашивали фаллоидином Alexa Fluor 488 (Invitrogen), а DAPI использовали в качестве контрольного окрашивания ядер. После промывки гидрогели визуализировали под флуоресцентным микроскопом (Zeiss AXIO).
Флуоресцентное окрашивание для гетерогенных ансамблей
HUVEC, фибробласты 3T3 и инкапсулирующие кардиомиоциты куриные гидрогели фиксировали 1% параформальдегидом в течение 1 ч при комнатной температуре и промывали.Гели были проницаемы с 0,3% Triton X-100 (Sigma) в 1% блокирующем растворе BSA (Sigma) в течение минимум 1 часа при комнатной температуре. Кардиомиоциты окрашивали гомодимером этидия-1 (Life Technologies). HUVEC окрашивали фаллоидином Alexa Fluor 488 (Life Technologies). Фибробласты 3T3 окрашивали DAPI (Life Technologies). После отмывки изображения гидрогелей были получены с помощью флуоресцентного микроскопа (Zeiss AXIO). Затем окрашенные инкапсулирующие клетки гели были собраны микророботом с высокой точностью (рис.5h – q).
МТТ-анализы
Клетки NIH 3T3 культивировали в среде Игла, модифицированной Дульбекко (DMEM) с добавлением 10% фетальной бычьей сыворотки и 1% смеси пенициллин-стрептомицин в увлажненной и содержащей 5% CO 2 -содержащей атмосфере при 37 ° C .
Для анализа жизнеспособности МТТ клетки высевали в 96-луночный планшет с плотностью 10 5 клеток на мл -1 (в общем объеме 100 мкл суспензии клеток на лунку). Когда клетки прикрепляли через 4 ч, в каждую лунку помещали по одному микророботу.Клетки инкубировали 5, 20 и 60 мин при 37 ° C. После инкубации микророботы были удалены, и среда для культивирования клеток была отсосана из лунок для всех образцов. Затем добавляли свежую среду для культивирования клеток и реагент МТТ (10% об. / Об.) И инкубировали в течение 4 часов. По истечении 4 ч образовавшийся формазан растворяли в 100 мкл растворяющего МТТ реагента SDS. Поглощение индуцированного красителя формазана измеряли с помощью многофункционального микропланшетного ридера BMG FLUOstar Galaxy на следующий день. Все определения проводили в шести повторах для каждого образца и проводили три независимых эксперимента.Анализы пролиферации МТТ проводили через 0, 1, 3, 5 и 7 дней после удаления микророботов из клеточных суспензий. Клетки без присутствия микророботов использовали в качестве контроля.
Статистические методы
Измерения интенсивности флуоресценции, полученные в результате анализа МТТ ( n = 12–15), статистически сравнивали с использованием параметрического одностороннего дисперсионного анализа (ANOVA) с тестом Tukey posthoc для множественных сравнений. Порог статистической значимости был установлен на 0.05 ( P <0,05).
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались. Чтобы принять файлы cookie с этого сайта, нажмите кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie. Вы должны отключить приложение при входе в систему или проконсультироваться с системным администратором.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie потребует, чтобы сайт создавал новый сеанс для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файле cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файлах cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
Пересечение между разборкой клеточной стенки, созреванием и восприимчивостью плодов к Botrytis cinerea
Аннотация
Созревание плодов характеризуется процессами, которые изменяют текстуру и вкус, а также резким увеличением восприимчивости к некротрофным патогенам, таким как Botrytis cinerea .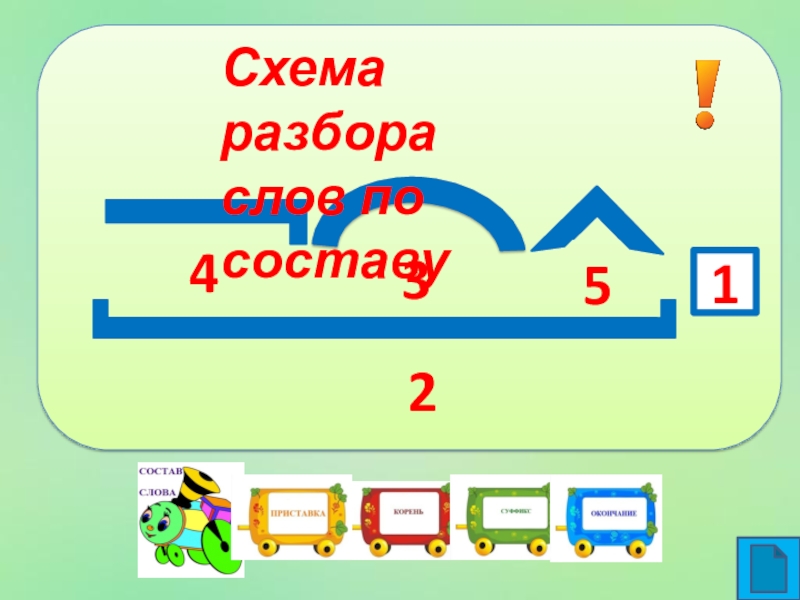 Разборка основных структурных полисахаридов клеточной стенки (CW) является важным процессом, связанным с созреванием и способствует размягчению плодов.У томатов полигалактуроназа (PG) и экспансин (Exp) входят в число белков CW, которые совместно участвуют в разборке CW, связанной со созреванием. Чтобы определить, влияет ли эндогенная разборка CW на регулируемое созреванием увеличение восприимчивости к некротропным патогенам, была оценена чувствительность B. cinerea в трансгенных плодах с подавленной экспрессией полигалактуроназы ( LePG ) и экспансина ( LeExp1 ). Подавление либо LePG , либо LeExp1 по отдельности не снижало восприимчивость, но одновременное подавление обоих резко снижало восприимчивость созревающих плодов до B.cinerea , как измерено по накоплению биомассы грибов и по развитию очагов мацерации. Эти результаты демонстрируют, что изменение разборки эндогенных CW растений во время созревания влияет на течение инфекции B. cinerea , возможно, путем изменения структуры или доступности субстратов CW для ферментов, разрушающих CW патогенов. Признание роли связанного со созреванием метаболизма ХС в восприимчивости к патогенам после уборки урожая может быть полезным при разработке и разработке стратегий по ограничению потерь патогенов во время хранения, обработки и распределения фруктов.
Разборка основных структурных полисахаридов клеточной стенки (CW) является важным процессом, связанным с созреванием и способствует размягчению плодов.У томатов полигалактуроназа (PG) и экспансин (Exp) входят в число белков CW, которые совместно участвуют в разборке CW, связанной со созреванием. Чтобы определить, влияет ли эндогенная разборка CW на регулируемое созреванием увеличение восприимчивости к некротропным патогенам, была оценена чувствительность B. cinerea в трансгенных плодах с подавленной экспрессией полигалактуроназы ( LePG ) и экспансина ( LeExp1 ). Подавление либо LePG , либо LeExp1 по отдельности не снижало восприимчивость, но одновременное подавление обоих резко снижало восприимчивость созревающих плодов до B.cinerea , как измерено по накоплению биомассы грибов и по развитию очагов мацерации. Эти результаты демонстрируют, что изменение разборки эндогенных CW растений во время созревания влияет на течение инфекции B. cinerea , возможно, путем изменения структуры или доступности субстратов CW для ферментов, разрушающих CW патогенов. Признание роли связанного со созреванием метаболизма ХС в восприимчивости к патогенам после уборки урожая может быть полезным при разработке и разработке стратегий по ограничению потерь патогенов во время хранения, обработки и распределения фруктов.
Растительные клетки окружены сложными композитными стенками, которые являются важными преформированными барьерами для патогенной инфекции (1, 2). Многие сапрофитные и патогенные организмы растений выделяют ферменты, разрушающие клеточную стенку (CWDE), которые специально нацелены на компоненты клеточной стенки растений (CW), так что патоген может проникать в хозяина и получать питательные вещества из переваренного материала стенок и клеточного содержимого (3) . Было показано, что некоторые CWDE являются факторами вирулентности (4–9).
Во время созревания многие фрукты, в том числе томаты, разбирают компоненты CW, что приводит к изменению реологических свойств CW и размягчению спелых фруктов (10). CW плодов томата состоит из двух взаимодействующих полисахаридных сетей, целлюлозно-гемицеллюлозной сети (Cel-Hem), содержащей основные элементы, придающие силу, встроенные в коэкстенсивную пектиновую сеть (11). Во время созревания белки, модифицирующие CW, в том числе полигалактуроназы (PG) и экспансины (Exps), действуют совместно, разрушая полимерные сети и тем самым способствуя размягчению плодов. PG гидролизуют гомогалактуронановые полимеры, которые являются основными компонентами пектиновой матрицы, и считается, что Exps влияют на взаимодействия внутри сети Cel-Hem, хотя никакие ферментативные функции не были окончательно связаны с Exps (12, 13).Хотя участие PG и Exps в метаболизме стенки, связанном со созреванием, очевидно (10, 14), мало что известно о том, как эти CWDE взаимодействуют с целью разборки CW.
CW плодов томата состоит из двух взаимодействующих полисахаридных сетей, целлюлозно-гемицеллюлозной сети (Cel-Hem), содержащей основные элементы, придающие силу, встроенные в коэкстенсивную пектиновую сеть (11). Во время созревания белки, модифицирующие CW, в том числе полигалактуроназы (PG) и экспансины (Exps), действуют совместно, разрушая полимерные сети и тем самым способствуя размягчению плодов. PG гидролизуют гомогалактуронановые полимеры, которые являются основными компонентами пектиновой матрицы, и считается, что Exps влияют на взаимодействия внутри сети Cel-Hem, хотя никакие ферментативные функции не были окончательно связаны с Exps (12, 13).Хотя участие PG и Exps в метаболизме стенки, связанном со созреванием, очевидно (10, 14), мало что известно о том, как эти CWDE взаимодействуют с целью разборки CW.
Созревание плодов характеризуется резким увеличением восприимчивости к некротрофным патогенам, что приводит к большим экономическим потерям скоропортящихся продуктов садоводства (14, 15). Метаболизм CW многих съедобных плодов широко изучался в контексте созревания (10), но вклад разборки CW, связанной со созреванием, на восприимчивость к патогенам экспериментально не определен.Обычно предполагается, что саморазборка стенки плода способствует восприимчивости, потому что многие из CWDE фруктов, которые участвуют в метаболизме стенки плода, также продуцируются вторгающимися патогенами и секретируются в ткани плода (16). Плоды томатов с нарушением созревания с мутациями Nr или или демонстрируют снижение размягчения, связанного со созреванием, и пониженную восприимчивость к патогенам (17, 18). Однако у этих генотипов отсутствие нормального метаболизма CW, связанного со созреванием, является лишь одним из многих затронутых процессов, поэтому нельзя сделать вывод, что CW-метаболизм мутантов per se способствует снижению восприимчивости к патогенам (18).Плоды, в которых подавлена экспрессия генов, кодирующих CWDE, таких как PG и пектинметилэстераза, по-видимому, имеют повышенную целостность ткани на стадиях полного созревания или перезревания (18–21).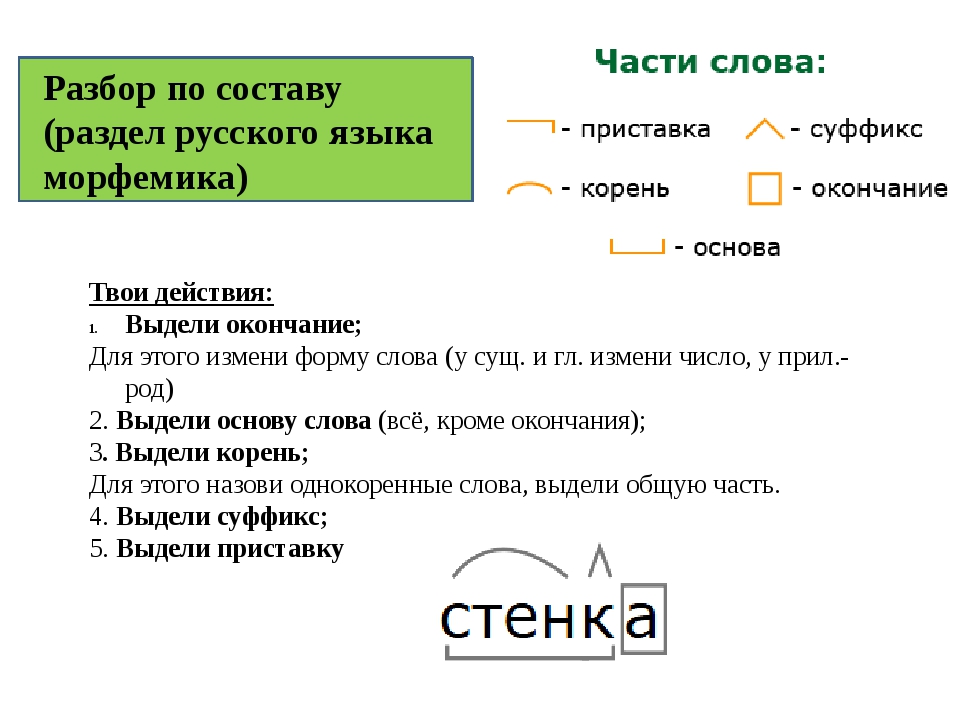 Подавление гена Exp, связанного со созреванием плодов томата, LeExp1 , привело к получению более плотных плодов с увеличенным сроком хранения (22), но не снизило восприимчивость к Botrytis cinerea и Alternaria alternata (4).
Подавление гена Exp, связанного со созреванием плодов томата, LeExp1 , привело к получению более плотных плодов с увеличенным сроком хранения (22), но не снизило восприимчивость к Botrytis cinerea и Alternaria alternata (4).
Сложность и структурная пластичность CW усложняют подходы, разработанные для понимания взаимосвязи между метаболизмом стенок, связанным со созреванием, размягчением плодов и повышенной восприимчивостью к патогенам.Это может объяснить, почему исследования, основанные на модифицированной экспрессии отдельных CWDEs в созревающих фруктах, не привели к значительному положительному воздействию ни на восприимчивость, ни на размягчение. Здесь мы исследовали влияние одновременного подавления экспрессии LePG и LeExp1 в созревающих плодах томата на восприимчивость к B. cinerea и твердость плода. Результаты демонстрируют, что совместное действие этих двух белков CW является важной детерминантой увеличения чувствительности к некротрофным патогенам, связанного со созреванием плодов.
Результаты
Характеристика линий трансгенных томатов.
A трансгенный Solanum lycopersicon cv. Линия Ailsa Craig (AC) с подавленным смыслом для LeExp1 экспрессии (-Exp) (23) и другая линия с антисмысловым подавлением для экспрессии LePG (-PG) (24) были скрещены с получением линий, одновременно подавленных для обеих LeExp1. и LePG (-PG-Exp) (22) .PG-белок (рис.1 A ) и активность (результаты не показаны) не обнаруживаются в красных спелых (RR) плодах -PG или -PG-Exp, а белок Exp значительно снижен в плодах RR -Exp или -PG-Exp. Ни одно из трансгенных растений не показало модифицированного фенотипа или изменений показателей качества спелых плодов, включая содержание глюкозы, фруктозы, каротиноидов или титруемой кислотности (результаты не показаны). Однако степень размягчения плодов трансгенных линий во время созревания существенно изменилась (рис. 1 B ): плоды -PG-Exp были тверже, чем плоды AC, -PG или -Exp дикого типа, хотя плоды -Exp были тверже, чем плоды AC на ранней стадии созревания, аналогично тому, что сообщалось (22, 23 ).
1 B ): плоды -PG-Exp были тверже, чем плоды AC, -PG или -Exp дикого типа, хотя плоды -Exp были тверже, чем плоды AC на ранней стадии созревания, аналогично тому, что сообщалось (22, 23 ).
Экспрессия LePG и LeExp1 влияет на размягчение созревающих плодов томата Ailsa Craig (AC).( A ) Вестерн-блоттинг демонстрирует, что белки, распознаваемые антителами к PG и Exp томатов, присутствуют в красных спелых (RR) AC плодах, что PG отсутствует в плодах -PG, что Exp отсутствует в плодах -Exp и что оба белки отсутствуют в плодах -PG-Exp. ( B ) Измерение силы сжатия каждого плода на 2 мм показывает, что уменьшение PG и Exp у фруктов значительно снижает размягчение на всех стадиях созревания. Твердость измеряли на стадии зрелого зеленого (MG) и на последующих стадиях созревания [брейкер (BR), светло-красный (LR) и RR].Буквы соответствуют значимым различиям между генотипами на каждой стадии созревания; P <0,05.
Чувствительность плодов к
B. cinerea . Одновременное подавление LePG и LeExp1 привело к резкому снижению восприимчивости светло-красных (LR) или RR плодов к B. cinerea [ P <0,005 через 1, 2 и 3 дня после инокуляция (dpi)], тогда как подавление только LePG или LeExp1 не привело к значительному изменению чувствительности (фиг.2 и 3). Чтобы понять, влияют ли спроектированные изменения в метаболизме КС плодов напрямую на рост грибов, количество биомассы B. cinerea на участках инфекции оценивали с помощью моноклонального антитела BC12.CA4, которое распознает эпитоп, локализованный в мицелии (25). Накопление биомассы грибов было значительно снижено, когда LePG и LeExp1 подавлялись по отдельности, и этот эффект был аддитивным, когда экспрессия обоих генов подавлялась (рис.4 А ).
Накопление биомассы грибов было значительно снижено, когда LePG и LeExp1 подавлялись по отдельности, и этот эффект был аддитивным, когда экспрессия обоих генов подавлялась (рис.4 А ).
Снижение PG и Exp существенно снижает восприимчивость к B. cinerea . Плоды LR и RR инокулировали спорами B. cinerea в трех из четырех точек прокола на каждом плоде (четвертый участок был имитирован водой), и симптомы заболевания оценивали при 1, 2 и 3 dpi.У плода проявлялись симптомы болезни, когда на любом засеянном участке развивалось расширяющееся мацеративное (пропитанное водой) поражение, и был показан процент плодов, показывающих какие-либо симптомы заболевания. Было проанализировано около 50 плодов каждого генотипа, и эксперимент проводился как минимум дважды.
Инжир.3.Снижение PG и Exp существенно снижает симптомы серой гнили у привитых LR или RR плодов. Изображение фруктов с разрешением 3 dpi.
Рис. 4. Снижение PG и Exp снижает рост патогенов на плодах, а состав CW из инфицированной ткани демонстрирует, что накопление биомассы грибов снижается на плодах -PG-Exp.( A ) Инфицированные ткани вырезали через 36 и 72 часа после инокуляции (hpi), и оценка биомассы B. cinerea в гомогенатах была основана на интенсивности сигнала (SI) с использованием моноклонального антитела BC12. CA4. Буквы соответствуют значительным различиям между генотипами на каждом hpi; P <0,001. ( B ) AIR получали из инфицированных тканей и анализировали колориметрически на содержание UA и NS, выраженное как отношения UA / NS. Печатные буквы соответствуют значительным различиям между генотипами в каждом hpi; курсивные буквы соответствуют значительным различиям между hpi для каждого генотипа; П <0.001. ( C ) Воздух из плодов RR при 36 и 72 hpi гидролизовали и дериватизировали для определения NS методом ГХ-МС. Показан выходной сигнал детектора ГХ-МС. Показаны времена элюирования пиков производных альдитолацетата. Ара, арабиноза; Ксил, ксилоза; Человек, манноза; Гал, галактоза; Glc, глюкоза; Ino, myo — внутренний стандарт-инозит. Относительное содержание каждого сахара выражается как общее количество ионов. ( I ) AC, 36 л.с. ( II ) переменного тока, 72 л.с. ( III ) −PG − Exp, 36 hpi.( IV ) −PG − Exp, 72 hpi. Повышенные значения Man и Glc указывают на повышенное содержание B. cinerea CW в AC фруктов AIR. Обозначение стержней показано на рис. 1. Б .
CA4. Буквы соответствуют значительным различиям между генотипами на каждом hpi; P <0,001. ( B ) AIR получали из инфицированных тканей и анализировали колориметрически на содержание UA и NS, выраженное как отношения UA / NS. Печатные буквы соответствуют значительным различиям между генотипами в каждом hpi; курсивные буквы соответствуют значительным различиям между hpi для каждого генотипа; П <0.001. ( C ) Воздух из плодов RR при 36 и 72 hpi гидролизовали и дериватизировали для определения NS методом ГХ-МС. Показан выходной сигнал детектора ГХ-МС. Показаны времена элюирования пиков производных альдитолацетата. Ара, арабиноза; Ксил, ксилоза; Человек, манноза; Гал, галактоза; Glc, глюкоза; Ino, myo — внутренний стандарт-инозит. Относительное содержание каждого сахара выражается как общее количество ионов. ( I ) AC, 36 л.с. ( II ) переменного тока, 72 л.с. ( III ) −PG − Exp, 36 hpi.( IV ) −PG − Exp, 72 hpi. Повышенные значения Man и Glc указывают на повышенное содержание B. cinerea CW в AC фруктов AIR. Обозначение стержней показано на рис. 1. Б .
Для определения доли массы CW гриба в общей массе стенки, изолированной от участков инфекции, измеряли уроновую кислоту (UA) и нейтральные сахара (NS), а также оценивали распределение сахаров. Б.cinerea КС содержат в основном NS и небольшое количество UA (отношение UA / NS ≈1 / 12; данные не показаны). Однако КС плодов томата содержат 25–30% УК, а остальное — НС. По мере того как B. cinerea заражение LR и RR AC плодов продолжается, соотношение UA / NS изолированного материала CW инфицированного сайта значительно снижается, но этого не наблюдалось в CW от инфицированных плодов -PG-Exp, инокулированных на любой стадии созревания ( Рис. 4 В ). Препараты для стенок из привитых фруктов содержат как плоды томата, так и B. cinerea стенок полисахаридов, и, таким образом, снижение отношения UA / NS, вероятно, будет отражать как накопление грибковых CW, так и потерю плодовых пектинов, богатых UA, вызванную действием патогенных микроорганизмов или фруктовых CWDE. То, что инфицированные препараты стенки содержат КС грибков, подтверждается повышенным содержанием маннозы и глюкозы в препаратах стенок инфицированных плодов по сравнению с содержанием этих сахаров в стенках, приготовленных из неинфицированных плодов (рис. С ).Глюкоза, образующаяся в результате окисления ацетилглюкозамина во время подготовки образца, и манноза являются основными компонентами B. cinerea CW. Таким образом, оценка накопления грибковой биомассы с использованием обнаружения моноклональных антител вместе с анализом сахаров в CW, выделенных из инфицированных плодов, подтверждает вывод о том, что накопление или рост грибковой массы было значительно больше в AC, чем в плодах -PG-Exp.
cinerea стенок полисахаридов, и, таким образом, снижение отношения UA / NS, вероятно, будет отражать как накопление грибковых CW, так и потерю плодовых пектинов, богатых UA, вызванную действием патогенных микроорганизмов или фруктовых CWDE. То, что инфицированные препараты стенки содержат КС грибков, подтверждается повышенным содержанием маннозы и глюкозы в препаратах стенок инфицированных плодов по сравнению с содержанием этих сахаров в стенках, приготовленных из неинфицированных плодов (рис. С ).Глюкоза, образующаяся в результате окисления ацетилглюкозамина во время подготовки образца, и манноза являются основными компонентами B. cinerea CW. Таким образом, оценка накопления грибковой биомассы с использованием обнаружения моноклональных антител вместе с анализом сахаров в CW, выделенных из инфицированных плодов, подтверждает вывод о том, что накопление или рост грибковой массы было значительно больше в AC, чем в плодах -PG-Exp.
Различия в составе или целостности CW плодов AC или -PG-Exp могут влиять на способность патогена метаболизировать или использовать компоненты CW и, таким образом, могут частично быть ответственными за сниженную восприимчивость плодов -PG-Exp. КС, выделенные из неинфицированных плодов четырех генотипов на стадиях созревания LR и RR, которые использовались для оценки чувствительности к грибам, добавляли в качестве источников питательных веществ для жидких культур B. cinerea . На фиг.5 показано, что рост грибов был в 3 раза больше в культурах, содержащих КС, выделенных из плодов AC, -PG или -Exp LR или RR, чем в культурах, содержащих КС, выделенных из плодов LR или RR -PG-Exp, что позволяет предположить, что LR и RR CW является источником питательных веществ для B.cinerea и что его полезность для роста B. cinerea зависит от кооперативного действия PG и Exp. Никаких различий в росте грибов не наблюдается на КС, приготовленных из зрелых зеленых (MG) плодов любого из генотипов (результаты не показаны).
Рост патогенов снижен в жидких культурах, содержащих -PG-Exp CW, по сравнению с культурами, поставляемыми с AC CW.Мицелии собирали с разрешением 1, 3 и 6 точек на дюйм из культур, содержащих AIR, из неинфицированных LR и RR плодов четырех генотипов. Биомассу грибов измеряли, как описано в легенде на рис. А . Обозначение стержней показано на рис. 1. Б . Буквы соответствуют значительным различиям между генотипами на каждом dpi; P <0,001.
Совместное действие PG и Exp влияет на целостность полисахаридных сетей CW фруктов.
Чтобы определить, какие модификации фракций КС плодов могут объяснять различную восприимчивость к патогенам четырех генотипов, КС были получены из неинфицированных MG, брейкеров, LR и RR плодов и последовательно фракционированы для получения водорастворимых, ионно и ковалентно связанных пектинов, основание -растворимые гемицеллюлозы и нерастворимые остатки целлюлозы. Анализ общего углеводного состава КС не выявил различий между генотипами (результаты не показаны). Когда LePG или LeExp1 подавлялись в AC либо по отдельности, либо в комбинации, солюбилизация пектина, измеряемая как переход менее растворимых полимеров пектина в водорастворимые формы, снижалась, и это было наиболее выражено в стенках от RR. фрукты (рис.6 А ). Подавление только LePG или LeExp1 в других генотипах томатов не вызывало снижения солюбилизации ионно или ковалентно связанного пектина в плодах RR (23, 26). Однако одновременное подавление LePG и LeExp1 привело к резкому снижению деполимеризации водорастворимой фракции (рис. B ) и в других фракциях пектина [вспомогательная информация (SI) Рис.8). Однако никаких изменений в составе сахара гемицеллюлозного полимера, растворимости или распределении по размерам не наблюдалось (данные не показаны), что подтверждает, что Exp не влияет на деполимеризацию ксилоглюканов (23), хотя считается, что белок Exp взаимодействует напрямую с ксилоглюканом. богатая сеть Cel-Hem (13)
B ) и в других фракциях пектина [вспомогательная информация (SI) Рис.8). Однако никаких изменений в составе сахара гемицеллюлозного полимера, растворимости или распределении по размерам не наблюдалось (данные не показаны), что подтверждает, что Exp не влияет на деполимеризацию ксилоглюканов (23), хотя считается, что белок Exp взаимодействует напрямую с ксилоглюканом. богатая сеть Cel-Hem (13)
Уменьшение PG и Exp в созревающих фруктах снижает солюбилизацию и деполимеризацию полисахаридов пектина. Пектины экстрагировали последовательно из LR и RR AIR четырех генотипов с использованием воды, CDTA и Na 2 CO 3 . ( A ) Гистограммы распределения UA в экстрактах пектина. ( B ) Изображения показывают фракционирование методом эксклюзионной хроматографии водорастворимых пектинов (WSF) из плодов AC и -PG-Exp на стадиях созревания LR и RR.Данные показывают, что ни увеличение содержания WSF в плодах, связанное с нормальным созреванием, ни уменьшение размера полимера, наблюдаемые в плодах AC, не наблюдаются в плодах -PG-Exp.
Связанный со созреванием сдвиг полимеров, богатых рамнозой (Rha) и арабинозой (Ara), во фракцию водорастворимой стенки, наблюдаемый в созревающих плодах AC, был почти устранен в трансгенных плодах -PG-Exp (Таблица 1). Это предполагает, что комбинированное действие PG и Exp увеличивает доступность CWDE других фруктов для их локализованных на стенках субстратов, предположительно включая рамноглалактуронан-I (RG-I) и связанные с ним боковые цепи арабинана.Интересно, что в то время как RG-I является частью сети пектинов CW, магистраль RG-I не должна быть целью ни PG, ни Exp.
Таблица 1.
Нейтральный сахарный состав (мол.%) WSF из плодов AC и −PG − Exp на четырех стадиях созревания
Совместное действие PG и Exp влияет на расслабление CW.
Плоды, которые значительно размягчаются на стадии полного созревания, обычно имеют КС, которые кажутся набухшими, как in planta , так и in vitro , и это связанное со созреванием набухание стенок коррелирует с солюбилизацией пектина (27, 28).КС выделяли из LR и RR AC и трансгенных плодов и суспендировали в воде. Изолированные CW демонстрируют, что уменьшение PG или Exp и, в частности, уменьшение PG и Exp приводит к тому, что стенки набухают намного меньше, чем стенки плодов AC (рис. 7). A ) и лишь немного больше стенок из фруктов MG AC (данные не показаны). Кроме того, при исследовании с помощью электронной микроскопии КС плодов RR -PG-Exp кажутся тоньше, чем стенки плодов RR AC (рис. B и C )
Инжир.7.CW набухает по мере созревания плодов томатов AC, но не по мере созревания плодов −PG − Exp. ( A ) Электронные микрофотографии показывают увеличенную толщину стенок (CW) клеток околоплодника плода RR и увеличенное расстояние между электронно-плотным фибриллярным материалом стенки в AC по сравнению с плодом -PG-Exp. (Масштабные полосы, 1 мкм.) ( B ) Толщина CW из 30 изображений, как в A , показывает, что стенки плодов RR AC (черная полоса) толще, чем стенки из RR −PG − Exp (белая полоса) фрукты.Буквы указывают на существенные различия между генотипами; P <0,01. ( C ) Путем сравнения глубины осажденных водных суспензий AIR из LR и RR AC фруктов наблюдается типичное набухание AIR, связанное со созреванием. Такого изменения не наблюдается, когда сэмплам LR и RR AIR из -PG-Exp разрешено оседать. (Шкала 1 см.)
Обсуждение
Одновременное подавление экспрессии LePG и LeExp1 , связанной со созреванием плодов томата, снижает восприимчивость к B.cinerea во время созревания, тогда как подавление LePG или LeExp1 само по себе не снижает восприимчивость, что указывает на то, что PG и Exp действуют совместно, поддерживая как размягчение (4, 22, 23, 29), так и полную патогенность B. cinerea . КС хозяина является первичной мишенью во время роста B. cinerea на растительной ткани (30). B. cinerea обладает широким спектром CWDE (31, 32), включая шесть PG (33). B. cinerea мутанты либо BcPG1 , либо BcPG2 приводили к значительному снижению вирулентности для нескольких хозяев, включая томат (5, 8).Кроме того, эктопическая экспрессия мощного PG-ингибирующего белка (PGIP) из плодов груши ( pPGIP ) снижает восприимчивость спелых плодов томатов к инфекции B. cinerea (6). Таким образом, способность патогена эффективно разбирать CW плодов томата, по-видимому, имеет решающее значение для полной вирулентности. Хотя патоген, по-видимому, обладает необходимыми инструментами для проведения целенаправленного разложения компонентов ХС растений, мы показали, что как его рост, так и развитие симптомов заболевания на плодах томата зависят от двух эндогенных белков ХС растений, которые являются ответственными, вместе с другими фруктами белки для некоторых аспектов метаболизма CW, связанного со созреванием.В других патосистемах для восприимчивости Arabidopsis к Erysiphe cichoracearum и B. cinerea , по-видимому, необходимы растительная активность пектатлиазы CWDE и полная пектинметилэстераза (34, 35).
Совместная эволюция плодов и их патогенов может быть причиной взаимосвязи между полным размягчением и восприимчивостью спелых фруктов к патогенам. LeExp1 и LePG не экспрессируются в плодах до начала созревания, перехода в развитии, который у томатов знаменует завершение созревания семян.Когда семена созревают, плод становится средством распространения семян, процессу, которому может способствовать размягчение, связанное со созреванием, и чувствительность к патогенам, связанная с созреванием. Зависимость патогена от саморазборки CW плодом гарантирует, что семена созревают до того, как разложение плодов инициируется CWDE патогенными микроорганизмами. Таким образом, активность CWDEs в отношении грибковой вирулентности и CWDE для созревания плодов в совокупности способствует завершению жизненных циклов как растений, так и патогенов.
Считается, чтоPG и Exp нацелены на структурно независимые полимерные системы CW; я.э., пектиновые сети стенки и сети Cel-Hem (13, 36). Смягчение фруктов требует комбинированных действий PG и Exp для разрыхления ткани стены, процесса, который, вероятно, отражает изменения во внутри- и интерполимерном связывании, которые делают сети более слабыми и менее прочно связанными друг с другом. Это может способствовать действию других CWDE, связанных со созреванием, о чем свидетельствует сниженная солюбилизация полимеров, богатых Ara- и Rha, в спелых плодах -PG-Exp. Наблюдаемые совместные эффекты PG и Exp на набухание и толщину CW также предполагают, что целостность ткани стенки зависит от состава и взаимодействия двух основных структурных сетей CW.Такое взаимодействие, в свою очередь, может изменить доступность субстрата как для фруктов, так и для патогенных CWDE. Снижение роста грибов в плодах при подавлении LePG и LeExp1 может быть прямым результатом этих изменений в доступности субстрата для патогенных CWDE. Однако есть несколько других возможных механизмов, которые могут способствовать PG- и Exp-зависимым изменениям восприимчивости к патогенам. Например, изменения в структуре CW плодов в результате снижения количества PG и Exp могут служить для удержания ингибиторов функции патогенов, таких как PGIP, потеря ассоциации CW которых во время созревания коррелирует с повышенной восприимчивостью к патогенам (37–39).Снижение PG и Exp также может изменять вызванные патогенами ответы, механизм, предполагаемый демонстрацией того, что изменения CW могут активировать новые пути защиты (40). Эти или подобные механизмы могут работать сами по себе или в сочетании со структурными изменениями CW, которые снижают питательные и / или сигнальные свойства продуктов разборки CW, которые обычно присутствуют в созревающих фруктах (41). Потребуются дальнейшие исследования, чтобы различать эти множественные и потенциально перекрывающиеся механизмы PG- и Exp-опосредованного увеличения восприимчивости к патогенам, которые сопровождают созревание плодов.
Теперь становится очевидным, что одновременное подавление LePG и LeExp1 в созревающих плодах одновременно снижает разборку стенок, замедляет размягчение плодов in vivo и задерживает или снижает связанную со созреванием восприимчивость к B. cinerea . Признание роли связанного со созреванием метаболизма ХС в восприимчивости к патогенам после уборки урожая может быть полезным при разработке и разработке стратегий по ограничению потерь патогенов во время хранения, обработки и распределения фруктов.
Материалы и методы
Растительный материал.
Трансгенные и контрольные линии томатов ( S , lycopersicum ) относились к сорту Ailsa Craig (AC) и выращивались на полях в Дэвисе, Калифорния. Трансгенная линия с уменьшенным PG (-PG) представляла собой антисмысловую линию LePG Gr105, предоставленную Д. Гриерсоном (Университет Ноттингема, Ноттингем, Великобритания) (24), а трансгенная линия с уменьшенным Exp (-Exp) содержала смысловую -подавляющая конструкция LeExp1 , строки 18-40, как описано в ссылке.23. Трансгенные линии с подавленной экспрессией как LePG , так и LeExp1 были получены путем скрещивания гомозиготных линий -PG и -Exp, как описано (22), и гомозиготное потомство подтверждено саузерн-блоттингом и анализом ПЦР (22). . Плоды метили в поле через 3–4 дня после цветения и собирали как плоды MG через 30–31 день, LR через 33–34 дня и RR через 35–36 дней после цветения. Стадии созревания подтверждены визуальным анализом внутренней и внешней окраски плодов.
Измерение плотности плода.
Плотность плода измеряли путем сжатия экватора на 2 мм с помощью плоского зонда из нержавеющей стали диаметром 2 мм (скорость испытания 0,5 мм / с) (TaXT2i Texture Analyzer; Texture Technologies). Фрукты измеряли на трех участках, и наблюдаемая плотность была сопоставима с результатами, приведенными в исх. 22.
Инокуляция
B. cinerea .Б.cinerea (Del 11) выращивали на 1% картофельном агаре с декстрозой, конидии собирали из спорулирующих колоний и фильтровали через стекловату. Фрукты стерилизовали 10% отбеливателем в течение 10 мин с последующим ополаскиванием деионизированной водой. Плоды, пораненные в четырех местах на глубину 2 мм, инокулировали 10 мкл, содержащим 5000 конидий B. cinerea , или 10 мкл воды. Плоды инкубировали при 20 ° C и высокой влажности, и восприимчивость определяли как заболеваемость (процент плодов, проявляющих симптомы мягкой гнили, распространяющиеся на любом из участков инокуляции).Развитие биомассы грибов оценивали с помощью набора QuickStix для B. cinerea (EnviroLogix), в котором используется моноклональное антитело BC12.CA4 (25). Тест является количественным в диапазоне интенсивности сигнала (SI) от 1 до ≈48 на основе экстрактов ткани томата с добавлением сухого мицелия B. cinerea из культур ( r 2 = 0,94; результаты не показаны). Ткань плода (0,3 г) собирали в разное время, объединяли (девять поражений на пул, три пула на линию), суспендировали и измельчали в буфере набора (1:10, вес / объем).Материал с перекрестной реакцией антител измеряли в 500 мкл тканевой суспензии, а интенсивность тестовой линии определяли с помощью считывающего устройства QuickStix (Envirologix).
Изоляция CW.
CW подготовка в соответствии с протоколами исх. 42 для получения сухого нерастворимого в спирте остатка (AIR). Использовали приблизительно 15 г ткани плодов томатов (экзокарпий и околоплодник) для каждой стадии созревания плодов и Botrytis — и имитацию инокулированных обработок.
In vitro Культура B. cinerea на изолированном материале CW.B. cinerea (три споры · мкл -1 ) выращивали в 10 мл среды Пратта, pH 4,5 (43), содержащей 10 мг AIR из фруктов LR и RR в качестве единственного источника органического питания. Было проведено два эксперимента с тремя независимыми повторами на каждый генотип и на dpi. При 1, 3 и 6 dpi культуры центрифугировали при 11000 × g в течение 5 минут и собирали 5 мл супернатанта.Осадки гомогенизировали в оставшихся 5 мл среды с использованием гомогенизатора Ultraturrax T25 Basic при 17000 × г в течение 1 мин. Гомогенат центрифугировали при 11000 × g в течение 5 минут, и этот супернатант объединяли с первым супернатантом для оценки биомассы грибов с помощью набора QuickStix, как описано выше, после разбавления 1: 5 мас. / Об. Собранного супернатанта в комплект буфера.
CW солюбилизация, измерения UA и NS.
Три мг AIR солюбилизировали с H 2 SO 4 (42) и солюбилизированным материалом CW (фрукты или фрукты плюс B.cinerea wall) был проанализирован колориметрически на общий UA с использованием m -фенилфенола (44) и NS с использованием антрона (45).
CW Фракционирование.
Фракции, обогащенные CW пектиновыми и гемицеллюлозными полимерами, последовательно химически экстрагировали из AIR. От ≈200 мг AIR, водорастворимый (WSF), ионно связанный (т.е. растворимый в хелаторе транс -1,2-диаминоциклогексан- N , N , N ‘, N’ -тетрауксусная кислота, CSF) и ковалентно связанные (т.е.е., растворимые в 50 мМ Na 2 CO 3 , NSF) пектины и гемицеллюлоза (т.е. растворимые в 4 M KOH, KSF) были разделены (42). Два независимых образца были извлечены из каждой стадии развития и каждого генотипа. Образцы анализировали в трех повторностях на NS и UA (44, 45).
Эксклюзивная хроматография (SEC).
Аликвоты WSF, CSF, NSF и KSF диализовали (Spectrapor, отсечка молекулярной массы 8 кДа) против дистиллированной воды в течение 1 дня при 4 ° C и лиофилизировали.Образцы WSF, CSF и NSF растворяли в 200 мМ ацетате аммония, pH 5,0, и анализировали с помощью SEC на колонке HW65 (Tosoh Bioscience). Образцы KSF растворяли в 0,1 М NaOH и фракционировали на колонке с сефарозой CL-4B (1,0 × 90 см) (Amersham Pharmacia), элюируя 0,1 М NaOH (42).
Анализ ГХ-МС.
Для анализа NS-сахарных композиций AIR или изолированных полимеров стенок, полученных из здоровой и инокулированной Botrytis ткани, 2 мг AIR или лиофилизированных образцов WSF, CSF, NSF и KSF гидролизовали в 2 M трифторуксусной кислоте ( 46) и преобразовали в ацетаты альдита (47) для анализа ГХ-МС (42).
Просвечивающая электронная микроскопия.
Околоплодник, удаленный из плода RR, фиксировали в фиксаторе Карновского с помощью вакуумно-микроволновой печи (48). Образцы промывали в 0,1 М натрий-фосфатном буфере, pH 7,2, подвергали микроволнам под вакуумом при 450 Вт в течение 40 секунд, затем фиксировали в течение 2 часов в 1% OsO 4 , забуференном 0,1 М натрий-фосфатным буфером, и второй раз подвергали микроволновой обработке при 450 Вт в течение 40 секунд. Через 30 минут в 0,1% дубильной кислоте при 0 ° C, а затем в 2% водном растворе уранилацетата в течение 1 часа образцы дегидратировали в ацетоне и заливали смолой Epon / Araldite.Ультратонкие срезы исследовали с помощью просвечивающего электронного микроскопа Philips CM120 Biotwin Lens (FEI).
CW Набухание
in vitro .Десять миллиграммов ВОЗДУХА суспендировали в 8 мл воды и встряхивали горизонтально в течение 24 часов. Затем пробирки помещали вертикально и оценивали набухание CW на основании высоты осажденного слоя AIR.
Статистический анализ.
Различия между группами исследовали с использованием ANOVA с последующим апостериорным тестированием (действительно значимое различие Тьюки, Tukey HSD) с использованием SYSTAT 11.0 (Systat). Для процентных значений статистический анализ проводился после углового преобразования.
Благодарности
Мы благодарим лабораторию электронной микроскопии Медицинского факультета Калифорнийского университета в Дэвисе, а также Цоки Чора, Уильяма Синко и Хайди Шер за помощь. Эта работа была поддержана грантом Национального научного фонда IOB 0544504, Комиссией Фулбрайта и Советом научных исследований (CONICET) Аргентины.
Сноски
- ¶ Кому следует направлять корреспонденцию. Электронная почта: alpowell {at} ucdavis.edu
Вклад авторов: D.C. и A.R.V. внес равный вклад в эту работу; D.C., A.R.V., F.M.D., A.B.B., J.M.L. и A.L.T.P. спланированное исследование; D.C., A.R.V., L.C.G. и A.L.T.P. проведенное исследование; D.C., A.R.V., L.C.G. и F.M.D. внесены новые реагенты / аналитические инструменты; Д.C., A.R.V., L.C.G., F.M.D., A.B.B., J.M.L. и A.L.T.P. проанализированные данные; и D.C., A.R.V., A.B.B., J.M.L. и A.L.T.P. написал газету.
Авторы заявляют об отсутствии конфликта интересов.
Эта статья представляет собой прямое представление PNAS. H.K. является приглашенным редактором по приглашению редакционной коллегии.
Эта статья содержит вспомогательную информацию на сайте www.pnas.org/cgi/content/full/0709813105/DC1.
- © 2008 Национальная академия наук США
Количественный анализ белков Р тельца дрожжей показывает принципы состава и специфичности
[Примечание редакции: далее следует план авторов по внесению исправлений.]
Основные опасения рецензентов можно резюмировать следующим образом:
1. Новизна работы не подчеркнута. Ясно, что он более всеобъемлющий, чем предыдущие работы этих авторов, но требует более перспективного и дополнительного тестирования.
Что касается новизны, мы считаем, что наша работа добавляет важные новые знания о Р-телах и биомолекулярных конденсатах в целом. Во-первых, наша работа — это единственный отчет об абсолютных или относительных концентрациях основных белков в любом конденсате. Такие концентрации являются важными знаниями при рассмотрении природы и биохимических функций тел фосфора (и, в более широком смысле, других конденсатов). Например, модели конденсатов как отсеков хранения подвергаются сомнению нашими данными на рисунке 3, показывающими, что только небольшая часть большинства белков присутствует в P-тельцах.Кроме того, модели конденсатов как участков высокой биохимической активности должны учитывать тот факт, что большинство неосновных компонентов сконцентрированы только в <10 раз относительно окружающей цитоплазмы и что большинство молекул все еще находится в цитоплазме. Таким образом, конденсатные функции с большей вероятностью возникают из основных компонентов или из совокупности неосновных компонентов (например, в каскаде реакций). Это важные концепции для данной области. Во-вторых, наши данные показывают, что лишь небольшая часть многих белков в P-телескопах имеет высокую концентрацию в структуре.Эта информация является ключевой для понимания строения конденсатов. Это говорит против большинства преобладающих моделей состава конденсата, которые рассматривают отсеки как трудноразрешимо сложные, с сотнями компонентов, все из которых неявно имеют одинаковый вес с точки зрения функциональной значимости. Наши данные показывают, что компартменты намного проще и предоставляют первые пути к надежному биомиметическому восстановлению.
Наконец, хотя нам предстоит еще поработать, чтобы понять корреляции на Рисунке 7, как эмпирический предиктор степени, в которой полноразмерные белки могут быть сконцентрированы в конденсате, они весьма ценны для данной области.
2. Авторам необходимо рассмотреть другие модели для сборки, а не только клиент / скаффолд. Расхождения между данными и их моделью не устранены. Есть несколько подробных экспериментов, которые могли бы сделать больше, чтобы прояснить туманные выводы (см. Ниже). Например, вместо мечения одним флуорофором они могут лучше оценить состав и вариацию с помощью попарного анализа двух флуорофоров.
3. Могут ли они предоставить какие-либо дополнительные данные или идеи, чтобы пролить свет на несоответствие, которое они указывают между их моделью и результатами для DCP2?
Обеспокоенность по поводу модели каркаса / клиента, по-видимому, частично возникает из-за неправильного представления рецензентов о самой модели и о нашем намерении представить корреляции валентностей на рисунке 7.Оглядываясь назад, мы понимаем, как возникли эти неправильные представления, и полагаем, что обширный пересмотр текста в сочетании с дополнительными данными должен привести к гораздо более четкому представлению и более точной модели. Короче говоря, мы не намеревались изобразить модель, в которой валентность взаимодействий является единственным параметром, важным для определения состава конденсата, и где РНК не важна. Скорее, как предполагают составители обзора, мы полагаем, что A) РНК является ключевым каркасом P-телец, B) важно сродство связывания белков друг с другом и с РНК и C) важна валентность взаимодействий.Мы также ошибочно отождествили «основные» белки с «каркасными» белками, а «неосновные» — с «клиентом» в предложении в начале обсуждения, цитируемом вторым рецензентом, вместо того, чтобы описать их как коррелированные, но не идентичные, что было наше намерение до конца обсуждения. Это вызвало ряд проблем, которые можно решить в пересмотренном введении и обсуждении.
Соответственно, мы не предполагали, что данные на Рисунке 7 представляют тест или валидацию основанной на валентности модели каркаса / клиента или результата, вытекающего из предыдущих данных в рукописи.Скорее, мы задумали это просто как эмпирическую корреляцию, которая потенциально может быть полезна при прогнозировании других конденсатов, где количественные данные изображений получить нелегко. Очевидно, это не было правильно, и в нашей презентации модель была основана исключительно на валентности, что не входило в наши намерения. Это можно исправить в доработке. Тем не менее, мы согласны с авторами обзора в том, что для нас важно понять, почему мутанты Dcp2 не попадают в ту же корреляцию, что и различные полноразмерные белки, и разработали дополнительный набор мутантов, чтобы лучше это понять.
Второй рецензент также обращает внимание на то, что стехиометрия ~ 1: 1 многих основных белков предполагает, что они могут формировать стереотипную сборку, которая связывает РНК, а затем рекрутирует другие белки, что приводит к несколько иной модели состава. У нас есть несколько мыслей по этому поводу. Во-первых, мы согласны с тем, что относительная стехиометрия поразительна. Фактически, в более ранней версии рукописи это обсуждалось довольно подробно, как и в комментариях рецензента.Тем не менее, мы в конечном итоге удалили это обсуждение, потому что A) относительная стехиометрия отличается у штаммов дикого типа, лишенных глюкозы, и B) широкий диапазон концентраций P-тел каждого компонента и отсутствие многокомпонентных корреляций в отдельных дрожжевых клетках сделали это сложно претендовать на конкретную сборку. Тем не менее, мы согласны с авторами обзора в том, что наше понимание состава тела P было бы значительно укреплено путем сбора данных с двумя цветами, где мы могли бы количественно определить абсолютные концентрации 2 белков одновременно в отдельных клетках.Хотя непрактично собирать такие данные обо всех возможных парах белков P-тел, мы создадим штаммы для изучения ключевых корреляций внутри основной группы и между стержневыми и неосновными белками. Эти данные позволят нам различать разные модели состава тела Р.
[Примечание редакции: авторы отправили на повторное рассмотрение после принятия решения после экспертной оценки. Далее следует письмо о решении после первого раунда проверки.]
Рецензенты считают, что они не могут поддержать повторное представление этой работы.Причины этого лучше всего резюмирует один из рецензентов, который утверждает:
«Я не сторонник повторного представления. Если я что-то не понял неправильно, авторы не планируют решать проблемы, связанные со значимостью и достоверностью их модели.
Авторы заявляют, что: «Как предполагают обозреватели, мы считаем, что A) РНК является ключевым каркасом для P-телец, B) важно сродство связывания белков друг с другом и с РНК и C) важна валентность взаимодействий» .
Но не предлагаю никаких экспериментов / данных, подтверждающих эти точки.
Мы приносим свои извинения за то, что в нашем первоначальном ответе не было достаточно четко заявлено, что мы намеревались серьезно пересмотреть нашу модель, лучше проиллюстрировать ее значение и проверить ее дальше. Все это мы сделали в ревизии.
Вкратце, здесь мы больше не представляем работу как тестирование модели строительных лесов / клиента для сборки конденсата. Скорее, теперь мы предлагаем гораздо более детальный взгляд на сборку P-телец, подчеркивая роль высокой связности (связанной с валентностью, но отличной от нее) между белками и РНК в производстве конденсата, но также обсуждая важность аффинности связывания и активности. процессы.Мы также обсуждаем несколько других моделей и объясняем, как они несовместимы с данными наших и других лабораторий как в этой статье, так и в литературе. Как было предложено рецензентами и обсуждено с редактором в апреле, теперь мы получили двухцветные данные (изучение корреляции между двумя компонентами Р-тельцов) и данные, исследующие роль высокоаффинных связывающих элементов в управлении рекрутированием Р-тельцов, которые оба оказались полезными при сравнении разных моделей.
Наконец, теперь мы гораздо более подробно описываем новизну наших открытий, которые включают, среди 5 пунктов, подробно описанных в сопроводительном документе: A) открытие, что, несмотря на большое количество молекул, присутствующих в P-телах, только небольшое количество (7) сильно сконцентрированы там с большими коэффициентами разделения и B) открытие, что межмолекулярная связь играет ключевую роль в управлении концентрациями молекул в P-телах.
Они также заявляют, что: «Мы не намеревались, чтобы данные на Рисунке 7 представляли тест или валидацию основанной на валентности модели каркаса / клиента или результата, вытекающего из предыдущих данных в рукописи. Скорее, мы намеревались это просто как эмпирическая корреляция, которая потенциально может быть полезна при прогнозировании других конденсатов, когда количественные данные визуализации получить нелегко ».
Это, похоже, не соответствует названию статьи, обещающей «композиционные принципы».Их предложение изобразить два компонента одновременно, не добавит ничего, кроме подтверждения того, что они уже показали?
Мы удалили количественные корреляции между валентностью и физическими характеристиками P-тел (исходный рисунок 7), что было проблематично по ряду причин. Наши двухцветные эксперименты предоставили важные проверки потенциальных моделей образования конденсата. Кроме того, как указано выше, мы значительно пересмотрели нашу модель.
Мы полагаем, что в рукописи изложены несколько принципов, касающихся состава P-тел.Они описаны в пересмотренном разделе «Обсуждение» статьи, который полностью переписан. К ним относятся: A) концепция (подтвержденная доказательствами) о том, что биологические конденсаты, которые кажутся довольно сложными при качественном анализе их компонентов, на самом деле могут иметь гораздо более простую первичную сложность и организацию при количественном рассмотрении; Б) идея о том, что различия между высококонцентрированными и слабоконцентрированными белками Р-телец, вероятно, тесно связаны с их паттернами межмолекулярной связи внутри Р-телец; C) идея о том, что взаимодействия, которые производят конденсаты, распределяются по их высоко валентным компонентам, они действуют с разной степенью кооперативности, способствуя формированию более крупной сборки; и D) предсказание того, что термодинамика образования конденсата и состав получающейся в результате структуры должны быть связаны, на основе моделей связности в сети взаимодействия; я.е. Удаление сильно связанной молекулы должно влиять как на концентрации других факторов, необходимых для образования конденсата, так и на относительные концентрации компонентов в конденсате.
Наконец, утверждение о том, что «наши данные показывают, что лишь небольшая часть многих белков в Р-тельцах сильно сконцентрирована в структуре. Эта информация является ключевой для концептуализации строения конденсатов. Это говорит против большинства преобладающих моделей состава конденсата, которые Отобразить отсеки как непреодолимо сложные, с сотнями компонентов, все неявно взвешенные в равной степени с точки зрения функциональной значимости.Наши данные предполагают, что компартменты намного проще и предоставляют первые пути к надежному биомиметическому восстановлению «.
Биомиметическое восстановление гранул РНК уже началось, и уже было показано, что нескольких компонентов может быть достаточно для имитации структуры конденсата (Feric et al., 2016, Putnam et al., 2019). Утверждение также, кажется, подразумевает, что знание концентрации каждого фактора поможет идентифицировать такие ключевые компоненты, концепция, которая остается непроверенной ».
Хотя работа, процитированная Feric и Putnam, элегантна, обе используют только два компонента в ядрышках и гранулах P соответственно.Таким образом, степень, в которой они захватывают свойства клеточных структур, неизвестна. Более того, насколько нам известно, не было установлено, что компоненты, используемые в этих реконструкциях, в количественном отношении являются доминирующими компонентами клеточных структур, или что другие компоненты имеют одинаковую концентрацию. Таким образом, опять же, степень, в которой эти воссозданные структуры воспроизводят биологические структуры, является неопределенной. На наш взгляд, важным достижением в биохимическом восстановлении конденсатов, которые очень напоминают клеточные конденсаты, является знание доминирующего (т.е. наиболее концентрированные) компоненты и их динамические свойства in vivo. Обладая этой информацией, можно комбинировать соответствующие молекулы in vitro при их общих клеточных концентрациях и узнать, образуют ли они конденсаты соответствующих концентраций компонентов (как относительных, так и абсолютных) и динамики. Если они соответствуют этим критериям для нескольких компонентов, то можно быть уверенным в том, что биохимия действительно является биомиметической. Без этой предварительной информации человек делает обоснованное предположение о том, какие белки следует комбинировать биохимически и насколько хорошо воссозданная структура имитирует клеточную структуру.
Резюме рецензента: В этой рукописи сообщается о количественном анализе состава тела фосфора в дрожжах. Авторы характеризуют концентрацию, обогащение относительно цитоплазматического пула и динамику 31 зарегистрированного белка P-тельца, меченного GFP. Они идентифицируют 19 белков, которые были достаточно сконцентрированы в P-тельцах в тестируемых условиях, и делят их на два класса: «основная группа», состоящая из 7 белков с высоким обогащением и низкой динамикой, а остальные 12 белков с низким обогащением и высокой динамикой.Авторы утверждают, что эти наблюдения поддерживают модель каркаса / клиента для сборки конденсата, где количество партнеров по взаимодействию для данного белка (валентность) может быть использовано для прогнозирования его обогащения конденсатами / P тел.
Мы тщательно пересмотрели текст и больше не представляем работу как тестирование модели строительных лесов / клиента для сборки конденсата. Фактически, на основе наших данных мы теперь представляем новый, более детальный взгляд на номенклатуру строительных лесов / клиентов, который лучше отражает поведение природных конденсатов и будет более полезным для полевых работ.Мы также удалили анализы, описывающие количественные отношения между обогащением тел P и валентностью взаимодействий. Однако мы утверждаем, что существует значимая качественная взаимосвязь между связностью в сети взаимодействия P-тела и молекулярным поведением, с множеством оговорок, которые теперь подробно описаны в разделе «Обсуждение». Подробнее о каждом из этих вопросов мы расскажем ниже.
Основные опасения рецензентов можно резюмировать следующим образом:
1) Новизна работы не подчеркнута.Ясно, что он более всеобъемлющий, чем предыдущие работы этих авторов, но требует более перспективного и дополнительного тестирования.
Мы тщательно пересмотрели обсуждение, чтобы выделить новые принципы, которые, по нашему мнению, вытекают из наших данных. Во-первых, наша работа — это первый всесторонний количественный анализ состава и динамики любого природного биомолекулярного конденсата. Во-вторых, это количественное определение показало, что, несмотря на большое количество молекул, присутствующих в P-телах, только небольшое количество (7) сильно сконцентрировано там с большими коэффициентами разделения.Это большое упрощение при рассмотрении формирования, регуляции и функции Р-телец. В-третьих, количественный анализ показал, что только небольшая часть большинства белков присутствует в P-тельцах, что имеет важное значение для функций конденсатов. В-четвертых, большинство (6 из 7) высококонцентрированных белков / комплексов сильно связаны в сети взаимодействия P-тельцов, и ни один из слабо концентрированных белков сильно связан. Эти корреляции предполагают, что связность играет важную роль в управлении составом структур.Однако важно отметить, что другие факторы, включая сродство связывания и активные процессы, также вносят вклад в молекулярное поведение, и теперь мы объясним такие сложности в нашем обсуждении. Наконец, основываясь на наших данных и других данных в литературе, мы пересмотрели понятие каркасов и клиентов в биомолекулярных конденсатах. Мы утверждаем, что эти термины не следует использовать для классификации молекул на бинарные группы, как мы делали раньше, а скорее как качественные дескрипторы степени, в которой молекула способствует образованию и составу конденсата.Таким образом, молекула должна быть описана как более подобная каркасу или более подобная клиенту, в зависимости от того, имеет ли она больший или меньший эффект, соответственно. Такое использование терминов лучше отражает экспериментальные данные, но при этом сохраняет идею о том, что одни молекулы вносят больший вклад, чем другие, в образование и состав конденсата.
2) Авторам необходимо рассмотреть другие модели для сборки, а не только клиент / скаффолд. Расхождения между данными и их моделью не устранены.Есть несколько подробных экспериментов, которые могли бы сделать больше, чтобы прояснить туманные выводы (см. Ниже). Например, вместо мечения одним флуорофором они могут лучше оценить состав и вариацию с помощью попарного анализа двух флуорофоров.
Теперь мы даем гораздо более детальное представление о сборке P телец, подчеркивая роль сильно связанных молекул (как белков, так и РНК) в производстве конденсата, но также обсуждая важность сродства связывания и других параметров.По просьбе обозревателей мы также выполнили двухцветную визуализацию, чтобы изучить корреляцию между обогащением между различными молекулярными парами (новый рисунок 3). Эти данные выявили значительную корреляцию (коэффициент Пирсона 0,6-0,7) между обогащением Dcp2 и Edc3, Pat1 и Xrn1. Корреляция Dcp2-Xrn1 особенно интересна, поскольку неизвестно, как эти два белка связываются друг с другом напрямую, что указывает на корреляции, опосредованные косвенной связностью в конденсате (вероятно, через РНК или другие компоненты ядра P-тела).Однако корреляции не столь сильны, чтобы предполагать, что стехиометрически определенный комплекс лежит в основе образования Р-телец (модель, предложенная одним из рецензентов). Более того, как мы приводим ниже, существующие данные говорят против модели, в которой РНК является единственной подобной каркасу молекулой в P-тельцах (вторая модель, предложенная в обзоре), так как ряд белков, как было показано генетически, играют важную роль в образование конденсата. Эти вопросы — кооперативность в рекрутинге, возможность стехиометрического комплекса, роли взаимодействий РНК-РНК и РНК-белок и другие факторы — теперь довольно подробно рассматриваются в разделе «Обсуждение» рукописи.Мы надеемся, что этот более подробный взгляд на образование конденсата понравится рецензентам.
3) Могут ли они предоставить какие-либо дополнительные данные или идеи, чтобы пролить свет на несоответствие, которое они указывают между их моделью и результатами для DCP2?
Мы решили эту проблему двумя способами. Во-первых, мы создали дополнительные мутанты Dcp2, чтобы лучше понять, как его связывание с Edc3 контролирует его обогащение P-тельцами (новый рисунок 6). Это приводит к важному выводу, что когда сродство между двумя конденсатными белками низкое, увеличение сродства может увеличить обогащение, но когда сродство уже высокое, его дальнейшее увеличение не увеличивает обогащение.Таким образом, настройка обогащения, вероятно, происходит за счет изменений в режиме низкого / умеренного сродства. Кроме того, как описано выше, новое обсуждение дает более детальное представление о найме, поскольку происходит из комбинации возможности подключения, аффинности и активных процессов. Вместе они объясняют поведение наших мутантов Dcp2. По сути, не все регионы Dcp2 в равной степени способствуют обогащению; удаление элемента связывания с высоким сродством (либо к РНК, либо к Edc3) имеет гораздо более выраженный эффект на обогащение, чем удаление элемента с низким сродством.
Рецензент № 1:
[…] Для дальнейшего улучшения новинки предлагаем дополнительные вопросы, на которые можно было бы ответить:
1) Основным достижением этого исследования является точное измерение концентрации GFP-слитых белков, которое выполняется путем сравнения интенсивности клеточного GFP со стандартными кривыми. Все выводы зависят от этих стандартов и вычитания фона. Несмотря на то, что лаборатория Розена ранее опубликовала статью, в которой использовался аналогичный подход (Banani et al., 2016), подробности этих калибровок и вычитания фона следует указывать в дополнительных материалах.
Мы добавили подробное описание получения изображения (показано на рисунке 1 — рисунок в приложении 2) и процедур анализа в раздел «Материалы и методы».
2) Чтобы оценить концентрацию белка в Р-тельцах, авторы сделали фундаментальное предположение: все Р-тельца имеют одинаковый и постоянный состав. Однако на Рисунке 1 авторы показали, что основные компоненты Р-телец, которые также являются наиболее распространенными белками, имеют сильную (4-5-кратную) изменчивость по концентрации и коэффициенту распределения.Неясно, как эта изменчивость влияет на относительные количественные характеристики, представленные в этой рукописи (рис. 3). Контрольный эксперимент, который мог бы ответить на этот вопрос, заключался бы в том, чтобы пометить несколько компонентов Р-телец спектрально различными флуоресцентными белками (2 или 3 одновременно) и убедиться, что ожидаемые относительные соотношения повторяются.
Отметим, что при измерении абсолютных концентраций различных видов мы не делали никаких предположений о том, имеют ли P-тела постоянный или переменный состав.Предполагая, что GFP-тегирование не изменяет концентрации данного белка (что подтверждают наши данные на рисунке 1 — рисунок в приложении 3), наши данные охватывают диапазон значений концентрации, взятых для каждого компонента в популяции клеток. Эти данные показывают, что составы имеют существенную изменчивость, как отмечает рецензент.
При интерпретации данных становится важным вопрос относительной изменчивости. Здесь мы благодарим рецензента за предложение многоцветных экспериментов. На новом рисунке 3 мы использовали двухцветную визуализацию для одновременной количественной оценки концентраций трех пар белков в отдельных клетках: Dcp2 / Edc3, Dcp2 / Pat1 и Dcp2 / Xrn1.Мы обнаружили, что во всех случаях концентрации тел P в парах значительно коррелируют (R Пирсона 0,6–0,7). Таким образом, концентрации этих компонентов колеблются вместе (хотя и с некоторой остающейся вариабельностью, поскольку значения R не равны 1). Учитывая высокую взаимосвязь взаимодействий между белками Р-тельцов, мы полагаем, что многие компоненты, вероятно, демонстрируют сходные корреляции, но более обширный анализ этого момента выходит за рамки этого и без того длительного исследования.
Могут ли быть в популяции клеток несколько типов P-телец с разными композиционными профилями — это сложный вопрос.Мы не видим убедительных доказательств мультимодального распределения в профилях концентрации, что говорит против такой возможности. Но, опять же, для всестороннего рассмотрения этой возможности потребуется гораздо больше данных, чтобы оценить, может ли распределение концентраций лучше соответствовать одной или нескольким популяциям.
3) В том же направлении неясно, экспрессировались ли белки, меченные GFP, которые использовались для количественной оценки компонентов Р-телец в фоне dcp1∆, из эндогенного локуса (Таблица 1: YRP1936, yRP2254, yRP2237, yRP2246, yRP2230, yRP1840, yRP1736, yRP2269, yRP1844, yRP1916, yRP1842).Авторы должны указать это, так как это будет иметь значительные последствия для количественной оценки, если будет смешанная популяция меченых и немаркированных белков.
Все меченные GFP и mCherry белки, использованные для создания рисунков 1-4, экспрессировались из своего эндогенного локуса. Об этом говорится в основном тексте и в Материалах и методах.
4) На рисунке 2 авторы обнаружили отрицательную корреляцию между скоростью восстановления фракции и концентрацией P-тел.Однако молекулярный механизм этой корреляции не обсуждается. Авторам следует расширить обсуждение и связать его с валентностью взаимодействия.
Как было предложено рецензентом, в Обсуждении мы теперь связываем связность в сети (количество различных молекулярных типов, с которыми взаимодействует данный белок, что связано с валентностью взаимодействия, количество различных молекул, с которыми взаимодействует данная молекула) с Поведение FRAP. Мы заявляем: «За исключением Xrn1 (см. Ниже), все основные белки обладают высокой валентностью взаимодействия (количество взаимодействующих молекул) и высокой связностью с другими белками Р-тельцов и РНК (≥ 4 непосредственно взаимодействующих молекул).[…] Эти особенности обычно должны обеспечивать более низкую концентрацию тел P и более быстрый и полный обмен ».
5) На рисунках 4 и 5 авторы измерили коэффициенты разделения нескольких мутантов Dcp2. Было бы важно продемонстрировать, что эти мутации существенно не влияют на стабильность и экспрессию белка, с помощью вестерн-блоттинга этих мутантов.
На рис. 5 — приложение к рисунку 1B и 1C и на рисунке 7 — приложение к рисунку 1A и 1B, теперь мы показываем вестерн-блоттинг почти всех мутантов Dcp2, исследованных под микроскопом.Эти данные показывают, что белки, сравниваемые на одном рисунке, экспрессируются примерно на одном уровне. Мутанты, не исследованные с помощью вестерн-блоттинга, были исследованы с помощью флуоресцентной визуализации для сравнения общего уровня экспрессии в проанализированных клетках (Рисунок 6 — рисунок в приложении 1). Мутанты Dcp2, сравниваемые на рисунке 6, действительно экспрессируются на несколько разных уровнях, например, Dcp2C∆5H, который слабо делится на P-тельца, экспрессируется на ~ 30% выше, чем Dcp2C∆5H-h2, который делит намного сильнее (рисунок 6 и рисунок 6). — приложение к рисунку 1).Однако направление этой разницы фактически усиливает наш вывод о том, что нижнее разделение Dcp2C∆5H происходит не из-за пониженной экспрессии.
6) На рисунке 5 авторы предполагают, что декапирующая активность Dcp2 влияет на коэффициент разделения и скорость восстановления. Однако, как авторы упоминают в тексте, неясно, связано ли это с увеличением цитоплазматической мРНК или прямым результатом каталитической активности Dcp2. Эта проблема может быть решена путем экспрессии вариантов Dcp2 в клетках дикого типа и индукции Р-телец с голоданием.
Благодарим рецензента за это предложение. Мы выполнили этот эксперимент, и результаты представлены на новом рисунке 7. Вкратце, в условиях голодания динамика и распределение количества Р-телец одинаковы для двух мутантов, что позволяет предположить, что эти параметры в основном отвечают на мРНК. уровни при нарушении катализа. Однако общая доля материала в P-тельцах у каталитического мутанта во время голодания все же выше. Таким образом, этот параметр P-телец, по-видимому, является ответом на потерю каталитической активности.На основании этих данных мы заключаем, что как увеличение уровней мРНК, так и потеря каталитической активности объясняют поведение мутанта WD.
7) На рис. 7 авторы обнаружили корреляцию между разделением Р-телец и валентностью взаимодействия белков. Это открытие было бы более убедительным, если бы авторы смогли продемонстрировать, что когда валентность взаимодействия белка увеличивается, этот белок становится более концентрированным в Р-телец. Например, это можно сделать путем слияния клиентского белка с доменом каркасного белка, такого как Dcp2C ∆5H.Слитый белок должен обладать большей валентностью взаимодействия, и можно ожидать, что слитый белок будет больше концентрироваться в Р-тельцах. Этот эксперимент также усилит вывод, сделанный на Рисунке 6.
Мы удалили рисунок 7 из статьи и больше не обсуждаем количественные отношения между валентностью и разделением. Проблема с вызовом одной только валентности состоит в том, что она игнорирует сродство (проблема, о которой мы упоминали в предыдущем тексте, но не особо подчеркивали), а сродство играет важную роль в разделении Dcp2.В исправленной рукописи мы рассмотрели это двумя способами. Во-первых, мы прямо заявляем, что данные здесь и в литературе показывают, что связывание РНК N-концевым доменом и связывание Edc3 с помощью HLM1 происходит с высокой аффинностью и сильно способствует разделению Dcp2, в то время как C-концевые элементы HLM имеют меньшее сродство и вносят меньший вклад (рис. 5). Во-вторых, мы провели эксперименты, аналогичные предложенным рецензентом. Мы добавили высокоаффинный Edc3-связывающий мотив, HLM1, как к Dcp2 ∆5H, который слабо разделяет, так и к Dcp2 дикого типа, который сильно разделяет.Мы обнаружили, что добавление HLM1 только увеличивает разделение первого белка (рис. 6D). Из этих данных мы заключаем, что когда сродство к Edc3 низкое (Dcp2 ∆5H), добавление дополнительного сайта связывания с высоким сродством может увеличивать разделение Dcp2. Но когда аффинность уже высока (дикий тип Dcp2), дальнейшее ее увеличение не имеет никакого эффекта. Этот результат, вероятно, является общим при рассмотрении рекрутирования белков в конденсаты.
Рецензент № 2:
[…]
Эта модель была получена группой Розена в предыдущем исследовании, в котором изучались свойства разделения фаз in vitro искусственного набора белков с различным количеством низкоаффинных связывающих мотивов (валентность).Модель предсказывает, что «каркасы» (белки, необходимые для сборки конденсата) демонстрируют высокое обогащение и высокую валентность, тогда как «клиенты» демонстрируют низкое обогащение и низкую валентность. В соответствии с моделью авторы отмечают грубую корреляцию между валентностью и обогащением среди исследованных белков тела 19 P (но см. Ниже). Чтобы проверить модель напрямую, они удалили связывающие мотивы в одном предсказанном «высокомалентном» протеине Р-тельца (Dcp2). Удивительно, но они не обнаружили сильной корреляции между валентностью и обогащением (многие мутанты Dcp2 не лежат в корреляциях на Рисунке 7.Нам непонятно, как разрешить это несоответствие). Они также обнаружили, что некоторые известные клиенты (белки, не важные для сборки P-телец) демонстрируют сильное обогащение, вопреки модели. Несмотря на эти несоответствия, авторы продолжают утверждать, что их результаты подтверждают модель каркаса / клиента.
Мы тщательно пересмотрели текст и наш анализ наших данных, чтобы отреагировать на эту критику. Наиболее важно то, что мы удалили рисунок 7 и устранили все обсуждения, связанные с количественной корреляцией между валентностью и концентрацией P-тел.Как утверждает рецензент, одна только валентность не может объяснить наши данные о мутантах Dcp2. Теперь мы представляем гораздо более детальное представление о том, как, по-видимому, определяются концентрация и динамические свойства компонентов P-тела. Мы считаем, что связность взаимодействий (количество типов молекул, с которыми контактирует конкретная молекула) играет важную роль, поскольку большинство (6 из 7) высококонцентрированных белков также сильно связаны (≥4 партнеров по взаимодействию), и все слабоконцентрированные белки имеют низкую связность (≤2 партнеров по взаимодействию).Кроме того, было показано, что большинство (те же 6 из 7) сильно связанных молекул вносят вклад в сборку тел P, и ни одна из молекул с низкой связностью не делает этого. Тем не менее, связность — не единственный фактор, и сродство связывания и активные процессы также могут быть очень важны. В этом свете мы теперь обсуждаем, как взаимодействия N-концевого домена Dcp2 с РНК и элемента HLM1 с Edc3 имеют более высокое сродство, чем другие взаимодействия Dcp2, и, таким образом, играют более важную роль в определении концентрации белка в P-тельцах. (объясняя данные об удалении).Точно так же Xrn1 связывает РНК с высоким сродством. Таким образом, несмотря на то, что его связность низкая, он сильно сконцентрирован в P-телах и демонстрирует умеренную динамику. Что касается активных процессов, гидролиз АТФ с помощью Dcp2 также вносит вклад в динамику белка. Таким образом, связь важна, но не вся история в определении поведения молекул конденсата. Эти вопросы обсуждаются в новом Обсуждении.
Благодаря этим соображениям мы пришли к другому взгляду на то, как следует использовать понятия «эшафот» и «клиент».Вместо того, чтобы классифицировать молекулы как каркас или клиент, мы теперь считаем, что эти термины следует использовать как дескрипторы. Молекула более подобна каркасу, если она играет большую роль в сборке Р-телец, и более подобна клиенту, если играет меньшую роль. Таким образом, мы можем объяснить тот факт, что разные молекулы по-разному влияют на делецию. Например. РНК, по-видимому, является наиболее подобным каркасу компонентом Р-телец, поскольку ее удаление РНКазой разрушает конденсаты (что согласуется с тем фактом, что все 19 компонентов Р-телец, количественно определяемые здесь, связывают РНК), в то время как Edc3 и Pat1 менее подобны каркасу, поскольку их делеционные штаммы сохраняют некоторые Р-тельца, хотя и меньшего размера, чем штаммы дикого типа.Напротив, все 12 изученных нами неосновных белков подобны клиенту, поскольку ни один из них не продемонстрировал значительного уменьшения сборки P-телец при удалении. Мы отмечаем, что связность (и, вероятно, центральность сети), по-видимому, играет роль в определении того, является ли молекула более похожей на каркас или более похожей на клиента, поскольку все наиболее связанные молекулы подобны каркасу (и более центральны в теле P. сеть взаимодействия) и молекулы с небольшим количеством подключений все похожи на клиентов (и более периферийны в сети).Эти вопросы обсуждаются в новом Обсуждении в новом разделе под названием «Общие принципы строительных лесов и клиентов в природных конденсатах».
Очень удивительно, что авторы не рассматривают другие модели, которые могли бы лучше соответствовать их данным. Одна возможность состоит в том, что РНК функционирует как истинный каркас для Р-телец и что белки рекрутируются в Р-тельца в силу их сродства к РНК или другим белкам, которые связывают РНК. В этом отношении интересно, что компоненты мультибелковых комплексов обнаруживаются в примерно стехиометрических количествах в P-тельцах, что позволяет предположить, что комплексы сохраняются в конденсатах.В этой альтернативной модели высокого сродства к РНК одного белка в комплексе было бы достаточно для обогащения всего комплекса, даже если ни одна из других субъединиц в комплексе не связывается с другими компонентами P-тельца.
Насколько мы понимаем этот комментарий, рецензент предполагает, что Р-тельца образуются посредством взаимодействий между мРНК, а затем эти мРНК рекрутируют основные белки Р-тельца за счет связывания РНК с высоким сродством. Однако несколько наблюдений в литературе опровергают эту модель.Например, как мы указываем в рукописи, формирование Р-телец явно требует белок-белковых взаимодействий, поскольку генетически показано, что Dcp2, Edc3, Dhh2, комплекс Lsm1-7 и Pat1 способствуют образованию Р-телец (например, Decker et al. , 2007; Sheth, Parker, 2006; Hondele et al., 2019; Rao, Parker, 2017). Более того, механизмы, с помощью которых белки способствуют образованию P-телец, могут быть напрямую связаны со специфическими белок-белковыми взаимодействиями, такими как димеризация Edc3 (Ling et al., 2008).Кроме того, некоторые белки нуждаются в других компонентах Р-телец для их повторного превращения в Р-тела. Напр., Dcp1 требует, чтобы Dcp2 рекрутировался в P-тела, а комплекс Lsm1-7 требует Pat1 (Teixeira and Parker, 2007). Таким образом, образование Р-телец и привлечение основных компонентов Р-телец к Р-тельцам требует взаимодействия белков.
Мы согласны с рецензентом в том, что РНК способствует образованию Р-телец. Явные данные свидетельствуют о том, что для образования Р-телец необходим пул нетранслирующих мРНК (Teixeira et al., 2005). Одна очевидная роль РНК состоит в обеспечении сайтов связывания для взаимодействующих белков, тем самым обеспечивая сборку Р-телец. В этой роли мы согласны с тем, что РНК функционирует подобно каркасу. Способствует ли РНК также образованию Р-телец посредством межмолекулярных взаимодействий РНК-РНК, еще предстоит установить. Тем не менее, модель, в которой РНК является единственным каркасом, который просто связывает белковые компоненты P-телец, маловероятна. Чтобы прояснить этот вопрос в рукописи, мы добавили этот аргумент в новое Обсуждение.
Более того, как описано выше, теперь мы представляем гораздо более подробный и детализированный вид сборки P-тела, который не зависит от простой бинарной классификации молекул как каркасов, так и клиентов. В пересмотренном Обсуждении также рассматривается вторая модель, предложенная здесь рецензентом, согласно которой высококонцентрированные белки образуют стехиометрический комплекс, который затем собирается на РНК с образованием более крупной структуры. Стехиометрический комплекс показал бы чрезвычайно высокую кооперативность при рекрутировании в P-тела, поскольку все элементы входили бы или выходили вместе.Хотя новый рисунок 3 действительно показывает корреляции между концентрациями Dcp2 в теле P с Edc3, Pat1 и Xrn1, корреляции недостаточно высоки (значения R Пирсона 0,6-0,7), чтобы предполагать стехиометрически определенный набор, даже если белки имеют примерно равные значения. средние концентрации в конденсате. Кроме того, концентрации P-телец Pat1 и комплекса Lsm1-7 снижаются примерно в два раза в штаммах дикого типа в условиях голодания по глюкозе по сравнению со штаммами dcp1Δ , что опять же говорит против дискретной сборки.Эти аргументы представлены в новом абзаце Обсуждения.
Авторы также утверждают, что их анализ показывает, что относительно небольшое количество основных белков (7) составляет большую часть содержания белка в P-тельцах. Однако, поскольку другие компоненты P-тела, возможно, еще не обнаружены, это утверждение может быть только приближенным в настоящее время. Более того, даже если это правда, значение этой гипотезы неясно — белки, присутствующие в низких концентрациях в P-тельцах, могут по-прежнему играть важную роль.
Мы удалили это утверждение из текста.
В заключение, хотя в этом обзоре документируются концентрации и динамика значительного количества белков P-тела, оно не дает существенного понимания «правил», которые регулируют состав P-тела. Модель, предложенная авторами, не согласуется с данными, и другие более правдоподобные модели не рассматриваются.
Как описано в нашем ответе на первый сводный комментарий в верхней части этого обзора, мы чувствуем, что наша работа выявила ряд новых и важных принципов, касающихся формирования и состава P-тел.В новом тексте представлено гораздо более подробное описание сборки P-тела, которое теперь согласуется со всеми нашими данными, и рассматривает множество различных механистических проблем и возможностей.
Рецензент № 3:
[…]
— Предел новизны. Уже было известно, что эти 9 белков являются основными белками P-тельца, которые также играют некоторую роль в функции P-тельца. Эти более ранние исследования также уже показали, что некоторые из этих белков содержат несколько доменов взаимодействия, через которые взаимодействуют компоненты P-тельца.
Как описано в ответе на Сводный пункт 1 в верхней части этого письма, мы считаем, что наша работа является новой во многих отношениях, включая то, что она является первым всеобъемлющим количественным анализом любого биомолекулярного конденсата. С помощью этого количественного анализа мы выявили два различных класса молекул на основе концентрации в структурах, чего нельзя было вывести из предыдущей работы, а также важную идею о том, что почти для всех компонентов только небольшая часть молекул присутствует в P тела.Анализ этих данных в свете известных молекулярных взаимодействий также выявил корреляции между физическим поведением компонентов P-тел и их молекулярной связностью, аффинностью связывания и активностью гидролиза АТФ. Опять же, это не могло быть выведено из предыдущих данных, поскольку они по своей сути зависят от нашего количественного анализа. Если рецензент сможет указать нам на соответствующую литературу, в которой документируется, что основные белки Р-телец, которые мы определяем как высокообогащенные, имеют более высокие коэффициенты разделения, чем другие идентифицированные компоненты Р-телец, мы были бы рады соответствующим образом пересмотреть нашу рукопись.
— Пределы количественного анализа. Конфокальный анализ имеет четкие пределы пространственного разрешения, а количественные измерения основаны на интенсивности флуоресценции, а не на измерениях отдельных молекул. Поскольку все компоненты помечены одним и тем же флуорофором (GFP), нельзя использовать парный анализ или анализ более высокого порядка для определения различий между гранулами, занятости белков, корреляции между белками, чтобы, например, проверить, все ли компоненты находятся в грануле одновременно. , и с аналогичной концентрацией.
Как подробно описано выше, мы выполнили двухцветную визуализацию нескольких пар белков P-телец и обнаружили существенную кооперативность между их обогащением. Это привело нас к более детальному рассмотрению сборки P body, которая описана в практически полностью переписанном разделе «Обсуждение» рукописи.
— Неясная актуальность анализа DCP2. Анализ DCP2, похоже, не сильно улучшает выводы статьи. Вместо того, чтобы показывать, что этот белок поливалентен, как это было показано в прошлом рассечением доменов других «каркасных» белков, было бы информативно, если бы авторы предоставили, действительно ли «клиентские» белки имеют меньше доменов взаимодействия.
Наш анализ Dcp2 проанализировал механизмы, с помощью которых он обогащается внутри P-тел. Хотя понятие многовалентности было продемонстрировано и для др. Белков, это рассмотрение предоставляет полезную информацию о Dcp2. Кроме того, в исследованиях Dcp2 мы обнаружили два ранее неизвестных свойства, которые, вероятно, имеют отношение к компонентам других конденсатов. Во-первых, мы обнаруживаем, что два фрагмента белка, которые только слабо разделяются на P-тельца по отдельности, при слиянии вместе сильно и совместно разделяются.Это происходит, даже если фрагменты связываются с разными компонентами P-тел, и, таким образом, не то же самое, что простая авидность. Когда мы приступили к работе, этот результат был для нас далеко не очевиден. Во-вторых, в новых данных, представленных на новом рисунке 6, мы обнаруживаем, что добавление элемента, связывающего Edc3 с высоким сродством к белку Dcp2, только увеличивает разделение на P тельца, когда Dcp2 имеет низкое сродство к Edc3. Когда сродство уже высокое, добавление дополнительных элементов привязки не имеет никакого эффекта. Таким образом, разделение компонента конденсата может быть насыщенным, вероятно, из-за ограниченных концентраций партнеров по связыванию.Опять же, это не был очевидный результат, когда мы начали эти эксперименты.
Что касается белков с поведением, подобным клиентскому (см. Ответ на пункт 1 сводки для нашего нового способа использования этого термина), как подробно описано выше, мы показываем в таблице 3 и на рисунке 2 — добавлении к рисунку 3, что все молекулы, которые слабо сконцентрированы в P Тельца имеют низкую связь с другими белками Р-телец и, как известно, не вносят значительный вклад в сборку Р-телец. Итак, действительно, поведение, подобное клиенту, коррелирует с ограниченным взаимодействием с другими компонентами тела Р.Корреляция не всегда верна в противоположном направлении. То есть существует небольшое количество белков, которые имеют лишь несколько связей с другими компонентами P-телец, но при этом сильно сконцентрированы в конденсате (Xrn1, Dcp1 и Pby1). Эти белки, как известно, обладают высоким сродством к другому высококонцентрированному компоненту P-тельца, который их рекрутирует. Эти вопросы теперь подробно рассматриваются в нашем обновленном Обсуждении.
[Примечание редакции: до принятия были предложены дальнейшие исправления, как описано ниже.]
Редакций:
Рецензенты сочли, что рукопись и работа вносят вклад в эту область, но необходимы некоторые исправления, чтобы учесть оговорки при интерпретации данных. Например, рецензент 2 считает, что термин «кооперативность» надлежащим образом не подтверждается данными и что мРНК не рассматривается как компонент P-телец.
Мы изменили наше описание данных на рисунке 5F, источника этого утверждения, на «синергетический», в котором говорится, что эффект объединения двух компонентов вместе приводит к большему эффекту, чем сумма компонентов по отдельности.В случае Dcp2 конструкция Dcp2 300Δh2 имеет ПК, равное 2,5, а конструкция Dcp2C Δ5H имеет ПК, равное 3,5, в то время как конструкция Dcp2Δh2 Δ5H, которая объединяет Dcp2 300Δh2 и Dcp2C Δ5H, имеет ПК 31, что> 2,5 *. 3.5. Таким образом, система соответствует определению синергии. Теперь отметим, что этот эффект может быть результатом кооперативного связывания с молекулами в теле P.
В рукописи РНК упоминается как важный компонент P-тельца. Мы считаем, что этого достаточно, особенно с учетом того, что статья сосредоточена на описании белков Р-тел, а не РНК.
Reviewer 3 считает, что функциональные свойства недостаточно изучены, особенно с учетом специфичности (почему исключены некоторые мРНК и RBP?).
Наше исследование не пытается рассмотреть функцию P-тел. Это сложный вопрос, который потребует гораздо больше и других данных, чем мы реально можем поместить в рукопись. Наша работа действительно обращается к специфичности рекрутирования белков в значительной степени посредством идеи синергетического рекрутирования Dcp2 в P тельца.То есть, резидентные белки часто содержат несколько слабых элементов рекрутирования, и, как следствие, белки, которые содержат мало таких элементов, рекрутируются плохо или вообще не рекрутируются. Рекрутирование РНК может следовать схожим принципам, и теперь мы прямо упоминаем эту идею.
Рецензент № 2:
В этой в значительной степени переработанной рукописи авторы сообщают о всесторонней количественной оценке составляющих белков биологического конденсата, обедненных Dcp1 Р-телец дрожжей. Благодаря этому они идентифицировали семь белков «ядра» Р-тельцов, которые существенно разделены, имеют высокую концентрацию и медленную подвижность при обогащении Р-тельцами (рис. 1 и 2).Концентрация этих семи белков в тельцах P положительно коррелирует, что согласуется с предыдущими данными, показывающими, что эти белки взаимодействуют биохимически (рис. 3). Сообщаемые параметры обогащения кажутся похожими, когда обедненные Dcp1 P-тельца сравниваются с P-тельцами дикого типа в условиях глюкозного голодания, однако Dcp1 и его партнер по связыванию Pby1 отсутствуют, и деградация РНК дефектна в модели P-тельцов, используемой на протяжении всего анализа. Таким образом, хотя на уровне их биофизического анализа P-тела, обедненные Dcp1, кажутся похожими на P-тела в физиологических условиях, функционально эти «P-тела» различны.Это снижает актуальность данного исследования.
Рецензент прав в том, что мы изучали Р-тельца в 2 различных условиях, только одно из которых является диким типом. Отметим, что это не полностью отличается от большинства исследований в этой области. Например, большинство анализов стрессовых гранул индуцируют в них мышьяк, токсин, который редко встречается в естественной биологии. Многие другие исследования связаны с избыточной экспрессией отдельных компонентов, что также является нефизиологической ситуацией.Тем не менее, такой анализ имеет существенное значение для понимания принципов, по которым собирается конденсат, и мы считаем, что наше исследование проводится в том же духе. В то время как Dcp1 — / — P тельца и P тельца дикого типа, лишенные глюкозы, могут иметь разные функции, что поразительно, так это то, что их составы тесно связаны. Кроме того, сохраняются ключевые паттерны состава — небольшое количество высококонцентрированных белков и большее количество слабоконцентрированных белков — что является важной особенностью этих клеточных структур, которая, насколько нам известно, не исследовалась где-либо еще (даже 3 недавно опубликованных Cell в статьях о стрессовых гранулах указаны только коэффициенты распределения, в которых отсутствует информация относительно абсолютных концентраций, указанных здесь).
На рис. 4 авторы наблюдают, что концентрации корового белка в Р-теле в значительной степени не зависят от стрессовых условий, однако авторы рассматривали только белковые компоненты, в то время как регулируемым субъектом стрессовых Р-телец являются мРНК, а не белки.
Мы заявляем, что наши данные не касаются изменений в содержании мРНК во время голодания: «… они не говорят о секвестрации / хранении РНК…». Мы также отмечаем, что основной смысл рисунка 4 состоит в том, чтобы показать, что для большинства белков Р тельца существенно не истощают количество молекул из цитоплазмы, не сравнивая Dcp1 — / — и Р тельца дикого типа, лишенные глюкозы.Если обозреватель заинтересован, недавние результаты, полученные на клетках млекопитающих, предполагают, что состав мРНК P-телец, по-видимому, в значительной степени регулируется скоростью их трансляции, возможно, с дополнительным входом от длины поли (A) хвоста (Matheny et al., 2019) .
Затем авторы анализируют локализацию P-тельца DCP2 в двойном нуль-мутанте Dcp1, Dcp2, который все еще образует некоторые P-тельца (хотя на рисунке S1A показано только присутствие небольших структур, подобных «P-тельцам», но без количественного сравнения с контрольными P-тельцами, в дикого типа при голодании по глюкозе и истощении Dcp1).
Мы не совсем уверены, о чем здесь спрашивает рецензент, поскольку мы количественно оценили PC и динамику для различных повторно экспрессируемых мутантов Dcp2. Теперь мы количественно оценили распределение по размеру точки Edc3 в двойном нулевом мутанте Dcp1, Dcp2, показанном на рисунке S5A (рисунок S1A не имеет отношения к этим экспериментам; мы предполагаем опечатку от автора обзора), и сообщаем, что на рисунке 5 — дополнение к рисунку 1А.
В исследованиях делеций авт. Обнаруживают, что различные домены Dcp2 вносят вклад в разделение P тел и демонстрируют некоторую избыточность между доменом h2 и др. Спиральными доменами в CTP.Авторы приходят к выводу, что элементы в Dcp2 действуют «кооперативно». Однако их данные показывают аддитивность или, в лучшем случае, синергию, но не кооперативность. Чтобы использовать термин кооперативность, авторам необходимо продемонстрировать, что начальное событие связывания увеличивает сродство связывания последующих событий связывания.
Как указано выше, мы изменили наше описание данных на рисунке 5F, источника этого утверждения, на «синергетический», в котором говорится, что эффект объединения двух компонентов вместе приводит к большему эффекту, чем сумма компонентов по отдельности.В случае Dcp2 конструкция Dcp2 300Δh2 имеет ПК, равное 2,5, а конструкция Dcp2C Δ5H имеет ПК, равное 3,5, в то время как конструкция Dcp2Δh2 Δ5H, которая объединяет Dcp2 300Δh2 и Dcp2C Δ5H, имеет ПК 31, что> 2,5 *. 3.5. Таким образом, система соответствует определению синергии. Теперь отметим, что этот эффект может быть результатом кооперативного связывания с молекулами в теле P.
Авт. Используют структурно-функциональное исследование DCP2, чтобы доказать, что связность, аффинность и каталитическая активность являются тремя основными факторами для рекрутирования DCP2 P телец и размера и количества P телец.Однако необходимо провести более конкретные эксперименты, чтобы укрепить этот аргумент или обобщить открытие для других компонентов P-телец или других типов RNP-телец: 1) проведение исследования делеции домена / сайта связывания на Edc3 или других коровых белках; 2) добавление валентного или высокоаффинного сайта связывания к клиентоподобному белку, чтобы превратить его в каркас (достаточность). 3) Тестирование модели с белками скаффолда стрессовых гранул.
Мы согласны с тем, что наши аргументы будут подкреплены дополнительными экспериментами, которые запланированы для дополнительных проектов и публикаций в будущем.
В целом, это исследование технически выполнено хорошо, но довольно ограничено по своему содержанию или новизне. Истинное функциональное считывание отсутствует, поскольку экспериментальная установка не связана с функцией P-тельца (например, с уровнем РНК в различных гранулах и активностью по удалению колпачков). Рукопись можно легко сжать до четырех рисунков, каждая из которых представляет собой одну точку для характеристики конкретного типа конденсата: Рисунок 1 и 2: обогащение определенных факторов в P-телах, Рисунки 3 и 4 соотношение концентраций между компонентами и в различных условиях. и рисунки 5 и 6 структурный анализ одного компонента, предполагающий частичную избыточность между спиралями CTD и рисунок 7 с исследованиями, касающимися связывания РНК.
Мы согласны с тем, что мы не пытались рассматривать функцию тела P в этой работе. Мы считаем, что такой анализ потребует отдельной публикации. Учитывая отсутствие ограничений по цифрам в eLife , мы не сжимали данные в меньшее количество цифр, так как мы опасаемся, что это затруднит чтение и понимание статьи (например, рисунок 4 не предназначен для сравнения различных условий, но скорее, чтобы подчеркнуть, что большинство молекул не присутствует в P-тельцах для большинства видов.Отсюда его название: «Р-тельца не сильно изолируют свои резидентные белки».
Рецензент № 3:
В этой переработанной рукописи Xing et al. проанализировать обогащение и динамические свойства белков тельца ~ 20 P в дрожжах. Описательная часть исследования достаточно обширна (31 белок включен в первоначальный обзор, 19, численность которых была определена количественно) и предполагает существование двух групп: белки, сильно обогащенные P-тельцами, некоторые из которых также необходимы для сборки P-телец — и менее обогащенные белки — меньшее количество которых влияет на сборку P-телец.Исходя из этого, авторы предполагают, что P-тельца «биохимически проще, чем предполагает протеомика». В лучшем случае это предположение, а в худшем — грубое упрощение, поскольку изобилие не обязательно является предиктором функции — особенно для ферментов.
Для ясности мы изменили вывод на «композиционно проще, чем предполагает протеомика». Это должно устранить основную озабоченность рецензента. Однако мы отмечаем, что наши данные на рисунке 4 противоречат утверждению рецензента о том, что численность не предсказывает функцию.Этот рисунок показывает, что для почти всех белков, обогащенных P-тельцами, подавляющее большинство молекулярных видов составляют , а не в организме, а скорее присутствуют в цитоплазме. Для всех молекул, кроме шести верхних, менее 20% находится в теле, даже при консервативной поправке на ненаблюдаемые субдифракционные Р-тела (без этой поправки значение падает до 8%). Таким образом, трудно утверждать, что количество P-тельца имеет большое значение для общей активности этого вида в клетке.Есть способы обойти эту проблему, включая, например, огромное увеличение удельной активности (активности на молекулу) внутри P-тельца по сравнению с цитоплазмой или совместную концентрацию нескольких молекул в каскаде. Но это требует дополнительных предположений, и, по нашему мнению, точка зрения первого порядка состоит в том, что изобилие действительно играет важную роль в формировании биохимических функций, возникающих из конденсатов, и, поскольку только небольшое количество молекул является высококонцентрированным, компартменты фактически являются , вероятно, будет биохимически проще, чем можно было бы представить из исследования протеомики, которое не определяет концентрации и, таким образом, по существу взвешивает все компоненты одинаково.
Во второй части исследования авторы исследуют домены в DCP2, необходимые для локализации P тела, и идентифицируют множественные домены, которые, по-видимому, функционируют частично избыточно. Они пришли к выводу, что совокупность синергетических взаимодействий управляет сборкой P-телец, к этому выводу пришли несколько других исследований (рассмотренные в Lu Na and Slavoff, 2018), включая анализы in vitro (Shutz et al., 2017).
В целом, у нас осталось исследование, которое будет полезным справочным материалом для будущих исследований тела P, но оно не соответствует обещанию «принципов состава и специфичности».В частности, вопрос специфичности (почему не все РНК-связывающие белки локализуются в P-тельцах?) Напрямую не рассматривается. Авторы проводят исследования структуры-функции только одного белка Р-тельца и никогда не рассматривают, что исключает другие РНК-связывающие белки из Р-тельцов.
Как указано выше, мы считаем, что наша работа действительно затрагивает специфичность рекрутирования белков в значительной степени посредством идеи синергетического рекрутирования Dcp2 в P тельца. То есть, резидентные белки часто содержат несколько слабых элементов рекрутирования, и, как следствие, белки, которые содержат мало таких элементов, рекрутируются плохо или вообще не рекрутируются.Рекрутирование РНК может следовать схожим принципам, и теперь мы прямо упоминаем эту идею.
Наша работа также предполагает, что другие связывающие РНК белки действительно накапливаются в Р-тельцах, но их уровень просто отражает количество доступных сайтов связывания в мРНК Р-тельцов. В частности, хотя РНК-связывающий белок MS2 обычно не накапливается в Р-тельцах, добавление множества сайтов связывания для этого белка в мРНК, резидентную в Р-тельцах, может привести к накоплению MS2 в Р-тельцах.Во-вторых, мы действительно наблюдаем другие РНК-связывающие белки в Р-тельцах, просто они менее эффективно делятся на Р-тельца, чем основные компоненты (рис. 1).
Отметим, что исключение молекул из конденсата (PC <1) будет очень трудно наблюдать и значимо количественно оценить в клетках, учитывая небольшой размер структур. Более того, более вероятно, что большинство молекул просто не будет задействовано (PC ~ 1), чем будет открыто исключено; мы не знаем о молекулах, исключенных из P-тел.В общем, физические механизмы, которые позволяют исключить образование конденсатов in vivo, вообще не поняты и выходят далеко за рамки настоящего исследования.
Мы также отмечаем, что, насколько нам известно, это первый раз, когда синергизм был количественно продемонстрирован для рекрутирования белка в конденсат in vivo. Упомянутая выше работа Sprangers интересна, но проводится исключительно in vitro. В клетках, чтобы заявить о синергии, необходимо удалить белок, а затем количественно оценить коэффициенты разделения для различных фрагментов при повторном введении индивидуально и вместе, как мы это сделали.Нам неизвестно, чтобы такой уровень количественного анализа проводился где-либо еще.
Дополнительные комментарии:
1) Авторы не последовательны в описании CTD. Во-первых, они предполагают, что «N-концевой домен, HLM1 и C-концевой домен — все необходимы для эффективного разделения и поддержания характерной медленной динамики Dcp2». Однако позже они говорят, что «удаление CTD не влияет на ПК с полноразмерным Dcp2». Представленные данные предполагают, что C-концевой домен не требуется (300-конец), но может компенсировать отсутствие HLM1.
Благодарим рецензента за обнаружение этого несоответствия. Мы изменили первое утверждение на «Поскольку N-концевой домен и HLM1 необходимы для эффективного разделения и поддержания характерной медленной динамики Dcp2, а C-концевой домен может компенсировать отсутствие HLM1, мы заключаем, что элементы, управляющие разделением и динамика распределяется по белку ».
2) Неповрежденный C-концевой домен никогда не тестируется; вместо этого авторы использовали CTD с 5 мутированными доменами HLM.Хотя авторы подразумевают, что эта версия CTD имеет более слабую связь с телами P, данные не показаны, и из текста неясно, почему была использована эта версия CTD.
У нас были эти данные, и теперь мы показываем их на рисунке 5 — рисунок в приложении 1С.
3) Для данных на фиг. 7 авторы используют конструкции DCP2 с пониженным связыванием с Edc3, чтобы исследовать роль связывания РНК и активности декапирования в ассоциации P-телец. Однако неясно, почему связывание РНК изучали с использованием мутанта Dcp2 300, а декапирование изучали на мутанте Dcp2Δh2.
По сути, это были исторические особенности работы, связанные с тем, когда мы получали различные конструкции. Мы не считаем, что различия здесь влияют на наши выводы.
4) Цифра, сравнивающая концентрацию тельцов P / коэффициент распределения и скорости обмена для различных мутантов DCP2 (аналогично рис. 2A и B), может быть более интуитивным способом сравнения всех различных мутантов DCP2, использованных в этом исследовании.
Мы считаем, что, хотя в некоторых отношениях такой сюжет может помочь, в других он отвлекает внимание, и не включает его.Принципиальный недостаток состоит в том, что на рисунке 2 мы специально подчеркиваем корреляцию между концентрацией тел P и обменными курсами. Но точки на рисунках 5 и 6 отличаются, и они сосредоточены на понимании различных аспектов рекрутирования Dcp2.
5) Исследования структурной функции DCP2 идентифицируют домены в DCP2, которые способствуют локализации P-телец. Достаточно ли этих доменов, чтобы направить белок, не являющийся P-телом, в P-тельца, не решается. Без таких «достаточных» экспериментов остается неясным, можно ли обобщить результаты для DCP2 для других белков.Например, они предполагают, что Xrn1 стабильно рекрутируется из-за его высокой аффинности связывания РНК, однако эта гипотеза не объясняет, почему другие высокоаффинные связывающие РНК белки исключаются из P тельцов.
В этом комментарии рассматриваются два момента. Во-первых, могут ли домены внутри Dcp2, которые способствуют локализации P-телец, действовать доминирующим образом. По сути, мы продемонстрировали этот феномен с GFP как чужеродным белком. Связанный с этим вопрос, но один выходит за рамки этой рукописи: может ли нацеливающий домен P-тельца Dcp2 перекрывать свойства других связывающих РНК белков, которые либо исключают их из P-тельцов, либо направляют эти связывающие РНК белки на другие конденсаты.