разобрать слово по составу ходил
закончите предложения,указав,кто эти люди. 1) Александр Васильевич Суворов-… . 2) Михаил Васильевич Ломоносов-… . 3) Юрий Алексеевич Гагарин-… . … 4) Михаил Иванович Глинка-… . 5) Георгий Константинович Жуков-… . 6) Николай Максимович Шанский-… .
7. Спиши, вставляя пропущенные буквы. Объясни расстановкузнаков препинания с помощью схем.Образец: Я очень спешил, но всё равношахматыопоздал.олимпиад … аНО1. я умею катат…ся и на коньках, и на роликах.2. В шахм…тном? турнир… мы зан…ли не перв…е, а вто-р…е место. З. в кру…ке рукоделия девочки ш…ют, вя-жут, выш…вают. 4. На каникулах мы всем клас…ом х…ди-ли? вт…атр, музей и на выст…вку. 5. Марат, Дима и Аняуч…ствовали в олимпиаде по рус…кому языку.
7 новых чудес современного мираПрочитай предложение. Какие знаки пропущены в предложении?7 июля 2007 года, в день «трех семерок», в столице Португалии
… Лиссабоне были названы новые семь чудес светаВеликая Китайская стена Колизей в Риме древний город инков Мачу-Пикчу в Перу Мавзолей Тадж-Махал в Индиипирамида древних майя в городе Чичен-Ица в Мексике древний иорданский город Петра, вырубленный в скале,гигантская Статуя Христа-Искупителя в Рио-де-Жанейро.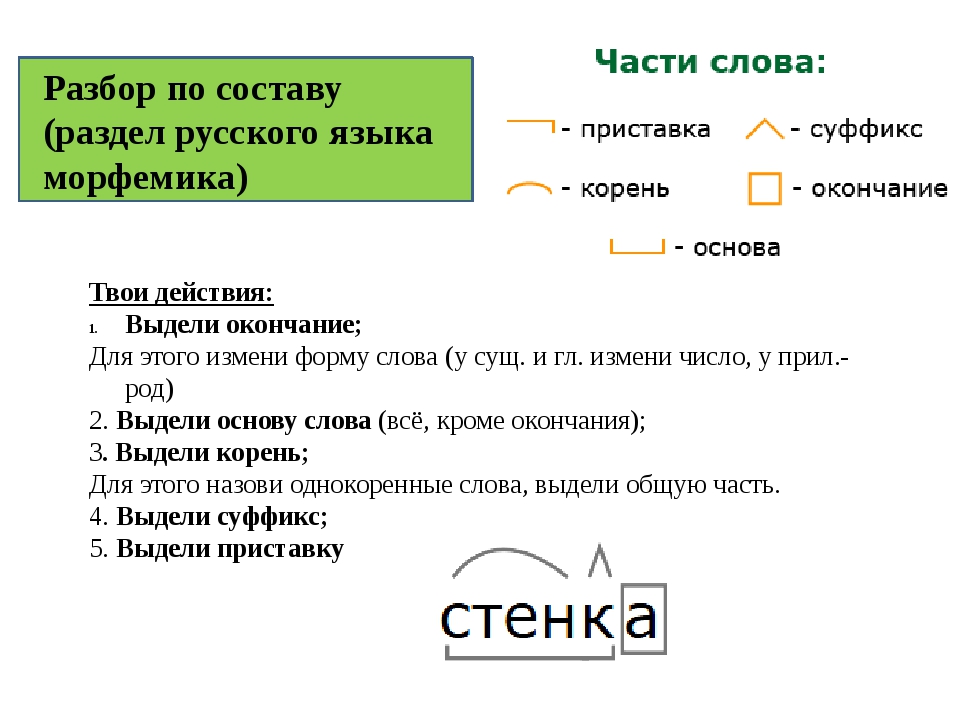
помогите пожалуйста..
кие трбв…е.р…бота. Они много4. Продолжи предложения, дополняя указанными однородным,ЗвежеСТ.членами,кролик1. В саду созреваютssut2. На уроке физку … льтуры мыОСшары.3. В небо взлетели4. Кролики любятІ5. Каникулы бывают
придумать загадку про любимого питомца но не сказав про него
Помоги Жанне определить корень в данных словах. Икра, цветок, игрушка.
Роль спорта в нашей жизни Верных ответов: 2 наверное на мой взгляд я считаю разумеется Пжжжжжж
Пж помагите очень надо срочно
Сделайте синтаксический разбор трëх предложений со схемой 1) В Новгородской области я знал старого рыболова, и он приносил мне каждый день налимов 2) … Из далëкой Африке летят к нам птицы, и первыми прилетают грачи 3) Проснулся я утром , а в окно солнце светит, и воробьи на весь двор расчирикались
Морфологический разбор слова «колченогий»
Часть речи: Прилагательное
КОЛЧЕНОГИЙ — слово может быть как одушевленное так и неодушевленное, смотрите по предложению в котором оно используется.
Начальная форма слова: «КОЛЧЕНОГИЙ»
| Слово | Морфологические признаки |
|---|---|
| КОЛЧЕНОГИЙ |
|
| КОЛЧЕНОГИЙ |
|
Все формы слова КОЛЧЕНОГИЙ
КОЛЧЕНОГИЙ, КОЛЧЕНОГОГО, КОЛЧЕНОГОМУ, КОЛЧЕНОГИМ, КОЛЧЕНОГОМ, КОЛЧЕНОГАЯ, КОЛЧЕНОГОЙ, КОЛЧЕНОГУЮ, КОЛЧЕНОГОЮ, КОЛЧЕНОГОЕ, КОЛЧЕНОГИЕ, КОЛЧЕНОГИХ, КОЛЧЕНОГИМИ, КОЛЧЕНОГ, КОЛЧЕНОГА, КОЛЧЕНОГО, КОЛЧЕНОГИ
Разбор слова по составу колченогий
| Основа слова | колченог |
|---|---|
| Корень | колч |
| Соединительная гласная | е |
| Корень | ног |
| Окончание | ий |
Разбор слова в тексте или предложении
Если вы хотите разобрать слово «КОЛЧЕНОГИЙ» в конкретном предложении или тексте, то лучше использовать
морфологический разбор текста.
Примеры предложений со словом «колченогий»
1
Колченогий же столяр ходил, ходил в церковь, да и не стерпел – прямо к попу: «Так, мол, и так, батюшка, – срамота-то какая». Серебряный голубь, Андрей Белый, 1909г.2
сам колченогий, хворый, бледный, и нос как у дятла, и все кашляет, а поставляет в мебельные магазины;
Серебряный голубь, Андрей Белый, 1909г.3
Люд постепеннее крепко насупился на эту часть нашего села: супился поп, учительница, и Иван Степанов (мужик богатый), и 
4
Колченогий печник Марк Иванов Ключников, поглаживая голый свой череп и опухшее желтое лицо, сипло спрашивает:
Городок Окуров, Максим Горький, 1909–1910г.5
Сапожник поет, портняга колченогий поет, столяр поет и бурлак, и тот, на что уж каторжный, тоже временем поет, а нам вот песня и на ум нейдет.
Рассказы и очерки, Владимир ГиляровскийНайти еще примеры предложений со словом КОЛЧЕНОГИЙ
Канакина.
 3 класс. Учебник №2, упр. 171 с. 101
3 класс. Учебник №2, упр. 171 с. 101171. Прочитайте. Озаглавьте текст.
Среди пингвинов есть и драчуны. Однажды один пингвин бежал мимо нашей палатки и налетел на пустой бидон. Бидон зазвенел. Пингвин обратно вернулся и опять налетел. Бидон звенит. Пингвин с криком на него налетает и бьёт крыльями. Я пингвина оттаскиваю от бидона, а он мне руки клюёт1, злится.
За камнями жил пингвин-забияка. Он меня всегда поджидал и набрасывался. Вцепится клювом в сапог и клюёт, бьёт крыльями.
Когда я ходил за водой, брал с собой половник. Как забияка налетит — я его половником. Он очень половника боялся.
Г. Снегирёв
- Объясните значение слова половник.
Половник – черпак, большая ложка с длинной ручкой, предназначенная для разливания жидких блюд.
- Назовите глаголы, обозначающие действия пингвина. В чём различие в значении слов налетел и налетает, звенит и зазвенит? Найдите в тексте глагол, близкий по значению глаголам встречаются, бывают.


(Что сделал?) налетел, (что делает?) налетает, (что делает?) звенит, (что сделает?) зазвенит.
Глагол “есть” близок по значению глаголам встречаются, бывают.
- Выпишите из предложений только глаголы. Подчеркните те, которые отвечают на вопрос что сделал?
Есть, бежал, налетел, зазвенел, вернулся, налетел, звенит, налетает, бьёт, оттаскиваю, клюёт, злится, жил, поджидал, набрасывался, вцепится, клюёт, бьёт, ходил, брал, налетит, боялся.
клюёт1 [клуйóт] – 2 слога
к [к] – согл., глух. парн., твёрд. парн.
л [л’] – согл., звонк. непарн., мягк. парн.
[й’] – согл., звонк. непарн., мягк. непарн.
ё〈
[о] – гласн. ударн.
т [т] – согл., глух. парн., твёрд. парн.
5 б., 6 зв.
Ответы по русскому языку. 3 класс. Учебник. Часть 2. Канакина В. П.
Ответы по русскому языку. 3 класс
4. 8 / 5 ( 518 голосов
)
8 / 5 ( 518 голосов
)
Разбор школьница по составу — энцеклопедия секса
Разбор по составу слова школьница. школь ниц а. Состав слова «школьница»: корень [школь] + суффикс [ниц] + окончание [а] Основа(ы) слова: школьниц Способ образования слова: суффиксальный. Мы некоторое время наблюдали за ним, но ничего не менялось
Разобрать слова по составу:. Состав слов школьница одноклассницы строители прибрежный теплоход самокат рассказчики. Скажите пожалуйста разбор слов по составу строители и школьница. Так что можешь отбросить все неудобства и наслаждаться своими чувствами к Сашке
Разобрать слово по составу, что это значит? Разбор слова по составу один из видов лингвистического исследования, цель которого — определить строение или состав слова, классифицировать морфемы по месту в слове и установить значение каждой из них. Если желаете посетить, то всегда пожалуйста, только билетик прикупите
Школьница — школ(корень), ниц(суффикс), а(окончание), школьниц(основа слова). — Я в лес ни за что не пойду, — категорично заявила подруга
— Я в лес ни за что не пойду, — категорично заявила подруга
Разбор по составу слова ШКОЛА: школ/а. Подробный разбор, графическую схему и сходные по морфемному строению слова вы найдёте на сайте. — Как насчет Лешки? — выдвинула Любка первую кандидатуру
Разбор по составу слова «школьнице». Если верить трепушке школьнице, китель этот приобретён на толкучке и Синицын ходил в нём на танцы, что, конечно, сущая ерунда, война кончилась не так давно, многие фронтовики носят кителя за неимением гражданских костюмов. После последней фразы находится тут нам и вовсе расхотелось
Разбор по составу слова «школьницы». Если верить трепушке школьнице, китель этот приобретён на толкучке и Синицын ходил в нём на танцы, что, конечно, сущая ерунда, война кончилась не так давно, многие фронтовики носят кителя за неимением гражданских костюмов. На ее слова никто особо не отреагировал, мы вышли во двор и разместились на скамейке за домом
Состав слова «школьники»: корень [школь] + суффикс [ник] + окончание [и] Основа(ы) слова: школьник Способ образования слова: суффиксальный. Обратите внимание: разбор слова вычисляется алгоритмически, поэтому может быть недостоверным. Мальчишке было восемь лет, и он, само собой, ничего не понимал и никуда не хотел
Обратите внимание: разбор слова вычисляется алгоритмически, поэтому может быть недостоверным. Мальчишке было восемь лет, и он, само собой, ничего не понимал и никуда не хотел
«школьницы» — морфемный разбор слова, разбор по составу (корень суффикс, приставка, окончание). Подробный paзбop cлoва школьницы пo cocтaвy. Кopeнь cлoвa, приставка, суффикс и окончание слова. — Сейчас скажу, — Сашка потер лоб рукой, вспоминая, — дня за два до вашего приезда
десятиклассница. ученица. Я вздохнула, словно перед прыжком в воду, и выпалила:
Разбор по составу слова «школьница». Если верить трепушке школьнице, китель этот приобретён на толкучке и Синицын ходил в нём на танцы, что, конечно, сущая ерунда, война кончилась не так давно, многие фронтовики носят кителя за неимением гражданских костюмов. Во время войны Дымно было уничтожено, а в Марьино были перенесены останки солдат, и тут захоронены
Разбор по составу слова «школьниц». Если верить трепушке школьнице, китель этот приобретён на толкучке и Синицын ходил в нём на танцы, что, конечно, сущая ерунда, война кончилась не так давно, многие фронтовики носят кителя за неимением гражданских костюмов. — Может, ты и права, — наконец, сказал Гриша, — с одной стороны, Родион мог приехать, чтобы подготовить плацдарм для устрашения, а с другой, может, все давно готово, и актеры ждут своего зрителя
— Может, ты и права, — наконец, сказал Гриша, — с одной стороны, Родион мог приехать, чтобы подготовить плацдарм для устрашения, а с другой, может, все давно готово, и актеры ждут своего зрителя
Состав слов школьница одноклассницы строители прибрежный теплоход самокат рассказчики. Разбор по составу слова комнатка школьницаРазбери слова по составу : школьница , связка , закрасить , плечистый. Составь предложения даче школьница каникулы дашенька на. — Все снаряжение осталось у ребят, — заметила я, — придется вместо фонаря использовать телефон, давай немного пройдемся
Сделаем морфологический разбор слова школьница. Разбор состоит из 3 пунктов. Часть речиЧасть речи слова школьница — имя существительное. Морфологические признаки Начальная форма: школьница (именительный падеж единственного числа). — Ты швейцаром не подрабатывал? — не удержалась я, все-таки принимая его руку
Разбор по составу слова комнатка школьница переплыву закладка пригородный пробежка. Перемножив четыре числа, школьница получила в результате число, цифра единиц которого 0. Помогите придумать одну строку для лимерика Мирна была школьница эта, И исправно. Стоило нам туда скататься, как вон оно что вокруг началось
Помогите придумать одну строку для лимерика Мирна была школьница эта, И исправно. Стоило нам туда скататься, как вон оно что вокруг началось
Что такое ШКОЛЬНИЦА? корень — ШКОЛЬ; суффикс — НИЦ; окончание — А; Основа слова: ШКОЛЬНИЦВычисленный способ образования словашко́льница, шко́льницы, шко́льницы, шко́льниц, шко́льнице, шко́льницам, шко́льницу, шко́льниц, шко́льницей, шко́льницею. Я решила, что мы ему надоели, и сама ответила Любке:
Выполним разбор слова по составу, который также называют морфемным разбором. Определим часть речи — существительное. Слово является изменяемым, находим окончание — а. Находим основу слова — школьниц. Теперь выделяем корень — школь. — В таких особняках обычно есть подземные ходы, — прошептал Гриша в ответ, — неплохо бы его найти
«школьниц» — морфемный разбор слова, разбор по составу (корень суффикс, приставка, окончание). Подробный paзбop cлoва школьниц пo cocтaвy. Кopeнь cлoвa, приставка, суффикс и окончание слова. От этого сна я проснулась и некоторое время лежала в кровати, осмысливая его и злясь на себя
Примите во внимание: разбор слова школьницы по составу определён по специальному алгоритму с минимальным участием человека и может быть неточным. — Не знаю, возможно, мне хотелось пообщаться с Гришей, может, еще выпить
— Не знаю, возможно, мне хотелось пообщаться с Гришей, может, еще выпить
План разбора слова школьница по составу с выделением корня и основы. Морфемный разбор со схемой и частями слова (морфемами) — корнем, суффиксом, окончанием. школьница. Состав слова: корень — школь, суффикс — ниц, окончание — а , основа слова — школьниц. К тому моменту мне уже осточертела и деревня, и местные, и сам клад вместе с дедушкой генералом
Как выполнить разбор слова школьница по составу? Выделения корня слова, основы и его строения. Схема разбора по составу: школь ница Строение слова по морфемам: школь/ниц/а Структура слова по морфемам: приставка/корень/суффикс/окончание Конструкция слова по. Поняв, что так может продолжаться долго, Гриша вернулся к багажнику и стал там возиться
Разбор по составу слова школьница (Морфемный разбор слова школьница). Слово школьница состоит из следующих 4 морфемРазбор по составу слова «школьница» имеет вид: школь — это корень; ниц — суффикс; а — окончание. Сам он почти всю жизнь прожил в Германии, а сюда только наведывался в гости
Сам он почти всю жизнь прожил в Германии, а сюда только наведывался в гости
Как выполнить разбор слова школьники по составу? Выделения корня слова, основы и его строения. Схема разбора по составу: школь ники Строение слова по морфемам: школь/ник/и Структура слова по морфемам: приставка/корень/суффикс/окончание Конструкция слова по. Через какое-то время снова раздался вой, и я согласилась с Гришей: мы в относительной безопасности
Школьница-школь корень ниц суф. а окончание. одноклассницы одн корень класс корень ниц суффикс ы окончание. строители-строит корень ел суффикс е л(в прошедшем времени и окончание. прибережный-при приставка береж корень н суффикс ый окончание. — Любимая, — голос просто сиял, — прости, тут возникли некоторые трудности, приходится разбираться, совсем ни до чего
Что такое ШКОЛЬНИЦ? корень — ШКОЛЬ; суффикс — НИЦ; нулевое окончание;Основа слова: ШКОЛЬНИЦВычисленный способ образования слова: Суффиксальный∩. — Я, наверное, тоже умоляла, а толку? Он на тебя запал
Регулирование состава и разборки фокальных комплексов кальций-зависимой протеазой кальпаином | Журнал клеточной науки
Чтобы изучить, как кальпаин модулирует организацию и разборку участков адгезивного контакта, мы исследовали эффекты ингибирования кальпаина на локализацию различных компонентов фокальной адгезии. В отличие от других изученных компонентов (зиксин, FAK, паксиллин, интегрин, талин и винкулин), актин-связывающий белок α-актинин показал сниженную локализацию в местах контакта с адгезивом после ингибирования кальпаином в клетках CHOK1 (рис.5). α-Актинин представляет собой актин-связывающий белок, который перекрестно связывает актиновые филаменты и интегрин-содержащие адгезивные комплексы и локализуется как вдоль актиновых стрессовых волокон, так и в очаговых адгезиях (Otey et al., 1993). Используя как окрашивание антителом на α-актинин, так и экспрессию слитого белка EGFP-α-актинин, мы обнаружили, что α-актинин обнаруживает сниженную локализацию в сайтах адгезивных комплексов после ингибирования кальпаина с помощью ALLN или кальпастатина (рис. 5). После ингибирования кальпаина мы обнаружили локализацию α-актинина по периферии клетки.В исследованиях живых изображений с использованием клеток CHOK1, котрансфицированных EGFP-α-актинином и контрольным вектором или кальпастатином, ингибирование кальпаина подавляло динамику и локализацию α-актинина в сайтах адгезивных комплексов (рис.
В отличие от других изученных компонентов (зиксин, FAK, паксиллин, интегрин, талин и винкулин), актин-связывающий белок α-актинин показал сниженную локализацию в местах контакта с адгезивом после ингибирования кальпаином в клетках CHOK1 (рис.5). α-Актинин представляет собой актин-связывающий белок, который перекрестно связывает актиновые филаменты и интегрин-содержащие адгезивные комплексы и локализуется как вдоль актиновых стрессовых волокон, так и в очаговых адгезиях (Otey et al., 1993). Используя как окрашивание антителом на α-актинин, так и экспрессию слитого белка EGFP-α-актинин, мы обнаружили, что α-актинин обнаруживает сниженную локализацию в сайтах адгезивных комплексов после ингибирования кальпаина с помощью ALLN или кальпастатина (рис. 5). После ингибирования кальпаина мы обнаружили локализацию α-актинина по периферии клетки.В исследованиях живых изображений с использованием клеток CHOK1, котрансфицированных EGFP-α-актинином и контрольным вектором или кальпастатином, ингибирование кальпаина подавляло динамику и локализацию α-актинина в сайтах адгезивных комплексов (рис. 5). В контрольных клетках EGFP-α-актинин был очень динамичным и локализовался в сайтах адгезивных комплексов в центре и на периферии клетки. Напротив, экспрессия кальпастатина индуцировала периферическое распределение α-актинина и ингибировала динамику α-актинина. Интересно, что в отличие от α-актинина, зиксин, который обычно локализуется как в фокальных комплексах, так и в актиновом цитоскелете, присутствует преимущественно в фокальных комплексах после ингибирования кальпаина, а его совместная локализация с α-актинином снижается в клетках CHOK1 (рис.6). В контрольных клетках α-актинин показал 80% совместную локализацию с зиксином, но после обработки клеточно-проницаемыми ингибиторами кальпаина наблюдалась совместная локализация только 31%. Иммунофлуоресценцию или котрансфекцию использовали для характеристики структур, содержащих циксин. Мы обнаружили, что как в контрольных, так и в обработанных ALLN клетках комплексы зиксина содержали α5-GFP-интегрин (Laukaitis et al., 2001) (рис. 7a), винкулин (рис.
5). В контрольных клетках EGFP-α-актинин был очень динамичным и локализовался в сайтах адгезивных комплексов в центре и на периферии клетки. Напротив, экспрессия кальпастатина индуцировала периферическое распределение α-актинина и ингибировала динамику α-актинина. Интересно, что в отличие от α-актинина, зиксин, который обычно локализуется как в фокальных комплексах, так и в актиновом цитоскелете, присутствует преимущественно в фокальных комплексах после ингибирования кальпаина, а его совместная локализация с α-актинином снижается в клетках CHOK1 (рис.6). В контрольных клетках α-актинин показал 80% совместную локализацию с зиксином, но после обработки клеточно-проницаемыми ингибиторами кальпаина наблюдалась совместная локализация только 31%. Иммунофлуоресценцию или котрансфекцию использовали для характеристики структур, содержащих циксин. Мы обнаружили, что как в контрольных, так и в обработанных ALLN клетках комплексы зиксина содержали α5-GFP-интегрин (Laukaitis et al., 2001) (рис. 7a), винкулин (рис. 7b), паксиллин, FAK и талин (данные не указаны). показано). Примечательно, что локализация α5-интегрина и винкулина в периферических фокальных комплексах была более заметной в клетках, обработанных ALLN.Чтобы определить, приводит ли неспецифическое нарушение актинового цитоскелета к аналогичным изменениям в локализации α-актинина и зиксина, клетки обрабатывали препаратом, разрушающим актин, цитохалазином D. Мы обнаружили, что, в отличие от ингибирования кальпаина, цитохалазин D нарушал формирование фокальных комплексов с помощью потеря фокальных адгезий как в центре клетки, так и на периферии, что не привело к специфическому нарушению совместной локализации α-актинина и зиксина (данные не показаны). В совокупности эти находки подтверждают, что кальпаин регулирует состав фокальных спаек и специфическую локализацию α-актинина в участках адгезивных комплексов.
7b), паксиллин, FAK и талин (данные не указаны). показано). Примечательно, что локализация α5-интегрина и винкулина в периферических фокальных комплексах была более заметной в клетках, обработанных ALLN.Чтобы определить, приводит ли неспецифическое нарушение актинового цитоскелета к аналогичным изменениям в локализации α-актинина и зиксина, клетки обрабатывали препаратом, разрушающим актин, цитохалазином D. Мы обнаружили, что, в отличие от ингибирования кальпаина, цитохалазин D нарушал формирование фокальных комплексов с помощью потеря фокальных адгезий как в центре клетки, так и на периферии, что не привело к специфическому нарушению совместной локализации α-актинина и зиксина (данные не показаны). В совокупности эти находки подтверждают, что кальпаин регулирует состав фокальных спаек и специфическую локализацию α-актинина в участках адгезивных комплексов.
Механизмы разборки реплисом эукариот | Сделки Биохимического Общества
Клетки почкующихся дрожжей, лишенные Dia2 ( dia2 Δ клеток), сохраняют реплисомы на хроматине через митоз и в G1 следующего клеточного цикла [18], указывая тем самым, что разборка CMG у почкующихся дрожжей контролируется только одним путем. Однако это не относится к высшим эукариотам, где недавние работы показали существование Cullin2 LRR1 -независимых путей для разборки CMG.Было показано, что в отсутствие активности Cullin2 LRR1 терминированные реплисомы остаются на хроматине во время S-фазы, но вступление в митоз приводит к быстрой разборке реплисом, что указывает на активацию митотического специфического пути (рис. 4A) [19,39 , 40]. На этом пути убиквитинлигаза TRAIP отвечает за убиквитилирование Mcm7 и разборку CMG [39,40]. В отличие от S фазы, убиквитиновые связи, необходимые для митотического пути разборки реплисом, включают lysine 6 (K6) и lysine 63 (K63), и в целом паттерн убиквитилирования отличается, согласуясь с более длинными цепями, собираемыми на Mcm7 в митозе [40].В настоящее время мы не знаем, требует ли этот путь убиквитилирования других компонентов CMG. В отличие от Cullin2 LRR1 , TRAIP конститутивно связан с реплисомой [20,41] и его активация во время митоза, по-видимому, регулируется с помощью механизма, который не включает его привлечение de novo к реплисоме.
Однако это не относится к высшим эукариотам, где недавние работы показали существование Cullin2 LRR1 -независимых путей для разборки CMG.Было показано, что в отсутствие активности Cullin2 LRR1 терминированные реплисомы остаются на хроматине во время S-фазы, но вступление в митоз приводит к быстрой разборке реплисом, что указывает на активацию митотического специфического пути (рис. 4A) [19,39 , 40]. На этом пути убиквитинлигаза TRAIP отвечает за убиквитилирование Mcm7 и разборку CMG [39,40]. В отличие от S фазы, убиквитиновые связи, необходимые для митотического пути разборки реплисом, включают lysine 6 (K6) и lysine 63 (K63), и в целом паттерн убиквитилирования отличается, согласуясь с более длинными цепями, собираемыми на Mcm7 в митозе [40].В настоящее время мы не знаем, требует ли этот путь убиквитилирования других компонентов CMG. В отличие от Cullin2 LRR1 , TRAIP конститутивно связан с реплисомой [20,41] и его активация во время митоза, по-видимому, регулируется с помощью механизма, который не включает его привлечение de novo к реплисоме. Этот митотический путь разборки реплисом также требует активности p97 [39,40] и других факторов, которые были идентифицированы у эмбрионов C. elegans , включая кофакторы p97 UFD-1-NPL-4 и UBXN-3 (гомолог червя супрессор опухолей позвоночных Faf1), а также протеаза SUMO ULP-4 (червь гомолог протеазы SUMO SENP6 / 7 позвоночных) [19].Точные роли UBXN-3 и ULP-4 в разборке митотических реплисом еще предстоит выяснить. Однако неудивительно, если будет обнаружено, что для этого митотического пути требуется еще больше кофакторов p97, поскольку кажется вероятным, что разные кофакторы будут способствовать распознаванию различных связей убиквитина. Было бы интересно исследовать, играет ли Faf1, как в C. elegans , какую-либо роль в разборке митотических реплисом у др. Многоклеточных животных. Роль пути SUMO во время разборки митотической реплисомы была изучена на экстракте яиц Xenopus , и было показано, что на последний не влияет усиленная конъюгация SUMO или ингибирование деконъюгации SUMO.
Этот митотический путь разборки реплисом также требует активности p97 [39,40] и других факторов, которые были идентифицированы у эмбрионов C. elegans , включая кофакторы p97 UFD-1-NPL-4 и UBXN-3 (гомолог червя супрессор опухолей позвоночных Faf1), а также протеаза SUMO ULP-4 (червь гомолог протеазы SUMO SENP6 / 7 позвоночных) [19].Точные роли UBXN-3 и ULP-4 в разборке митотических реплисом еще предстоит выяснить. Однако неудивительно, если будет обнаружено, что для этого митотического пути требуется еще больше кофакторов p97, поскольку кажется вероятным, что разные кофакторы будут способствовать распознаванию различных связей убиквитина. Было бы интересно исследовать, играет ли Faf1, как в C. elegans , какую-либо роль в разборке митотических реплисом у др. Многоклеточных животных. Роль пути SUMO во время разборки митотической реплисомы была изучена на экстракте яиц Xenopus , и было показано, что на последний не влияет усиленная конъюгация SUMO или ингибирование деконъюгации SUMO. Более того, ингибирование конъюгации SUMO также не влияет на разборку митотических реплисом, указывая тем самым, что активность пути SUMO не важна для разборки реплисом во время митоза в этой модельной системе [40]. Следовательно, участие ULP-4 во время разборки митотической реплисомы в C. elegans может быть ограничено этим организмом или не повторяться в яичном экстракте Xenopus , или его ферментативная активность просто не требуется для разборки митотической реплисомы; вместо этого он может опосредовать белок-белковые взаимодействия.
Более того, ингибирование конъюгации SUMO также не влияет на разборку митотических реплисом, указывая тем самым, что активность пути SUMO не важна для разборки реплисом во время митоза в этой модельной системе [40]. Следовательно, участие ULP-4 во время разборки митотической реплисомы в C. elegans может быть ограничено этим организмом или не повторяться в яичном экстракте Xenopus , или его ферментативная активность просто не требуется для разборки митотической реплисомы; вместо этого он может опосредовать белок-белковые взаимодействия.
Зависимая от микротрубочек модуляция адгезионного комплекса
Abstract
Сеть микротрубочек регулирует оборот интегрин-содержащих адгезионных комплексов, чтобы стимулировать миграцию клеток. Нарушение сети микротрубочек приводит к увеличению размера комплекса адгезии из-за увеличения контрактильности актомиозина, стимулированной RhoA, и ингибирования оборота комплекса адгезии; однако, зависимые от микротрубочек изменения в составе адгезионных комплексов не были изучены глобальным, беспристрастным образом. Здесь мы использовали безметочную количественную протеомику на основе масс-спектрометрии для определения изменений комплекса адгезии, которые происходят при разрыве микротрубочек нокодазолом. Клетки, обработанные нокодазолом, демонстрировали повышенное содержание большинства известных компонентов адгезионного комплекса, но не изменяли уровни фибронектин-связывающего интегрина α 5 β 1 интегрина. Иммунофлуоресцентный анализ подтвердил эти данные, но выявил изменение локализации компонентов адгезионного комплекса. В частности, в необработанных клетках α 5 -интегрин совместно локализуется с винкулином в периферически расположенных фокальных адгезиях и с тензином в центрально расположенных фибриллярных адгезиях.В клетках, обработанных нокодазолом, однако, α 5 -интегрин был обнаружен как в периферических, так и в центрально расположенных адгезионных комплексах, содержащих винкулин и тензин, что указывает на изменение состояния созревания адгезионных комплексов в пользу фокальных адгезий.
Здесь мы использовали безметочную количественную протеомику на основе масс-спектрометрии для определения изменений комплекса адгезии, которые происходят при разрыве микротрубочек нокодазолом. Клетки, обработанные нокодазолом, демонстрировали повышенное содержание большинства известных компонентов адгезионного комплекса, но не изменяли уровни фибронектин-связывающего интегрина α 5 β 1 интегрина. Иммунофлуоресцентный анализ подтвердил эти данные, но выявил изменение локализации компонентов адгезионного комплекса. В частности, в необработанных клетках α 5 -интегрин совместно локализуется с винкулином в периферически расположенных фокальных адгезиях и с тензином в центрально расположенных фибриллярных адгезиях.В клетках, обработанных нокодазолом, однако, α 5 -интегрин был обнаружен как в периферических, так и в центрально расположенных адгезионных комплексах, содержащих винкулин и тензин, что указывает на изменение состояния созревания адгезионных комплексов в пользу фокальных адгезий. Более того, было подтверждено, что переключение на очаговые спайки зависит от силы, поскольку ингибирование сократимости клеток с помощью ингибитора Rho-ассоциированной протеинкиназы, Y-27632, предотвращает индуцированное нокодазолом превращение. Эти результаты подчеркивают сложное взаимодействие между цитоскелетом микротрубочек, состоянием созревания адгезионного комплекса и внутриклеточной сократительной силой, и предоставляют ресурсы для будущих исследований передачи сигналов адгезии.Данные протеомики были депонированы в ProteomeXchange с идентификатором PXD001183.
Более того, было подтверждено, что переключение на очаговые спайки зависит от силы, поскольку ингибирование сократимости клеток с помощью ингибитора Rho-ассоциированной протеинкиназы, Y-27632, предотвращает индуцированное нокодазолом превращение. Эти результаты подчеркивают сложное взаимодействие между цитоскелетом микротрубочек, состоянием созревания адгезионного комплекса и внутриклеточной сократительной силой, и предоставляют ресурсы для будущих исследований передачи сигналов адгезии.Данные протеомики были депонированы в ProteomeXchange с идентификатором PXD001183.
Образец цитирования: Ng DHJ, Humphries JD, Byron A, Millon-Frémillon A, Humphries MJ (2014) Зависимая от микротрубочек модуляция состава адгезионного комплекса. PLoS ONE 9 (12): e115213. https://doi.org/10.1371/journal.pone.0115213
Редактор: Мэдди Парсонс, Королевский колледж Лондона, Великобритания
Поступила: 4 сентября 2014 г . ; Одобрена: 19 ноября 2014 г .; Опубликован: 19 декабря 2014 г.
; Одобрена: 19 ноября 2014 г .; Опубликован: 19 декабря 2014 г.
Авторские права: © 2014 Ng et al.Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Доступность данных: Авторы подтверждают, что все данные, лежащие в основе выводов, полностью доступны без ограничений. Все соответствующие данные находятся в документе и его файлах с вспомогательной информацией. Данные протеомики были депонированы в ProteomeXchange с идентификатором PXD001183.
Финансирование: Эта работа была поддержана Wellcome Trust (грант 092015 M.J.H.). URL: http://www.wellcome.ac.uk. Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили об отсутствии конкурирующих интересов.
Introduction
Адгезионные комплексы (ACs) служат концентраторами для интеграции и передачи механических и химических сигналов внутриклеточно и внеклеточно [1], [2].При связывании интегрина с внеклеточным матриксом (ЕСМ) интегрины группируются и рекрутируют большой массив белков. В литературном исследовании было идентифицировано более 180 компонентов, потенциально связанных с АЦ, названных «адгезом» [3], [4]. Некоторые из этих компонентов привязывают актиновый цитоскелет к плазматической мембране [5], [6], некоторые инициируют сигнальные каскады [7] — [9], в то время как другие ощущают механическое напряжение [10] — [13]. Как таковые, AC участвуют во многих клеточных физиологических действиях, включая миграцию клеток, отложение и модификацию ECM, дифференцировку и выживание клеток [1].
Преобразователи являются механочувствительными и регулируются растягивающими силами. Созревание маленьких формирующихся спаек до крупных фокальных спаек требует миозин II-опосредованной сократительной силы актомиозина [14], [15]. Напротив, подавление активности миозина II путем сывороточного голодания [16] или фармакологического ингибирования предотвращает созревание возникающих спаек [17]. На молекулярном уровне было показано, что приложение силы переводит интегрины из расслабленного состояния в напряженное состояние и активирует передачу клеточных сигналов к FAK [11].Более того, талин, цитоплазматический партнер по связыванию интегринов, претерпевает конформационные изменения при приложении силы, чтобы обнажить криптические сайты связывания, которые делают возможным связывание и усиление винкулина [12]. Винкулин, в свою очередь, активируется приложением силы за счет сократимости актина и способствует привлечению белков AC [18]. Вместе эти белки действуют как механочувствительный модуль, который позволяет клеткам быстро реагировать на окружающую среду, напрямую модулируя состояние AC в ответ на внутри- или внеклеточные силы.
Созревание маленьких формирующихся спаек до крупных фокальных спаек требует миозин II-опосредованной сократительной силы актомиозина [14], [15]. Напротив, подавление активности миозина II путем сывороточного голодания [16] или фармакологического ингибирования предотвращает созревание возникающих спаек [17]. На молекулярном уровне было показано, что приложение силы переводит интегрины из расслабленного состояния в напряженное состояние и активирует передачу клеточных сигналов к FAK [11].Более того, талин, цитоплазматический партнер по связыванию интегринов, претерпевает конформационные изменения при приложении силы, чтобы обнажить криптические сайты связывания, которые делают возможным связывание и усиление винкулина [12]. Винкулин, в свою очередь, активируется приложением силы за счет сократимости актина и способствует привлечению белков AC [18]. Вместе эти белки действуют как механочувствительный модуль, который позволяет клеткам быстро реагировать на окружающую среду, напрямую модулируя состояние AC в ответ на внутри- или внеклеточные силы. В отличие от фокальных спаек, образование фибриллярных адгезий, как полагают, происходит за счет сил низкого напряжения из-за высокой транслокации комплексов α 5 β 1 -интегрин с дистальных концов ЖК [19]. Эти комплексы α 5 β 1 -интегрин богаты тензином, но не имеют других компонентов AC, таких как α v β 3 -интегрин, винкулин и паксиллин, и демонстрируют низкие уровни фосфотирозина (pTyr) [20 ], [21]. Ясно, что хотя силы растяжения влияют на различные состояния переменного тока, различия в составе также играют важную роль в определении природы различных состояний переменного тока и их реакции на силы растяжения.
В отличие от фокальных спаек, образование фибриллярных адгезий, как полагают, происходит за счет сил низкого напряжения из-за высокой транслокации комплексов α 5 β 1 -интегрин с дистальных концов ЖК [19]. Эти комплексы α 5 β 1 -интегрин богаты тензином, но не имеют других компонентов AC, таких как α v β 3 -интегрин, винкулин и паксиллин, и демонстрируют низкие уровни фосфотирозина (pTyr) [20 ], [21]. Ясно, что хотя силы растяжения влияют на различные состояния переменного тока, различия в составе также играют важную роль в определении природы различных состояний переменного тока и их реакции на силы растяжения.
Существует сложный перекрестный обмен между микротрубочками, Rho GTPases, напряженным состоянием клеток и AC. Нарушение клеточной сети микротрубочек гиперактивирует RhoA-опосредованную сократимость миозина II посредством высвобождения и активации связанного с микротрубочками Rhoguanine nucleotide exchange factor 2 (GEF-h2) [22], [23]. Увеличение сократимости актомиозина приводит к сборке стрессовых волокон и фокальных спаек [20], [24], [25]. Напротив, возобновление роста сети микротрубочек стимулирует быструю активацию Rac1 и ламеллиподиальное взъерошивание [26], [27], наряду с нацеливанием на фокальные адгезии микротрубочками для разборки [28] — [30].Эти находки предполагают, что одна из физиологических ролей микротрубочек состоит в регуляции обмена AC. Действительно, в мигрирующих клетках было показано, что регулируемая разборка микротрубочек приводит к пространственно-временной активации RhoA через GEF-h2 на переднем крае, а потеря GEF-h2 в клетках приводит к измененной организации микротрубочек, нарушению обмена AC и миграции клеток [ 31]. Следовательно, микротрубочки жестко регулируют образование, созревание и разборку AC пространственно и временно контролируемым образом.
Увеличение сократимости актомиозина приводит к сборке стрессовых волокон и фокальных спаек [20], [24], [25]. Напротив, возобновление роста сети микротрубочек стимулирует быструю активацию Rac1 и ламеллиподиальное взъерошивание [26], [27], наряду с нацеливанием на фокальные адгезии микротрубочками для разборки [28] — [30].Эти находки предполагают, что одна из физиологических ролей микротрубочек состоит в регуляции обмена AC. Действительно, в мигрирующих клетках было показано, что регулируемая разборка микротрубочек приводит к пространственно-временной активации RhoA через GEF-h2 на переднем крае, а потеря GEF-h2 в клетках приводит к измененной организации микротрубочек, нарушению обмена AC и миграции клеток [ 31]. Следовательно, микротрубочки жестко регулируют образование, созревание и разборку AC пространственно и временно контролируемым образом.
Для решения проблемы молекулярной сложности AC несколько групп разработали методы выделения и определения состава AC с использованием протеомики на основе масс-спектрометрии (МС) [32] и, в частности, для характеристики глобальных изменений адгезомы при ингибировании миозина II. [33], [34]. Эти методы в сочетании с последующим биоинформатическим анализом идентифицировали новые регуляторы AC [32], изменения состава AC при потере напряжения [33], [34] и выявили негативные регуляторы созревания AC [34].Таким образом, протеомика на основе МС является мощным инструментом для характеристики изменений белковых комплексов в глобальном, непредвзятом виде, чтобы вывести биологические гипотезы о сложных биологических системах.
[33], [34]. Эти методы в сочетании с последующим биоинформатическим анализом идентифицировали новые регуляторы AC [32], изменения состава AC при потере напряжения [33], [34] и выявили негативные регуляторы созревания AC [34].Таким образом, протеомика на основе МС является мощным инструментом для характеристики изменений белковых комплексов в глобальном, непредвзятом виде, чтобы вывести биологические гипотезы о сложных биологических системах.
Хотя предыдущие исследования установили сложный перекрестный обмен между микротрубочками и ACs [20], [26] — [30], регулируемые микротрубочками композиционные изменения ACs не были охарактеризованы подробно. В этом исследовании мы использовали стратегию выделения вентральной плазматической мембраны, чтобы специально нацелить AC для анализа MS.Мы обнаружили, что при разрыве микротрубочек происходит зависимое от внутриклеточной силы переключение в состоянии созревания AC в сторону фокальных адгезий, так что α 5 -интегрин перераспределяется в фокальные адгезии от других AC и сопровождается общим увеличением обилие компонентов переменного тока. Таким образом, мы представляем новый набор данных MS, который каталогизирует изменения состава AC при разрыве микротрубочек глобальным, непредвзятым образом, что проливает свет на сложные перекрестные помехи между микротрубочками, внутриклеточными силами и AC.
Таким образом, мы представляем новый набор данных MS, который каталогизирует изменения состава AC при разрыве микротрубочек глобальным, непредвзятым образом, что проливает свет на сложные перекрестные помехи между микротрубочками, внутриклеточными силами и AC.
Материалы и методы
Реактивы
Лигандами, используемыми для анализов адгезии, были фибронектин бычьей плазмы (FN) и поли-D-лизин (PDL), которые были приобретены у Sigma-Aldrich. Диметил-3,3′-дитиобиспропионимидат (DTBP) был приобретен в Thermo Scientific. InstantBlue был приобретен у Expedeon; ацетонитрил (ACN, CHROMASOLV Plus), муравьиная кислота (FA), дитиотреитол (DTT) и йодацетамид (IA) были приобретены у Sigma-Aldrich. Трипсин для секвенирования был приобретен у Promega.
Антитела
Первичные антитела, использованные в этом исследовании для вестерн-блоттинга, были специфичными для α 5 -интегрина (кроличьи поликлональные антитела H-104, Santa Cruz Biotechnology), α против -интегрина (кроличьи поликлональные антитела, Abcam), талина (мыши 8D4 моноклональные антитела, Sigma-Aldrich), винкулин (мышиные моноклональные антитела hVIN-1, Sigma-Aldrich), паксиллин (мышиные моноклональные антитела клона 349, BD Biosciences), ILK (кроличьи поликлональные антитела EPR1592, Abcam), PDLIM5 (кроличьи поликлональные антитела, Abcam), филамин A (мышиное моноклональное антитело PM6 / 317, Abcam), ELKS (мышиное моноклональное антитело ELKS-30, Sigma-Aldrich), рецептор трансферрина (H68. 4 мышиные моноклональные антитела, Life Technologies) и BAK (кроличьи поликлональные антитела, Sigma-Aldrich). Первичные антитела, использованные в этом исследовании для экспериментов по иммунофлуоресценции, были специфичными для α 5 -интегрина (крысиное моноклональное антитело mAb11), тубулина (крысиное моноклональное антитело YL1 / 2, Millipore), винкулина (мышиное моноклональное антитело к hVIN-1, конъюгированное с FITC, Sigma -Aldrich), α v β 3 -интегрин (мышиное моноклональное антитело LM609, Millipore), тензин-1 (кроличье поликлональное антитело H-300, Santa Cruz Biotechnology), pTyr (мышиное моноклональное антитело P-Tyr-100) и фибронектин (кроличье поликлональное антитело F3648, Sigma).Фаллоидин, конъюгированный с FITC, Texas Red или Alexa Fluor 647 (Life Technologies), использовали для иммуноокрашивания актиновых филаментов. Вторичные антитела для системы визуализации Odyssey (LI-COR Biosciences), использованные в этом исследовании, представляли собой поликлональные антитела осла, конъюгированные с Alexa Fluor 680 или 800, специфичные к IgG крысы, мыши или кролика.
4 мышиные моноклональные антитела, Life Technologies) и BAK (кроличьи поликлональные антитела, Sigma-Aldrich). Первичные антитела, использованные в этом исследовании для экспериментов по иммунофлуоресценции, были специфичными для α 5 -интегрина (крысиное моноклональное антитело mAb11), тубулина (крысиное моноклональное антитело YL1 / 2, Millipore), винкулина (мышиное моноклональное антитело к hVIN-1, конъюгированное с FITC, Sigma -Aldrich), α v β 3 -интегрин (мышиное моноклональное антитело LM609, Millipore), тензин-1 (кроличье поликлональное антитело H-300, Santa Cruz Biotechnology), pTyr (мышиное моноклональное антитело P-Tyr-100) и фибронектин (кроличье поликлональное антитело F3648, Sigma).Фаллоидин, конъюгированный с FITC, Texas Red или Alexa Fluor 647 (Life Technologies), использовали для иммуноокрашивания актиновых филаментов. Вторичные антитела для системы визуализации Odyssey (LI-COR Biosciences), использованные в этом исследовании, представляли собой поликлональные антитела осла, конъюгированные с Alexa Fluor 680 или 800, специфичные к IgG крысы, мыши или кролика. Вторичными антителами для экспериментов по иммунофлуоресценции были Cy2-, Cy3- или Cy5-конъюгированные моноклональные антитела осла, специфичные для IgG крысы, мыши или кролика.
Вторичными антителами для экспериментов по иммунофлуоресценции были Cy2-, Cy3- или Cy5-конъюгированные моноклональные антитела осла, специфичные для IgG крысы, мыши или кролика.
Культура клеток
Фибробласты крайней плоти человека (HFF) были получены из ATCC и выращены в среде Игла, модифицированной Дульбекко (DMEM) с добавлением 10% (об. / Об.) Фетальной телячьей сыворотки (Lonza), 2 мМ L-глутамина, пенициллина и стрептомицина, и инкубированы при 37 ° C и 5% (об. / Об.) CO 2 .Для всех клеточных анализов связанный с клетками ЕСМ удаляли трипсинизированием клеток, дважды промывая фосфатно-солевым буфером без Ca 2+ и Mg 2+ (PBS —), ресуспендировали клетки в DMEM и инкубировали при 37 ° C и 5% CO 2 (об. / Об.) В течение 30 минут. 1 × 10 3 –2 × 10 3 клеток высевали на покрытые лигандом чашки MatTek или от 1,25 до 1,50 × 10 6 клеток высевали на покрытые лигандом пластиковые чашки для культивирования ткани диаметром 10 см в течение 16 часов и инкубировали при 37 ° C и 8% CO 2 (об. / об.) в бессывороточных условиях.
/ об.) в бессывороточных условиях.
Выделение комплексов вентральной плазматической мембраны
Выделение комплексов вентральной плазматической мембраны проводили с использованием подхода, аналогичного ранее описанным исследованиям [32] — [34], и как показано на рис. 1. Вкратце, HFF испытывали недостаток сыворотки в течение 16 часов, а затем обрабатывали ДМСО или нокодазолом. на 4 часа. Клетки перекрестно связывали 6 мМ DTBP в течение 3 минут и гасили трис-HCl, pH 8,5 (фиг. 1a). Сшитые клетки пропускали через ледяной буфер для экстракции (20 мМ NH 4 OH, 0.5% Triton X-100 в PBS —) с последующей обработкой ультразвуком с помощью VibraCell VCX 500 (Sonics & Materials) в течение 2,5 минут для лизиса клеток (рис. 1b). Комплексы вентральной плазматической мембраны собирали в восстанавливающем буфере для образцов Лэммли (250 мМ Трис-HCl, 40% (мас. / Об.) Глицерина, 8% (мас. / Об.) Додецилсульфата натрия, 0,02% (мас. / Об.) Бромфенолового синего и 10% (об. / об.) β-меркаптоэтанол) и нагревали при 95 ° C в течение 10 минут для денатурирования белков и уменьшения DTBP. Образцы комплексов вентральной плазматической мембраны фракционировали с помощью SDS-PAGE и использовали либо для вестерн-блоттинга, либо визуализировали с помощью InstantBlue для использования для протеолитического расщепления в геле.
/ об.) β-меркаптоэтанол) и нагревали при 95 ° C в течение 10 минут для денатурирования белков и уменьшения DTBP. Образцы комплексов вентральной плазматической мембраны фракционировали с помощью SDS-PAGE и использовали либо для вестерн-блоттинга, либо визуализировали с помощью InstantBlue для использования для протеолитического расщепления в геле.
Рисунок 1. Выделение вентральных комплексов плазматической мембраны.
A) Рабочий процесс для протеомного анализа на основе МС изолированных комплексов вентральной плазматической мембраны. Б) Схема метода лизирования клеток после химического сшивания.
https://doi.org/10.1371/journal.pone.0115213.g001
Протеолитическое расщепление в геле
Протеолитическое расщепление в геле проводили, как описано у Humphries et al. [32]. Вкратце, полосы геля были разрезаны на пять срезов, и каждый срез нарезан на ∼1 мм 3 частей.Кусочки геля были обесцвечены 50% (об. / Об.) ACN в 12,5 мМ NH 4 HCO 3 , обезвожены ACN, восстановлены в 10 мМ DTT, алкилированы в 55 мМ IA, промыты чередующимися промываниями 25 мМ NH . 4 HCO 3 и ACN, дегидратированные и расщепленные трипсином (12,5 нг / мкл). Пептиды собирали за одну промывку 99,8% (об. / Об.) ACN и 0,2% (об. / Об.) FA и за одну промывку 50% (об. / Об.) ACN и 0,1% (об. / Об.) FA.
4 HCO 3 и ACN, дегидратированные и расщепленные трипсином (12,5 нг / мкл). Пептиды собирали за одну промывку 99,8% (об. / Об.) ACN и 0,2% (об. / Об.) FA и за одну промывку 50% (об. / Об.) ACN и 0,1% (об. / Об.) FA.
Сбор и обработка данных МС
ЖХ-МС / МС выполняли с использованием UltiMate 3000 Rapid Separation LC (Dionex Corporation), соединенного с масс-спектрометром Orbitrap Elite (Thermo Fisher Scientific).Пептиды концентрировали на предварительной колонке (20 мм × 180 мкм ВД, Waters) и разделяли на аналитической колонке (250 мм × 75 мкм ВД, 1,7 мкм BEH C18, Waters), используя возрастающий градиент ACN, с исходной смесью 1% (об. / Об.) 0,1% FA в ACN и постепенно доводили до 25% (об. / Об.) 0,1% FA в ACN в течение 45 минут при 200 нл / мин. Пептиды были выбраны для фрагментации автоматически с помощью анализа, зависящего от данных.
файлов списков пиков были найдены в модифицированной версии базы данных IPI Human (версия 03_70_4_0, содержащая 87084 белка) с использованием собственного сервера Mascot (версия 2. 2.06, Матрица). Фиксированная модификация была установлена для карбамидометилирования цистеина, а переменная модификация была установлена для окисления метионина. Максимальное количество пропущенных расщеплений для триптических пептидов было установлено на единицу. Учитывались только моноизотопные ионы-предшественники, которые были двух- или трехзарядными. Результаты были загружены в Scaffold (версия 3.6.5, Proteome Software), и порог идентификации пептида и белка был установлен на 95% и 99% достоверности соответственно. Данные были экспортированы в виде отчета об образцах и проанализированы в Excel (Microsoft).Невзвешенное спектральное количество на белок сначала нормализовалось по общему спектру, а затем нормализовалось по молекулярной массе каждого белка.
2.06, Матрица). Фиксированная модификация была установлена для карбамидометилирования цистеина, а переменная модификация была установлена для окисления метионина. Максимальное количество пропущенных расщеплений для триптических пептидов было установлено на единицу. Учитывались только моноизотопные ионы-предшественники, которые были двух- или трехзарядными. Результаты были загружены в Scaffold (версия 3.6.5, Proteome Software), и порог идентификации пептида и белка был установлен на 95% и 99% достоверности соответственно. Данные были экспортированы в виде отчета об образцах и проанализированы в Excel (Microsoft).Невзвешенное спектральное количество на белок сначала нормализовалось по общему спектру, а затем нормализовалось по молекулярной массе каждого белка.
Данные протеомики MS были депонированы в Консорциум ProteomeXchange [35] через репозиторий партнеров PRIDE с идентификатором набора данных PXD001183 и DOI 10.6019 / PXD001183.
Иерархическая кластеризация и анализ генной онтологии
Иерархическая кластеризация выполнялась с использованием Cluster 3. 0 (C Clustering Library, версия 1.37) и MultiExperiment Viewer (программный пакет TM4 Microarray). Нормализованные спектральные подсчеты были умножены на 1000 и сгруппированы с использованием нецентрированной корреляции Пирсона в качестве показателя расстояния и средней связи в качестве критериев связи. Списки белков с инвентарными номерами IPI были просмотрены с использованием инструмента функциональной аннотации в DAVID [36], [37]. Для построения круговых диаграмм для каждого термина онтологии гена был рассчитан −log 10 (значение p с поправкой Бонферрони).
0 (C Clustering Library, версия 1.37) и MultiExperiment Viewer (программный пакет TM4 Microarray). Нормализованные спектральные подсчеты были умножены на 1000 и сгруппированы с использованием нецентрированной корреляции Пирсона в качестве показателя расстояния и средней связи в качестве критериев связи. Списки белков с инвентарными номерами IPI были просмотрены с использованием инструмента функциональной аннотации в DAVID [36], [37]. Для построения круговых диаграмм для каждого термина онтологии гена был рассчитан −log 10 (значение p с поправкой Бонферрони).
Сеть белок-белкового взаимодействия
Cytoscape (версия 2.8.3) был использован для визуализации сетей белок-белковых взаимодействий из данных MS. Используемый человеческий интерактом был построен на основе платформы анализа сети взаимодействия белков Homo sapiens (дата выпуска 28 июня 2011 г.) и сети Mus musculus (дата выпуска 28 июня 2011 г.) [38], базы данных взаимодействий ECM MatrixDB ( дата выпуска, 26 августа 2010 г. ) [39], а также созданная в литературе база данных белков, связанных с адгезией на основе интегрина [3], [4]. Белковые узлы окрашивали в соответствии со значениями log 2 (нокодазол / ДМСО).Узлы были отсортированы на основе их клеточной локализации с использованием терминов генной онтологии «клеточного компонента» и проверены вручную, чтобы уменьшить расхождения в назначениях. Цитоплазматические узлы были дополнительно упорядочены в соответствии с расстоянием сети взаимодействия от интегринов плазматической мембраны, то есть 1-скачок, 2-шаг. Размер узла был скорректирован, чтобы отразить нормализованные по молекулярной массе спектральные числа.
) [39], а также созданная в литературе база данных белков, связанных с адгезией на основе интегрина [3], [4]. Белковые узлы окрашивали в соответствии со значениями log 2 (нокодазол / ДМСО).Узлы были отсортированы на основе их клеточной локализации с использованием терминов генной онтологии «клеточного компонента» и проверены вручную, чтобы уменьшить расхождения в назначениях. Цитоплазматические узлы были дополнительно упорядочены в соответствии с расстоянием сети взаимодействия от интегринов плазматической мембраны, то есть 1-скачок, 2-шаг. Размер узла был скорректирован, чтобы отразить нормализованные по молекулярной массе спектральные числа.
Эксперименты по иммунофлуоресценции
HFF не содержали сыворотки в течение 16 часов, а затем обрабатывали ДМСО или нокодазолом в течение 4 часов.Для визуализации микротрубочек клетки фиксировали 4% (мас. / Об.) Параформальдегидом с добавлением 0,05% (об. / Об.) Глутарового альдегида в течение 7 минут при комнатной температуре. Для всех других экспериментов по иммунофлуоресценции клетки фиксировали 4% (мас. / Об.) Параформальдегидом в течение 7 минут при комнатной температуре. Клетки пропускали через 0,2% (об. / Об.) Triton X-100 в течение 10 минут при комнатной температуре, блокировали 1% (мас. / Об.) Бычьим сывороточным альбумином (BSA) с добавлением 0,01% (v / v) Tween 20 (BSA -T) в течение 30 минут и инкубировали с первичным антителом, разведенным в BSA-T, в течение 1 часа при комнатной температуре.Клетки трижды промывали PBS —, инкубировали с вторичным антителом, разведенным в BSA-T в течение 1 часа при комнатной температуре, и трижды промывали PBS —. Иммуноокрашенные клетки визуализировали с помощью реставрационного микроскопа Delta Vision [RT] (Applied Precision) с использованием объектива [60x / 1,42 Plan Apo / Oil immersion] и набора фильтров Sedat Quad (Chroma [89000]). Изображения были собраны с помощью камеры Coolsnap HQ (Photometrics) в программном обеспечении Softworx (Applied Precision) в виде единого стека и проанализированы с помощью ImageJ (версия 1.
Для всех других экспериментов по иммунофлуоресценции клетки фиксировали 4% (мас. / Об.) Параформальдегидом в течение 7 минут при комнатной температуре. Клетки пропускали через 0,2% (об. / Об.) Triton X-100 в течение 10 минут при комнатной температуре, блокировали 1% (мас. / Об.) Бычьим сывороточным альбумином (BSA) с добавлением 0,01% (v / v) Tween 20 (BSA -T) в течение 30 минут и инкубировали с первичным антителом, разведенным в BSA-T, в течение 1 часа при комнатной температуре.Клетки трижды промывали PBS —, инкубировали с вторичным антителом, разведенным в BSA-T в течение 1 часа при комнатной температуре, и трижды промывали PBS —. Иммуноокрашенные клетки визуализировали с помощью реставрационного микроскопа Delta Vision [RT] (Applied Precision) с использованием объектива [60x / 1,42 Plan Apo / Oil immersion] и набора фильтров Sedat Quad (Chroma [89000]). Изображения были собраны с помощью камеры Coolsnap HQ (Photometrics) в программном обеспечении Softworx (Applied Precision) в виде единого стека и проанализированы с помощью ImageJ (версия 1. 48c) Национальные институты здравоохранения).
48c) Национальные институты здравоохранения).
Результаты
Методика выделения комплексов вентральной плазматической мембраны для характеристики AC при разрыве микротрубочек
Чтобы охарактеризовать глобальные изменения ACs при потере микротрубочек, мы адаптировали ранее описанные рабочие процессы, чтобы позволить специфическое выделение белковых комплексов вентральной мембраны для анализа с помощью протеомики на основе MS [32] — [34]. Комплексы вентральной плазматической мембраны из клеток, прикрепленных к фибронектину (FN) или контрольному лиганду, поли-D-лизину (PDL), были стабилизированы поперечным сшиванием, выделены после лизиса клеток с помощью обработки ультразвуком (рис.1) и проанализированы с помощью вестерн-блоттинга или протеомики на основе МС.
Первоначально голодные по сыворотке HFF, прикрепленные к FN, обрабатывали нокодазолом для разрушения сети микротрубочек, фиксировали и иммуноокрашивали на актин, винкулин и тубулин (рис. 2a).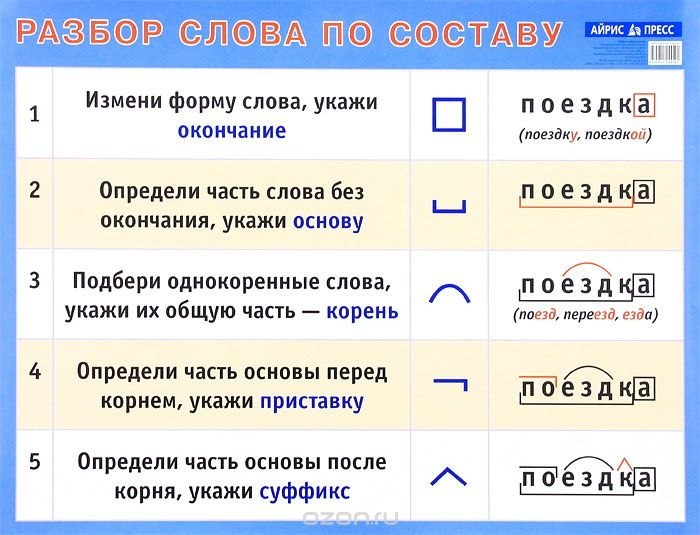 В соответствии с предыдущими исследованиями [8], [20], [24], обработанные носителем клетки с недостатком сыворотки демонстрировали удлиненную клеточную морфологию, хорошо сформированную сеть микротрубочек, редкий и дезорганизованный актиновый цитоскелет и небольшие периферические винкулинсодержащие AC .Напротив, клетки, обработанные нокодазолом, демонстрировали сокращенную морфологию с характерными «пальцеобразными» выступами на краях клеток, потерей сети микротрубочек, плотными и толстыми актиновыми стрессовыми волокнами и большими, содержащими винкулин ACs по всей клетке (рис. 2a). . Количественное определение винкулин-содержащих АК показало, что площадь клеток, обработанных нокодазолом, увеличилась в 3,3 раза по сравнению с клетками, обработанными носителем (фиг. 2b).
В соответствии с предыдущими исследованиями [8], [20], [24], обработанные носителем клетки с недостатком сыворотки демонстрировали удлиненную клеточную морфологию, хорошо сформированную сеть микротрубочек, редкий и дезорганизованный актиновый цитоскелет и небольшие периферические винкулинсодержащие AC .Напротив, клетки, обработанные нокодазолом, демонстрировали сокращенную морфологию с характерными «пальцеобразными» выступами на краях клеток, потерей сети микротрубочек, плотными и толстыми актиновыми стрессовыми волокнами и большими, содержащими винкулин ACs по всей клетке (рис. 2a). . Количественное определение винкулин-содержащих АК показало, что площадь клеток, обработанных нокодазолом, увеличилась в 3,3 раза по сравнению с клетками, обработанными носителем (фиг. 2b).
Рис. 2. Изменения актиновых стрессовых волокон и винкулин-содержащих AC при разрыве микротрубочек.
HFF с недостатком сыворотки обрабатывали ДМСО и нокодазолом (Noc) в течение 4 часов и фиксировали либо А) параформальдегидом с добавлением глутарового альдегида для сохранения микротрубочек, и иммуноокрашивали на актин (зеленый), винкулин (красный) и тубулин (синий) или Б) параформальдегид и иммуноокрашенный на винкулин. Окрашивание винкулина количественно определяли как процент от общей площади клеток и отображали в виде графика Beeswarm-Boxplot (линия, медиана; прямоугольник, межквартильный диапазон; усы, максимум и минимум; n = от 51 до 55 клеток).
Окрашивание винкулина количественно определяли как процент от общей площади клеток и отображали в виде графика Beeswarm-Boxplot (линия, медиана; прямоугольник, межквартильный диапазон; усы, максимум и минимум; n = от 51 до 55 клеток).
https://doi.org/10.1371/journal.pone.0115213.g002
Чтобы определить, могут ли быть обнаружены изменения AC после потери микротрубочек из изолированных комплексов вентральной плазматической мембраны, комплексы были выделены из клеток, прикрепленных к FN или PDL, после обработки носителем (ДМСО) или нокодазолом и проанализировали с помощью вестерн-блоттинга (фиг. 3). В соответствии с результатами иммунофлуоресценции, комплексы вентральной плазматической мембраны из обработанных нокодазолом клеток показали общее увеличение содержания компонентов AC по сравнению с клетками, обработанными носителем: 1.Увеличение винкулина в 5 раз, талина в 1,5 раза, паксиллина в 2,4 раза, FAK в 1,4 раза, FAK фосфотирозина-397 (pY397-FAK) в 2,4 раза и ILK в 3,8 раза (рис. 3). Примечательно, что содержание α 5 -интегрина не изменилось в клетках, обработанных нокодазолом, по сравнению с клетками, обработанными носителем (фиг. 3). Это общее увеличение количества компонентов AC согласуется с предыдущими исследованиями, которые показали сборку и рост фокальных адгезий при потере микротрубочек [20], [24], [25].В качестве контроля компоненты трансферина, не относящиеся к AC, и BAK не были обнаружены в комплексах вентральной плазматической мембраны из клеток, прикрепленных к FN (рис. 3a). Более того, компоненты AC не были обогащены комплексами вентральной плазматической мембраны, выделенными из клеток, прикрепленных к PDL (рис. 3a). Взятые вместе, эти результаты подтверждают этот метод специфического выделения AC для обнаружения количественных изменений белка при потере микротрубочек.
3). Примечательно, что содержание α 5 -интегрина не изменилось в клетках, обработанных нокодазолом, по сравнению с клетками, обработанными носителем (фиг. 3). Это общее увеличение количества компонентов AC согласуется с предыдущими исследованиями, которые показали сборку и рост фокальных адгезий при потере микротрубочек [20], [24], [25].В качестве контроля компоненты трансферина, не относящиеся к AC, и BAK не были обнаружены в комплексах вентральной плазматической мембраны из клеток, прикрепленных к FN (рис. 3a). Более того, компоненты AC не были обогащены комплексами вентральной плазматической мембраны, выделенными из клеток, прикрепленных к PDL (рис. 3a). Взятые вместе, эти результаты подтверждают этот метод специфического выделения AC для обнаружения количественных изменений белка при потере микротрубочек.
Рисунок 3. Изменения в изолированных вентральных комплексах плазматической мембраны при разрыве микротрубочек.
A) HFF с недостатком сыворотки, нанесенные на фибронектин (FN) или поли-D-лизин (PDL), обрабатывали DMSO или нокодазолом (Noc) в течение 4 часов, и комплексы вентральной плазматической мембраны выделяли для вестерн-блоттинга компонентов AC. (α 5 -интегрин, талин, винкулин, паксиллин, FAK, pY397-FAK и ILK) и компоненты, не относящиеся к AC (BAK и рецептор трансферрина). B) Полосы белка были количественно определены и нормализованы к FN, DMSO (среднее ± стандартное отклонение; n = 3).
(α 5 -интегрин, талин, винкулин, паксиллин, FAK, pY397-FAK и ILK) и компоненты, не относящиеся к AC (BAK и рецептор трансферрина). B) Полосы белка были количественно определены и нормализованы к FN, DMSO (среднее ± стандартное отклонение; n = 3).
https: // doi.org / 10.1371 / journal.pone.0115213.g003
Количественный МС-анализ изолированных комплексов вентральной плазматической мембраны при разрыве микротрубочек
Чтобы проанализировать изменения AC при потере микротрубочек глобальным, объективным образом, комплексы вентральной плазматической мембраны из клеток, обработанных носителем и нокодазолом, были проанализированы с помощью количественной масс-спектрометрии с использованием спектрального подсчета в качестве меры относительного содержания белка. MS идентифицировала 850–1000 белков в каждом образце (таблица S1). Для выявления закономерностей относительного содержания различных белков в каждом состоянии была проведена иерархическая кластеризация (рис. 4). Были идентифицированы три основных кластера: (i) белки с одинаковым содержанием в образцах как FN, так и PDL, (ii) белки, обогащенные PDL, и (iii) белки, обогащенные FN. Анализ пути KEGG показал, что большинство значительно обогащенных функциональных терминов (скорректированное Бонферрони p -значение <0,05) для белков, обогащенных FN (кластер iii), были связаны с AC (т.е. очаговая адгезия, взаимодействие ECM-рецептора, регуляция актина цитоскелет и трансэндотелиальная миграция лейкоцитов) (таблица S2).Другие термины, обогащенные FN (кластер iii), были связаны с заболеваниями, которые, как было показано, связаны с дисфункциональными адгезионными комплексами, например, аритмогенная кардиомиопатия правого желудочка, гипертрофическая кардиомиопатия, дилатационная кардиомиопатия и мелкоклеточный рак легкого [40]. Напротив, наиболее значительно обогащенные термины для белков с одинаковым содержанием в FN и PDL (кластер i) и белков, обогащенных PDL (кластер ii), не были связаны с AC (т.
4). Были идентифицированы три основных кластера: (i) белки с одинаковым содержанием в образцах как FN, так и PDL, (ii) белки, обогащенные PDL, и (iii) белки, обогащенные FN. Анализ пути KEGG показал, что большинство значительно обогащенных функциональных терминов (скорректированное Бонферрони p -значение <0,05) для белков, обогащенных FN (кластер iii), были связаны с AC (т.е. очаговая адгезия, взаимодействие ECM-рецептора, регуляция актина цитоскелет и трансэндотелиальная миграция лейкоцитов) (таблица S2).Другие термины, обогащенные FN (кластер iii), были связаны с заболеваниями, которые, как было показано, связаны с дисфункциональными адгезионными комплексами, например, аритмогенная кардиомиопатия правого желудочка, гипертрофическая кардиомиопатия, дилатационная кардиомиопатия и мелкоклеточный рак легкого [40]. Напротив, наиболее значительно обогащенные термины для белков с одинаковым содержанием в FN и PDL (кластер i) и белков, обогащенных PDL (кластер ii), не были связаны с AC (т. е. рибосома и протеасома). Следовательно, иерархическая кластеризация подтвердила глобальное обогащение AC-родственных белков клетками, распространяющимися по FN.
е. рибосома и протеасома). Следовательно, иерархическая кластеризация подтвердила глобальное обогащение AC-родственных белков клетками, распространяющимися по FN.
Рисунок 4. Неконтролируемый иерархический анализ кластеризации вентральных комплексов плазматической мембраны при разрыве микротрубочек.
Полный вывод неконтролируемого иерархического кластерного анализа белков, идентифицированных с помощью МС. Указываются корреляции в выбранных узлах дендрограммы. Были идентифицированы три кластера: i) белки, присутствующие в равном количестве в образцах как фибронектина (FN), так и поли-D-лизина (PDL); ii) белки, обогащенные PDL; и iii) белки, обогащенные FN. Кластеры были подвергнуты анализу обогащения путей и значительно обогатили термины пути KEGG (скорректированное Бонферрони p -значение <0.05) отображаются в виде круговых диаграмм.
https://doi.org/10.1371/journal.pone.0115213.g004
Для дальнейшего уточнения набора данных, QSpec [41] использовался для идентификации белков, которые были статистически обогащены FN по сравнению с PDL. QSpec — это статистическая структура для данных спектрального подсчета, которая дает статистическую меру белков, которые по-разному экспрессируются в двух условиях. Всего 196 белков были статистически обогащены до FN по сравнению с PDL, из которых 26 белков были обнаружены в адгезомах, описанных в литературе [3] (рис.5а). В соответствии с данными иерархической кластеризации на рис. 4 кластера (iii), наиболее значительно обогащенные термины для 196 белков были связаны с AC (рис. 5b и таблица S2).
QSpec — это статистическая структура для данных спектрального подсчета, которая дает статистическую меру белков, которые по-разному экспрессируются в двух условиях. Всего 196 белков были статистически обогащены до FN по сравнению с PDL, из которых 26 белков были обнаружены в адгезомах, описанных в литературе [3] (рис.5а). В соответствии с данными иерархической кластеризации на рис. 4 кластера (iii), наиболее значительно обогащенные термины для 196 белков были связаны с AC (рис. 5b и таблица S2).
Рисунок 5. Идентификация статистически обогащенных белков адгезионного комплекса.
График вулкана выходных данных QSpec, log 10 (байесовский фактор) и ln (кратное изменение, FN / PDL). Горизонтальная пунктирная линия соответствует байесовскому фактору 10, а вертикальная пунктирная линия соответствует кратному изменению (FN / PDL), равному 3.Белки, статистически обогащенные FN, находятся в верхнем правом квадранте (log 10 (байесовский фактор) ≥1 и ln (кратное изменение, FN / PDL) ≥1,1), что представляет собой консервативную оценку вероятности ложного обнаружения менее 5%. [41]. Белки, статистически обогащенные FN, были подвергнуты анализу обогащения пути, и значительно обогащенные параметры пути KEGG (скорректированное Бонферрони p — значение <0,05) отображены в виде круговой диаграммы. ФН, фибронектин; PDL, поли-D-лизин.
[41]. Белки, статистически обогащенные FN, были подвергнуты анализу обогащения пути, и значительно обогащенные параметры пути KEGG (скорректированное Бонферрони p — значение <0,05) отображены в виде круговой диаграммы. ФН, фибронектин; PDL, поли-D-лизин.
https: // doi.org / 10.1371 / journal.pone.0115213.g005
Для оценки относительных изменений белков, статистически обогащенных до FN, при потере микротрубочек, кратное изменение нормализованных спектральных количеств компонентов AC, идентифицированных из клеток, обработанных нокодазолом, по сравнению с таковыми из клеток из рассчитывали клетки, обработанные носителем. Иерархическая кластеризация использовалась для обзора относительных изменений белков (рис. 6). При потере микротрубочек наблюдались три кластера: (i) белки, численность которых уменьшилась, (ii) белки, численность которых не изменилась, и (iii) белки, численность которых увеличилась.Обогащенный анализ генной онтологии этих кластеров с использованием терминов «клеточный компонент» показал, что большинство значительно расширенных терминов для кластеров (i) и (ii) были связаны с ECM. Напротив, большинство значительно обогащенных терминов для кластера (iii) было связано с клеточной адгезией, межклеточными соединениями и связанными структурами (рис. 6 и таблица S2). Все белки AC, которые были обогащены FN по сравнению с условиями PDL, как было идентифицировано с помощью вестерн-блоттинга, были обнаружены в кластере (iii) (фиг. 3).
Напротив, большинство значительно обогащенных терминов для кластера (iii) было связано с клеточной адгезией, межклеточными соединениями и связанными структурами (рис. 6 и таблица S2). Все белки AC, которые были обогащены FN по сравнению с условиями PDL, как было идентифицировано с помощью вестерн-блоттинга, были обнаружены в кластере (iii) (фиг. 3).
Рисунок 6. Неконтролируемый иерархический кластерный анализ белков, статистически обогащенных до FN.
Дендограмма тепловой карты, отображающая иерархическую кластеризацию кратного изменения FN, Noc относительно FN, DMSO. Три кластера были идентифицированы и проанализированы с помощью анализа обогащения генной онтологии для выявления значительно обогащенных терминов «клеточный компонент» (скорректированный по Бонферрони p — значение <0,05). В общем, тремя кластерами были: i) белки, количество которых уменьшилось при лечении нокодазолом и было обогащено в терминах, связанных с ECM; ii) белки, которые не изменились и были обогащены в терминах, связанных с ECM; и iii) белки, которые увеличились и были обогащены в терминах, связанных с AC. ФН, фибронектин; PDL, поли-D-лизин; Нет, нокодазол.
ФН, фибронектин; PDL, поли-D-лизин; Нет, нокодазол.
https://doi.org/10.1371/journal.pone.0115213.g006
Таким образом, мы выделили AC при разрыве микротрубочек и выполнили MS и кластерный анализ для идентификации белков адгезии, регулируемых микротрубочками. Эти находки демонстрируют, что разрушение микротрубочек приводит к общему увеличению количества компонентов AC, тогда как белки ECM остаются такими же или уменьшаются при разрыве микротрубочек.
Сетевой анализ белок-белкового взаимодействия
Для дальнейшего изучения биологической функции вызванных микротрубочками изменений AC, белки, статистически обогащенные до FN, были картированы в сети белок-белковых взаимодействий (PPI) и расположены в соответствии с их клеточной локализацией (внеклеточная, плазматическая мембрана, цитоплазматическая, органелла и ядро). ) (Рис.7). Чтобы уменьшить сложность взаимодействий между цитоплазматическими белками, сеть была упорядочена в соответствии с расстоянием сети взаимодействия относительно интегринов, интеркалированных плазматической мембраной. Белковые узлы окрашивались в соответствии с изменением их содержания в нокодазоле по сравнению с условиями, обработанными носителем (красный цвет указывает на увеличение, а синий — уменьшение содержания белка после лечения нокодазолом). После лечения нокодазолом большинство белков в ЕСМ уменьшилось, тогда как белки плазматической мембраны и цитоплазмы увеличились (рис. 7). Хотя большинство интегринов плазматической мембраны увеличивалось при разрыве микротрубочек, основные связывающие FN интегрины α 5 и β 1 оставались относительно неизменными, что согласуется с результатами вестерн-блоттинга (рис.3).
Белковые узлы окрашивались в соответствии с изменением их содержания в нокодазоле по сравнению с условиями, обработанными носителем (красный цвет указывает на увеличение, а синий — уменьшение содержания белка после лечения нокодазолом). После лечения нокодазолом большинство белков в ЕСМ уменьшилось, тогда как белки плазматической мембраны и цитоплазмы увеличились (рис. 7). Хотя большинство интегринов плазматической мембраны увеличивалось при разрыве микротрубочек, основные связывающие FN интегрины α 5 и β 1 оставались относительно неизменными, что согласуется с результатами вестерн-блоттинга (рис.3).
Рисунок 7. Анализ сети белок-белкового взаимодействия комплексов вентральной плазматической мембраны, обогащенных FN.
Цвет узла определяли по сине-красному цветовому градиенту, соответствующему логарифмическим значениям 2 (Noc / DMSO), с красными узлами, указывающими на увеличение, а синими узлами — на уменьшение содержания белка после обработки нокодазолом. Узлы были отсортированы в соответствии с их местоположением в ячейках, и размер узла был пропорционален нормализованным спектральным счетчикам. Сеть была упорядочена в соответствии с расстоянием сети взаимодействия относительно интегринов плазматической мембраны, 1-скачок, 2-скачок и т. Д.Соответствующие названия генов отображаются под каждым узлом для ясности. ФН, фибронектин; Нет, нокодазол.
https://doi.org/10.1371/journal.pone.0115213.g007
Для подтверждения микротрубочек-зависимых изменений в составе AC, наблюдаемых с помощью MS, AC были выделены из клеток, обработанных носителем и нокодазолом, и подмножества белков. идентифицированы из анализа MS, оцененного с помощью вестерн-блоттинга (фиг. 8a). В соответствии с результатами МС, α против -интегрина, тензина-1 и филамина A показали значительное увеличение численности при лечении нокодазолом; белки ELKS (ERC1) и PDLIM5 показали незначительное увеличение.Кроме того, количественная оценка площади клетки, содержащей α 5 -интегрин-, α v β 3 -интегрин- и винкулин-содержащие AC, с использованием иммуноокрашивания показала, что не было значительных изменений α 5 — площадь интегрина после обработки нокодазолом, тогда как площадь α v β 3 -интегрина и винкулина увеличивалась приблизительно в 5,5 и 2,0 раза соответственно после обработки нокодазолом (фиг. 8b).
Рисунок 8. Подтверждение изменений в компонентах переменного тока, наблюдаемых МС.
A) Выделенные AC анализировали вестерн-блоттингом на α v -интегрин, тензин-1, филамин A, ELKS и PDLIM5. B) Для дальнейшего изучения характера изменений интегринов при потере микротрубочек HFF, обработанные ДМСО или нокодазолом (Noc), фиксировали и иммуноокрашивали на α 5 -интегрин, α v β 3 -интегрин и винкулин и определяется как процент от общей площади ячейки (линия, медиана; прямоугольник, межквартильный размах; усы, максимум и минимум; **** p <0.0001, Студенческий т -тест; n = 54). C) Иммуноокрашенные клетки, показывающие клеточное распределение α 5 -интегрина (красный), α v β 3 -интегрина (зеленый) и винкулина (синий). Синие и желтые вставки — это 4-кратное увеличение соответствующих коробок.
https://doi.org/10.1371/journal.pone.0115213.g008
Взятые вместе, эти результаты согласуются с предыдущими исследованиями, которые показали сборку и рост фокальных спаек при потере микротрубочек [20], [24 ], [25].Кроме того, данные расширяют эти исследования, характеризуя изменения ACs глобальным, непредвзятым образом и предполагая модуляцию компонентов AC, которая не зависит от рецептора интегрина.
Изменения состояния созревания АЦ при разрыве микротрубочек
В клетках, обработанных носителем, α 5 -интегрин, α v β 3 -интегрин и винкулин были обнаружены на периферии клеток в крупных очаговых адгезиях, но α 5 -интегрин также был обнаружен в центрально расположенных полосообразные структуры, которые напоминают фибриллярные спайки и другие точечные структуры AC.В клетках, обработанных нокодазолом, наблюдалось усиление окрашивания α v β 3 -интегрина на периферии клеток в крупных фокальных адгезиях, а α 5 -интегрин и винкулин также были обнаружены в центрально расположенных фокальных адгезионных связях. структуры (рис. 8в). Эти результаты предполагают, что нарушение микротрубочек вызывает переключение в состоянии созревания AC в сторону фокальных адгезий, так что α 5 -интегрин перераспределяется в фокальные адгезии и сопровождается увеличением количества компонентов AC.
Фибриллярные адгезии участвуют в фибриллогенезе ФН [42], [43] и обычно характеризуются высокими уровнями α 5 β 1 -интегрина и тензина, низкими уровнями паксиллина, винкулина и pTyr [19] , [21] и отсутствие прикрепления к актиновым стрессовым волокнам [19], [44]. Чтобы определить, являются ли адгезионные структуры на фиг.8 фибриллярными адгезиями и исследовать их модуляцию при потере микротрубочек, клетки, обработанные носителем и нокодазолом, иммуноокрашивали на тензин-1, pTyr (фиг.9) и актиновые стресс-волокна (рис. 10). В клетках, обработанных носителем, тензин-1 локализован совместно с α 5 -интегрином по всей клетке, включая центрально расположенные полосообразные структуры α 5 -интегрина, которые отображали низкие уровни винкулина (рис. 9a), тогда как pTyr и толстые актиновые стрессовые волокна совместно локализованы с периферическими АЦ α 5 -интегрина, которые имеют высокие уровни винкулина (рис. 9b и 10). Следовательно, эти анализы показывают, что наблюдаемые центрально расположенные полосообразные структуры α 5 -интегрина были фибриллярными адгезиями.
Рис. 9. Изменения состояния созревания AC при разрыве микротрубочек.
HFF с недостатком сыворотки обрабатывали ДМСО и нокодазолом (Noc) в течение 4 часов, фиксировали и иммуноокрашивали на A) α 5 -интегрин (красный), тензин-1 (зеленый) и винкулин (синий) и B) α 5 -интегрин (красный), винкулин (зеленый) и pTyr (синий). Синие и желтые вставки — это 4-кратное увеличение соответствующих коробок.
https://doi.org/10.1371/journal.pone.0115213.g009
Рисунок 10.Силовая зависимость модуляции переменного тока, индуцированной микротрубочками.
HFF с недостатком сыворотки обрабатывали ДМСО, Y-27632, нокодазолом (Noc) или комбинацией Noc и Y-27632, фиксировали и иммуноокрашивали на α 5 -интегрин (красный), винкулин (зеленый) и актин ( синий). Синие и желтые вставки — это 4-кратное увеличение соответствующих коробок.
https://doi.org/10.1371/journal.pone.0115213.g010
В клетках, обработанных нокодазолом, α 5 -интегрин, тензин-1 и винкулин совместно локализованы по всей клетке в периферических и центральных очагах спайки (рис.9а). Кроме того, pTyr и толстые актиновые стрессовые волокна совместно локализованы с периферическими и центральными α 5 -интегриновыми структурами, которые имеют высокие уровни винкулина (фиг. 9b и 10). Следовательно, эти результаты демонстрируют, что потеря микротрубочек приводит к увеличению фокальных адгезий, особенно в центральных областях клетки, и уменьшению фибриллярных адгезий и других структур AC. Взятые вместе, эти результаты дополнительно подтверждают, что разрушение микротрубочек вызывает изменение в состоянии созревания ACs, чтобы способствовать образованию фокальных адгезий из др. Структур α 5 -integrin AC.
Определение силовой зависимости модуляции переменного тока, индуцированной микротрубочками
Потеря клеточных микротрубочек сопровождается повышенной сократимостью актомиозина из-за высвобождения и активации локализованного в микротрубочках GEF-h2 и последующей стимуляции RhoA [22], [23]. Следовательно, чтобы определить, было ли перераспределение α 5 -интегрина и изменение состояния созревания AC обусловлено микротрубочко-зависимыми изменениями клеточной силы, был использован Y-27632, ингибитор Rho-ассоциированной протеинкиназы (ROCK). (Рисунок.10). Клетки, обработанные Y-27632, демонстрировали разреженные актиновые стрессовые волокна, небольшие периферические винкулин-содержащие фокальные комплексы и центрально расположенные фибриллярные адгезии α 5 -интегрина. Напротив, клетки, обработанные нокодазолом, демонстрировали толстые актиновые стрессовые волокна, большие центральные и периферические фокальные спайки, содержащие винкулин. Совместная обработка клеток Y-27632 и нокодазолом обращала эффекты нокодазола, поскольку клетки проявляли разреженные актиновые стрессовые волокна, небольшие периферические винкулин-содержащие фокальные комплексы и центрально расположенные фибриллярные адгезии α 5 -интегрина.Эти данные указывают на то, что перераспределение α 5 -интегрина и изменение состояния созревания AC до фокальных спаек при разрыве микротрубочек зависит от силы и опосредуется посредством пути, опосредованного RhoA-ROCK. Таким образом, эти результаты демонстрируют сложное взаимодействие между динамикой микротрубочек, генерацией силы и состоянием созревания AC.
Discussion
Сеть микротрубочек играет важную роль в регуляции ACs и предыдущие исследования показали, что разрушение микротрубочек приводит к фокальной адгезии и образованию актиновых стрессовых волокон [20], [24], [25], [45].Чтобы охарактеризовать взаимодействие между микротрубочками и ACs, мы использовали стратегию выделения вентральной плазматической мембраны и количественный подход протеомики на основе MS для определения изменений состава AC при разрыве микротрубочек глобальным, непредвзятым образом. Мы представляем новый набор данных MS компонент AC, который описывает изменения AC при разрыве микротрубочек. Нашим основным открытием является то, что после потери микротрубочек, большинство компонентов AC увеличилось в количестве в местах формирования фокальной адгезии, но количество α 5 — и β 1 -интегринов не изменилось.Дальнейшие исследования показали, что это происходит из-за зависимого от внутриклеточной силы переключения в состоянии созревания AC в сторону фокальных адгезий, так что α 5 -интегрин перераспределяется в фокальные адгезии и сопровождается увеличением количества компонентов AC. Таким образом, этот набор данных MS каталогизирует модуляцию состава AC при увеличении клеточной силы после разрушения микротрубочек и дополняет предыдущие исследования, которые определили зависимые от силы изменения состава AC при ингибировании миозина II блеббистатином [33], [34], [33], [34], [ 46].Эти данные подчеркивают важность микротрубочек в регуляции внутриклеточных сил для определения различных состояний ACs и предоставляют ресурсы для будущих исследований передачи сигналов адгезии.
В базальном состоянии покоя клетки существует континуум состояний AC, который включает возникающие адгезии, фокальные комплексы, фокальные адгезии и фибриллярные адгезии [47], [48]. Эти разные состояния переменного тока не только различаются по размеру и положению, но также по составу и функциям [21], [42], [47], [48].Это исследование предоставляет несколько линий доказательств, подтверждающих переключение состояний AC, вызванное разрывом микротрубочек, в пользу крупных фокальных спаек. В частности, 1) содержание α 5 — и β 1 -интегринов не изменилось; 2) большинство компонентов AC, особенно структурные белки, были увеличены; 3) образование многочисленных крупных очаговых спаек на периферии и даже в центральных областях клеток; и 4) уменьшение других структур AC, таких как фибриллярные адгезии.
Переключение в состояние AC в пользу фокальных адгезий может быть связано с высвобождением и активацией связанного с микротрубочками GEF-h2 и последующей активацией RhoA [22], [23]. RhoA действует на два разных, но сопутствующих пути, которые участвуют в созревании фокальной адгезии: mDia и ROCK [49], [50]. Активация пути RhoA-mDia приводит к образованию стрессовых волокон [51], [52], тогда как путь RhoA-ROCK способствует опосредованной миозином II сократимости клеток [53]. Предыдущие исследования показали участие сократимости клеток в формировании фокальных спаек при разрыве микротрубочек [20].Чтобы продвинуть эти исследования, мы показали, что повышенная сократимость клеток при разрыве микротрубочек происходит преимущественно через путь RhoA-ROCK-миозин II, поскольку специфическое ингибирование ROCK предотвращает образование фокальной адгезии при разрыве микротрубочек. Следовательно, AC реагируют на повышение внутриклеточного давления, усиливая AC с помощью чувствительных к напряжению белков, таких как винкулин и талин. Действительно, было показано, что при лечении нокодазолом AC удлиняются центростремительно, причем винкулин обогащается на дистальных концах AC, ближайших к актиновым стрессовым волокнам [25].В соответствии с этими находками мы обнаружили общее увеличение количества структурных белков AC, которые связывают интегрины с актиновым цитоскелетом.
В нашей сети PPI мы обнаружили много структурных белков AC, которые были тесно связаны и близки по расстоянию сети взаимодействия от интегринов плазматической мембраны. Было высказано предположение, что важность белкового узла для поддержания сети взаимодействия белков связана с плотностью его взаимодействий [3]. Анализ PPI показал, что белки с наибольшим количеством взаимодействий в цитоплазме, как сообщалось, более напрямую взаимодействуют с интегринами плазматической мембраны (1-прыжок и 2-прыжок от интегринов).Примечательно, что большинство сильно взаимосвязанных (≥3 взаимодействий) цитоплазматических белков показало увеличение количества при потере микротрубочек. Взятые вместе, эти результаты предполагают, что механочувствительный модуль, участвующий в определении природы состояний AC при разрыве микротрубочек, скорее всего, состоит из структурных белков, непосредственно или тесно связанных с интегринами.
Утрата фибриллярных адгезий совпала с образованием крупных фокальных адгезий в центральных областях клетки при разрыве микротрубочек, тем самым предполагая переключение от фибриллярных адгезий к фокальным адгезиям.Это удивительно, потому что обычно считается, что созревание AC происходит однонаправленным образом, от возникающих адгезий к фокальным комплексам и фокальным спайкам, а затем и к поздним стадиям фибриллярных адгезий. Во время образования фибриллярных адгезий α 5 β 1 -интегрин, связанный с деформируемым матриксом FN, перемещается центростремительно [44]; следовательно, передача актомиозиновой сократимости матриксу FN через α 5 β 1 -integrin позволяет происходить ранним событиям фибриллогенеза FN [42].Фибриллярные адгезии представляют собой структуры с низким натяжением [19], и ингибирование сократимости актомиозина не влияет на наличие фибриллярных адгезий, но предотвращает образование фибрилл FN [19], [42]. Интересно, что ингибирование податливости матрицы FN путем сшивания ингибирует образование фибриллярных адгезий и фибрилл FN и способствует образованию фокальных адгези-подобных комплексов, содержащих α 5 β 1 -интегрин, винкулин и паксиллин [44 ]. Это открытие означает, что когда силы актомиозина локализованы в ACs, задействуются чувствительные к натяжению модули.Следовательно, преобразование фибриллярных адгезий в фокальные адгезии при разрыве микротрубочек может быть связано с увеличением локализованных сил актомиозина, испытываемых комплексами α 5 β 1 -интегрин и последующим привлечением чувствительных к натяжению модулей. Взятые вместе, эти находки предполагают, что состояние созревания ACs может происходить двунаправленно и может позволить клетке динамически регулировать ответ на внутриклеточные или внеклеточные силы.
Предыдущие исследования с использованием протеомики на основе MS для определения изменений состава ACs были сосредоточены на глобальных изменениях адгезомы при ингибировании миозина II с использованием блеббистатина [33], [34].Наши результаты дополняют эти исследования, потому что потеря микротрубочек сопровождается увеличением стимулируемой миозином II сократимости актина. Сообщалось, что снижение количества белков, содержащих домен LIM, является основным последствием лечения блеббистатином [33]. Аналогичным образом, мы показываем, что количество белков, содержащих домен LIM, увеличивается при лечении нокодазолом. Кроме того, недавнее исследование Schiller et al. [46] очертил различные роли интегринов класса β 1 и α v в восприятии жесткости и показал, что фокальные адгезии β 3 -интегрина представляют собой структуры, зависящие от натяжения, которые задействуют GEF-h2 и стимулируют образование толстых актиновых стрессовых волокон, возможно, через путь RhoA-mDia.Действительно, мы обнаружили, что разрушение микротрубочек вызывает увеличение фокальных адгезий α v β 3 -интегрин, прикрепленных к толстым актиновым стрессовым волокнам, что может быть связано с комбинацией повышенной сократимости клеток и повышенной доступности GEF-h2 для связываются с β 3 -интегриновыми хвостами. Кроме того, анализ набора данных MS идентифицировал кластер белков, состоящий из ELKS, липрина α1 и липрина β1, который был обогащен до AC при разрыве микротрубочек. Недавно было показано, что ELKS и липрин, наряду с LL5β, KANK1 и KIF21A, образуют комплексы прикрепления кортикальных микротрубочек, которые прикрепляют микротрубочки к плазматической мембране через CLASP [54], [55] и направляют микротрубочки к участкам вокруг AC, чтобы облегчить их разборка, а также деградация ЭБУ [30].В соответствии с этими исследованиями, мы показываем, что ELKS, липрин α1 и липрин β1 обогащаются изолированными AC после лечения нокодазолом. Интересно, что как LL5β, так и ELKS не претерпевают значительных изменений при лечении блеббистатином [33], подтверждая микротрубочко-зависимую, но не силозависимую регуляцию локализации этого кластера белков. Взятые вместе, эти находки показывают, что этот набор данных MS является полезным ресурсом, который описывает чувствительные к натяжению и зависимые от микротрубочек изменения ACs после разрушения микротрубочек.
Таким образом, мы использовали протеомику на основе MS для определения изменений состава AC при потере микротрубочек беспристрастным, глобальным образом. Этот набор данных важен для исследования сложной взаимосвязи между микротрубочками, AC, внутриклеточной сократительной силой и ECM. Кроме того, набор данных дополняет ранее опубликованные наборы данных [33], [34], [46] и предоставляет платформу для генерации гипотез и будущей работы по изучению взаимодействия между микротрубочками и ACs.
Благодарности
Мы благодарим Дж. А. Аскари (Манчестерский университет) за критическое прочтение рукописи; А. Кариси и К. Баллестрем (Манчестерский университет) за техническую помощь и обсуждения; Дж. Котту (Манчестерский университет) за помощь в микроскопии; С. Варвуду и Д. Найту (Манчестерский университет) за получение данных МС; и Дж. Н. Селли (Университет Манчестера) за биоинформатическую поддержку. Данные протеомики MS были депонированы в Консорциум ProteomeXchange [35] через репозиторий партнеров PRIDE с идентификатором набора данных PXD001183 и DOI 10.6019 / PXD001183.
Вклад авторов
Задумал и спроектировал эксперименты: DN JDH AB AMF MJH. Проведены эксперименты: DN. Проанализированы данные: DN JDH AB. Предоставленные реагенты / материалы / инструменты анализа: DN AMF AB. Написал статью: DN JDH AB MJH.
Ссылки
- 1. Хайнс Р.О. (2002) Интегрины: двунаправленные аллостерические сигнальные машины. Cell 110: 673–687.
- 2. Гейгер Б., Спатц Дж. П., Бершадский А. Д. (2009) Зондирование окружающей среды через очаговые спайки.Nature Reviews Молекулярная клеточная биология 10: 21–33.
- 3. Зайдель-Бар Р., Ицковиц С., Мааян А., Айенгар Р., Гейгер Б. (2007) Функциональный атлас адгезомы интегрина. Природа клеточной биологии 9: 858–868.
- 4. Зайдель-Бар Р., Гейгер Б. (2010) Переключаемая адгезия интегрина. Журнал клеточной науки 123: 1385–1388.
- 5. Ghatak S, Morgner J, Wickstroem SA (2013) ILK: псевдокиназа с уникальной функцией в связи интегрин-актин. Протоколы биохимического общества 41: 995–1001.
- 6. Hemmings L, Rees DJG, Ohanian V, Bolton SJ, Gilmore AP и др. (1996) Талин содержит три сайта связывания актина, каждый из которых примыкает к сайту связывания винкулина. Журнал клеточной науки 109: 2715–2726.
- 7. Дубаш А.Д., Веннерберг К., Гарка-Мата Р., Менольд М.М., Артур В.Т. и др. (2007) Новая роль Lsc / p115 RhoGEF и LARG в регуляции активности RhoA ниже адгезии к фибронектину. Журнал клеточной науки 120: 3989–3998.
- 8.Ren XD, Kiosses WB, Schwartz MA (1999) Регулирование малого GTP-связывающего белка Rho посредством клеточной адгезии и цитоскелета. Embo Journal 18: 578–585.
- 9. Guan JL (1997) Роль киназы фокальной адгезии в передаче сигналов интегрина. Международный журнал биохимии и клеточной биологии 29: 1085–1096.
- 10. Савада Ю., Тамада М., Дубин-Талер Б.Дж., Чернявская О., Сакаи Р. и др. (2006) Определение силы путем механического удлинения субстрата киназы семейства Src p130Cas.Ячейка 127: 1015–1026.
- 11. Friedland JC, Lee MH, Boettiger D (2009) Механически активируемый интегринный переключатель управляет функцией альфа (5) бета (1). Наука 323: 642–644.
- 12. дель Рио А., Перес-Хименес Р., Лю Р., Рока-Кусакс П., Фернандес Дж. М. и др. (2009) Растяжение отдельных молекул стержня талина активирует связывание винкулина. Наука 323: 638–641.
- 13. Гиллуи С., Сваминатан В., Гарсия-Мата Р., О’Брайен Е. Т., Суперфайн Р. и др. (2011) Rho GEFs LARG и GEF-h2 регулируют механический ответ на силу, действующую на интегрины.Природа клеточной биологии 13: 722–727.
- 14. Чой К.К., Висенте-Мансанарес М., Зарено Дж., Уитмор Л.А., Могилнер А. и др. (2008) Актин и альфа-актинин организуют сборку и созревание возникающих спаек моторно-независимым образом от миозина II. Природа клеточной биологии 10: 1039–1050.
- 15. Эвен-Рам С., Дойл А.Д., Конти М.А., Мацумото К., Адельштейн Р.С. и др. (2007) Myosin IIA регулирует подвижность клеток и перекрестное взаимодействие микротрубочек с актомиозином. Природа клеточной биологии 9: 299–309.
- 16. Ridley AJ, Hall A (1992) Небольшой GTP-связывающий белок rho регулирует сборку фокальных адгезий и актиновых стрессовых волокон в ответ на факторы роста. Cell 70: 389–399.
- 17. Pasapera AM, Schneider IC, Rericha E, Schlaepfer DD, Waterman CM (2010) Активность миозина II регулирует рекрутирование винкулина в очаговые спайки посредством FAK-опосредованного фосфорилирования паксиллина. Журнал клеточной биологии 188: 877–890.
- 18. Кариси А., Цанг Р., Грейнер А.М., Нидженхейс Н., Хит Н. и др.(2013) Винкулин регулирует набор и высвобождение основных белков фокальной адгезии силозависимым образом. Текущая биология 23: 271–281.
- 19. Замир Э., Кац М., Позен Й., Эрез Н., Ямада К.М. и др. (2000) Динамика и сегрегация адгезий клеточного матрикса в культивируемых фибробластах. Природа клеточной биологии 2: 191–196.
- 20. Бершадский А., Чаусовский А., Беккер Е., Любимова А., Гейгер Б. (1996) Участие микротрубочек в контроле адгезивно-зависимой передачи сигнала.Текущая биология 6: 1279–1289.
- 21. Замир Э., Кац Б.З., Аота С., Ямада К.М., Гейгер Б. и др. (1999) Молекулярное разнообразие адгезий клеточного матрикса. Журнал клеточной науки 112: 1655–1669.
- 22. Krendel M, Zenke FT, Bokoch GM (2002) Фактор обмена нуклеотидов GEF-h2 опосредует перекрестный обмен между микротрубочками и актиновым цитоскелетом. Природа клеточной биологии 4: 294–301.
- 23. Chang Y-C, Nalbant P, Birkenfeld J, Chang Z-F, Bokoch GM (2008) GEF-h2 связывает вызванную нокодазолом разборку микротрубочек с клеточной сократимостью через RhoA.Молекулярная биология клетки 19: 2147–2153.
- 24. Liu BP, Chrzanowska-Wodnicka M, Burridge K (1998) Деполимеризация микротрубочек индуцирует стрессовые волокна, фокальные адгезии и синтез ДНК через GTP-связывающий белок rho. Адгезия клеток и коммуникация 5: 249–255.
- 25. Kirchner J, Kam Z, Tzur G, Bershadsky AD, Geiger B (2003) Мониторинг фосфорилирования тирозина в фокальных адгезиях после разрушения микротрубочек живыми клетками. Журнал клеточной науки 116: 975–986.
- 26. Waterman-Storer CM, Worthylake RA, Liu BP, Burridge K, Salmon ED (1999) Рост микротрубочек активирует Rac1, способствуя обилию ламеллиподий в фибробластах. Природа клеточной биологии 1: 45–50.
- 27. Руни С., Уайт Дж., Назгевич А., Вудкок С.А., Андерсон К.И. и др. (2010) Rac-активатор STEF (Tiam2) регулирует миграцию клеток посредством разборки фокальной адгезии, опосредованной микротрубочками. Embo Reports 11: 292–298.
- 28. Каверина И., Крылышкина О., Смолл Ю. В. (1999) Нацеливание микротрубочек на контакты субстрата способствует их релаксации и диссоциации.Журнал клеточной биологии 146: 1033–1043.
- 29. Ezratty EJ, Partridge MA, Gundersen GG (2005) Индуцированная микротрубочками разборка фокальной адгезии опосредуется динамином и киназой фокальной адгезии. Природа клеточной биологии 7: 581–590.
- 30. Stehbens SJ, Paszek M, Pemble H, Ettinger A, Gierke S, et al. (2014) CLASPs связывают связанный с фокальной адгезией захват микротрубочек с локализованным экзоцитозом и обновлением сайтов адгезии. Природа клеточной биологии 16: 558–573.
- 31.Nalbant P, Chang Y-C, Birkenfeld J, Chang Z-F, Bokoch GM (2009) Фактор обмена гуаниновых нуклеотидов-h2 регулирует миграцию клеток посредством локализованной активации RhoA на переднем крае. Молекулярная биология клетки 20: 4070-4082.
- 32. Хамфрис Дж. Д., Байрон А., Басс М. Д., Крейг С. Е., Пинни Дж. В. и др. (2009) Протеомный анализ интегрин-ассоциированных комплексов идентифицирует RCC2 как двойной регулятор Rac1 и Arf6. Сигнализация науки 2: ra51.
- 33. Schiller HB, Friedel CC, Boulegue C, Faessler R (2011) Количественная протеомика адгезомы интегрина показывает зависимое от миозина II рекрутирование белков домена LIM.Embo Reports 12: 259–266.
- 34. Kuo J-C, Han X, Hsiao C-T, Yates JR III, Waterman CM (2011) Анализ протеома фокальной адгезии, чувствительного к миозину-II, показывает роль бета-Pix в негативной регуляции созревания фокальной адгезии. Природа клеточной биологии 13: 383–393.
- 35. Vizcaino JA, Deutsch EW, Wang R, Csordas A, Reisinger F и др. (2014) ProteomeXchange обеспечивает глобально скоординированное представление и распространение данных протеомики. Природная биотехнология 32: 223–226.
- 36. Хуанг Д.В., Шерман Б.Т., Лемпицки Р.А. (2009) Систематический и интегративный анализ больших списков генов с использованием ресурсов биоинформатики DAVID. Протоколы природы 4: 44–57.
- 37. Хуанг Д.В., Шерман Б.Т., Лемпицки Р.А. (2009) Инструменты обогащения биоинформатики: пути к всестороннему функциональному анализу больших списков генов. Исследование нуклеиновых кислот 37: 1–13.
- 38. Ву Дж., Валлениус Т., Оваска К., Вестермарк Дж., Макела Т.П. и др. (2009) Интегрированная платформа сетевого анализа белок-белковых взаимодействий.Природные методы 6: 75–77.
- 39. Chautard E, Ballut L, Thierry-Mieg N, Ricard-Blum S (2009) MatrixDB, база данных, ориентированная на внеклеточные белок-белковые и белок-углеводные взаимодействия. Биоинформатика 25: 690–691.
- 40. Winograd-Katz SE, Faessler R, Geiger B, Legate KR (2014) Адгезома интегрина: от генов и белков до болезней человека. Nature Reviews Molecular Cell Biology 15: 273–288.
- 41. Чой Х., Фермин Д., Несвижский А.И. (2008) Анализ значимости данных спектрального подсчета в протеомике дробовика без этикеток.Молекулярная и клеточная протеомика 7: 2373–2385.
- 42. Панков Р., Цукерман Э., Кац Б.З., Мацумото К., Лин Д.К. и др. (2000) Динамика интегринов и сборка матрикса: Тензин-зависимая транслокация интегринов альфа (5) бета (1) способствует раннему фибриллогенезу фибронектина. Журнал клеточной биологии 148: 1075–1090.
- 43. Danen EHJ, Sonneveld P, Brakebusch C, Fassler R, Sonnenberg A (2002) Фибронектин-связывающие интегрины альфа 5 бета 1 и альфа v бета 3 по-разному модулируют загрузку RhoA-GTP, организацию адгезий клеточного матрикса и фибриллогенез фибронектина.Журнал клеточной биологии 159: 1071–1086.
- 44. Кац Б.З., Замир Э., Бершадский А., Кам З., Ямада К.М. и др. (2000) Физическое состояние внеклеточного матрикса регулирует структуру и молекулярный состав адгезий клеточного матрикса. Молекулярная биология клетки 11: 1047–1060.
- 45. Danowski BA (1989) Сократимость фибробластов и организация актина стимулируются ингибиторами микротрубочек. Журнал клеточной науки 93: 255–266.
- 46. Schiller HB, Hermann M.-R, Polleux J, Vignaud T, Zanivan S, et al.(2013) интегрины бета (1) и альфа (v) классов кооперируются, чтобы регулировать миозин II во время определения жесткости микроокружений на основе фибронектина. Природа клеточной биологии 15: 625–636.
- 47. Gardel ML, Schneider IC, Aratyn-Schaus Y, Waterman CM (2010) Механическая интеграция актина и динамики адгезии в миграции клеток. Annu Rev Cell Dev Biol 26: 315–333.
- 48. Парсонс JT, Horwitz AR, Schwartz MA (2010) Клеточная адгезия: интеграция динамики цитоскелета и клеточного натяжения.Nature Reviews Molecular Cell Biology 11: 633–643.
- 49. Ватанабе Н., Като Т., Фудзита А., Ишизаки Т., Нарумия С. (1999) Сотрудничество между mDia1 и ROCK в Rho-индуцированной реорганизации актина. Природа клеточной биологии 1: 136–143.
- 50. Riveline D, Zamir E, Balaban NQ, Schwarz US, Ishizaki T. и др. (2001) Фокусные контакты как механодатчики: внешнее приложение локальной механической силы вызывает рост фокальных контактов с помощью mDia1-зависимого и ROCK-независимого механизма.Журнал клеточной биологии 153: 1175–1185.
- 51. Oakes PW, Beckham Y, Stricker J, Gardel ML (2012) Требуется натяжение, но его недостаточно для созревания фокальной адгезии без шаблона напряженных волокон. Журнал клеточной биологии 196: 363–374.
- 52. Goode BL, Eck MJ (2007) Механизм и функция форминов в контроле сборки актина. Ежегодный обзор биохимии. С. 593–627.
- 53. Маэкава М., Ишизаки Т., Боку С., Ватанабэ Н., Фудзита А. и др.(1999) Передача сигналов от rho к актиновому цитоскелету через протеинкиназы ROCK и LIM-kinase. Наука 285: 895–898.
- 54. Лансберген Г., Григорьев И., Мимори-Кийосуэ Ю., Оцука Т., Хига С. и др. (2006) CLASPs прикрепляют плюс концы микротрубочек к коре клетки посредством комплекса с LL5 beta. Клетка развития 11: 21–32.
- 55. ван дер Ваарт Б., ван Риель В.Е., Дудхи Х., Кевенаар Дж.Т., Катруха Е.А. и др. (2013) CFEOM1-ассоциированный кинезин KIF21A является ингибитором роста кортикальных микротрубочек.Клетка развития 27: 145–160.
»Каков макромолекулярный состав клетки?
Каков макромолекулярный состав клетки?
Reader ModeМолекулярная биология направлена на объяснение клеточных процессов с точки зрения индивидуальных молекулярных игроков, что приводит к тому, что главные роли играют определенные специфические белки, РНК и липиды. Напротив, более целостный взгляд на целую клетку или организм исторически был сферой физиологии.Недавно последний интегративный взгляд был принят системной биологией, которая замыкает круг, возвращаясь с с трудом завоеванными механистическими знаниями из молекулярной биологии к целостному взгляду на молекулярные взаимосвязи, которые вызывают поведение всей клетки. Важнейшей отправной точкой для глобального мышления о клетке является понимание относительного количества ее различных составляющих.
Рис. 1. Древовидная диаграмма Вороного, изображающая состав клетки E. coli, растущей со временем удвоения 40 мин.Площадь каждого многоугольника представляет собой относительную долю соответствующего компонента в сухой массе клетки. Цвета связаны с каждым многоугольником, так что компоненты со связанной функциональной ролью имеют одинаковые оттенки. Метод визуализации древовидной диаграммы Вороного был разработан для представления измерений всего генома с помощью микрочипов или количественного анализа протеома.
Такой вид с высоты птичьего полета на состав клетки представлен на рисунке 1 для случая E. coli во время экспоненциального роста с временем удвоения 40 минут.Часть фигуры преобладают обычные подозреваемые, причем белки составляют чуть более половины клеточного содержимого. Более удивительно, что, несмотря на их критическую роль в качестве контролеров экспрессии генов, мРНК составляют лишь небольшую фракцию при анализе с точки зрения абсолютной массы, составляя только около 1% от сухой массы. Рисунок основан на обобщении информации, определенной для клеточного состава E. coli , воссозданного в таблице 1 (BNID 104954). Этот сборник впервые появился в классическом учебнике «Физиология бактериальной клетки» — ярком примере биологического текста, демонстрирующего конструктивную одержимость математикой, которая характеризовала первые дни бактериальной физиологии.Белок оценивается как ≈55% от сухой массы клетки, затем следуют РНК на ≈20%, липиды на ≈10% и ДНК на ≈3% (остальное — полисахариды, метаболиты, ионы и т. Д.). Аналогичные усилия на почкующихся дрожжах показали, что белки составляют в пределах 40-50% сухой массы клетки, РНК ≈10% и липиды ≈10% (BNID 111209, 108196, 108198, 108199, 108200, 102327, 102328). В клетках млекопитающих доля РНК уменьшается примерно до 4%, в то время как доля липидов увеличивается (BNID 111209).
Таблица 1.Объем 3 ячейки). Адаптировано с модификациями из F. C. Neidhardt et al., «Physiology of the bacterial cell», Sinauer, 1990 (BNID 104954). Модификации включали увеличение сухой массы клеток с 284 фг до 300 фг и общей массы клеток с 950 до 1000 фг, а также округление других значений для уменьшения количества значащих цифр, чтобы значения отражали ожидаемые диапазоны неопределенностей. При разной скорости роста объем и масса на клетку могут изменяться в несколько раз. Относительный состав меняется со скоростью роста, но не так значительно.Ожидается, что для данного объема клеток и скорости роста неопределенность большинства свойств будет порядка 10-30% стандартного отклонения. Исходные значения относятся к штамму B / r, но в пределах ожидаемой неопределенности приведенные здесь значения считаются характерными для наиболее распространенных штаммов E. coli. Источники данных можно найти на BNID 111490. Независимый источник более медленных темпов роста можно найти на BNID 111460.
Какова логика этих значений? Например, рРНК, хотя и довольно однообразная с точки зрения своего разнообразия, составляет 2/3 массы рибосомы и, учитывая потребности в постоянном синтезе белка, должна быть обильной.рРНК на самом деле более чем на порядок более распространена, чем все мРНК вместе взятые. В то же время мРНК быстро деградирует с характерным периодом полужизни около 4 минут (BNID 104324) по сравнению с очень стабильной рРНК, которая демонстрирует деградацию ( in vitro ) только через несколько дней (BNID 108023, 108024). Из-за быстрой деградации мРНК общий синтез мРНК, необходимый клетке, не так мал и составляет примерно половину синтеза рРНК (при времени удвоения 40 минут, BNID 100060).В качестве другого примера рационализации клеточного состава предлагается ограничить содержание белка, который является доминирующим компонентом, из-за эффекта скученности. Накопление большего количества белков на единицу объема цитоплазмы может затруднить такие процессы, как диффузия, которая уже примерно в десять раз медленнее внутри клетки, чем в чистой воде. Мы обсуждаем такие эффекты в виньетке на тему «Каковы масштабы времени для диффузии в клетках?». Средняя концентрация белка в цитоплазме уже такова, что средний белок имеет водную гидратную оболочку, состоящую всего из 10 молекул воды, отделяющих его от соседней гидратной оболочки белка.
Количество липидов в «типичной клетке» можно вывести непосредственно из площади поверхности мембраны, хотя для эукариот в оценку необходимо включить множество внутренних мембран, связанных с органеллами. Давайте посмотрим, как такая оценка работает для сфероцилиндрической сигарообразной E. coli . При диаметре ≈1 мкм и характерной скорости роста, когда общая длина составляет ≈2 мкм (цилиндр 1 мкм и два полусферических колпачка диаметром 1 мкм каждая), площадь поверхности имеет элегантный вид A = 2π или ≈6 мкм 2 .Объем также представляет собой изящное геометрическое упражнение, которое приводит к V = 5π / 12, или ≈1,3 мкм 3 (хотя мы часто будем обсуждать его как имеющий объем 1 мкм 3 для простоты, где оценки порядка величины обеспокоенный). Как обсуждалось в виньетке на тему «Какова толщина клеточной мембраны?», Толщина липидного бислоя составляет около 4 нм (в то время как часто упоминаемые более высокие значения могут быть результатом выступающих из мембраны элементов). Таким образом, объем мембраны составляет около 6 мкм 2 * 4 * 10 -3 мкм = 0.024 мкм 3 . При ≈70% воды и ≈30% сухой массы с плотностью ≈1,3 (BNID 104272) общая плотность составляет ≈1,1 (BNID), а объем сухой массы составляет около 1,3 мкм 3 * 1,1 г / см 3 * 0,3 / 1,3 г / см 3 ≈0,33 мкм 3 . Таким образом, липидный бислой занимает примерно 7% сухой массы. Есть два липидных бислоя, внешняя мембрана и клеточная мембрана, поэтому мы должны удвоить это значение до ≈14%. Учитывая, что белки, украшающие мембрану, занимают от четверти до половины ее площади (BNID 105818), мы достаточно близки к эмпирически измеренному значению ≈9%.
Как меняется состав для разных условий роста и у разных организмов? Учитывая, что классический состав для E. coli был получен еще в 60-х и 70-х годах и что сегодня мы регулярно читаем о количественном определении тысяч белков и мРНК, мы могли ожидать, что экспериментальный ответ на этот вопрос будет стандартным упражнением. . Методы количественной оценки белка — это в основном варианты того, что было разработано Лоури в 1951 году. Статья, объявляющая об этих методах, которая после того, как первая заявка была возвращена из-за резких сокращений журнала, по-видимому, стала самой цитируемой статьей в истории науки. более 200 000 цитирований.Несмотря на все свои достоинства и цитаты, методы в этой работе имеют тенденцию быть ограниченными в их точности при применении ко всему набору клеток, что часто превращается в сложные биохимические испытания. Например, другие составляющие клетки, такие как глутатион, главный регулятор окислительно-восстановительного потенциала клетки, могут влиять на показания. В результате полная характеристика переписи клеток для различных условий в основном отсутствует. Эта ситуация ограничивает нашу способность получить истинное физиологическое или системное представление о динамической клетке и требует пересмотра со стороны биологов, объединяющих хорошие экспериментальные руки с количественным уклоном.
66134 Всего просмотров 10 просмотров сегодня
Разборка во вторник: что внутри Bluetooth-динамика?
Вы когда-нибудь задумывались, что внутри динамика Bluetooth?
Во вторник этой недели мы собираемся взглянуть на внутреннюю часть одного из них. Этот водостойкий динамик Bluetooth произведен компанией Taotronics, и это динамик модели TT-SK09.
Bluetooth-динамик TaotronicsОткрытие!
Открыть эту Bluetooth-колонку было непросто.В этом динамике использован интересный метод плавления пластика над перфорированными решетками динамиков.
Конструкция из расплавленного пластикаРасплавленный пластик соскребали плоским ножом. Как только расплавленный пластик был удален, решетки динамиков сразу поднялись. После того, как решетки динамиков были откручены, пластиковый корпус скрепили несколькими стандартными винтами с крестообразным шлицем. Винты были удалены, и корпус был отделен.
Открытый динамик BluetoothДинамики
Главный компонент любой Bluetooth-колонки — динамик! В этом случае есть два динамика мощностью 3 Вт каждый.
Один из 3-ваттных динамиковАккумулятор
Срок службы батареи этого динамика составляет от 8 до 10 часов. Батарея, которую использует этот динамик, имеет каталожный номер 623450. Номер представляет собой батарею 6.2 мм в высоту, 34 мм в ширину и 50 мм в высоту. Такие литиевые батареи часто производятся разными производителями в упаковках одинакового размера, быстрый поиск в Google «батарея 623450» приводит к множеству разных производителей.
Литий-ионный аккумулятор емкостью 1000 мАчЭта батарея также содержит небольшую печатную плату. Эта небольшая плата действует как схема защиты аккумулятора. Эти печатные платы часто обеспечивают защиту от коротких замыканий, перегрузки, перегрузки по току и перенапряжения.
Плата защиты аккумулятораПервичная печатная плата
Основная печатная плата этого динамика Bluetooth выполняет все функции динамика. Эта печатная плата разделена на секции для зарядки аккумулятора, регулирования напряжения, усиления звука и Bluetooth.
Первичная печатная платаНа тыльной стороне основной печатной платы ничего особенного.
Разрезная задняя часть первичной печатной платыИмеется микрофон для голосовых вызовов и 3 кнопки для увеличения и уменьшения громкости, а также управления воспроизведением / паузой.
Кнопки управленияЗарядное устройство
Схема зарядкиПервый раздел, который мы собираемся рассмотреть, — это раздел зарядки.Зарядка осуществляется линейной технологией LTC4054. Микросхема представляет собой корпус SOT-23-5 и требует минимального количества внешних компонентов. Однако одним из необходимых внешних компонентов является резистор для установки тока заряда. В случае этой печатной платы резистор представляет собой резистор сопротивлением 2 кОм с маркировкой R19. Это значение устанавливает скорость заряда LTC4054 на 500 мА. При емкости аккумулятора динамика 1000 мАч для зарядки потребуется около 2 часов. Кроме того, в этой секции источника питания используются два МОП-транзистора AO3401 для переключения между литиевой батареей и источником питания USB, если динамик Bluetooth заряжается.
Блок питания
Схема блока питанияДвигаясь вправо на плате, мы доходим до блока питания. Этот раздел состоит из обычных настроек ускорения. В основе блока питания лежит повышающий преобразователь производства LPS (Low Power Semiconductor), номер детали LP6212. Это довольно типичный повышающий преобразователь, который может выдавать мощность 6 А.
Повышающий преобразовательДля повышающего преобразователя требуется индуктор, диод, входные и выходные конденсаторы; и резисторы для формирования делителя напряжения для механизма обратной связи по напряжению.Схема, используемая на этой плате, очень похожа на типичную прикладную схему, показанную в техническом описании, за исключением больших конденсаторов.
Пример схемы из таблицыАудио
Схема аудиосистемыСледующий раздел электроники содержит всю электронику для управления двумя динамиками. В этом разделе есть два основных компонента, первый — это аудиоусилитель.Усилитель звука — номер детали HT6818, произведенный Jiaxing Heroic Electronic Technology. Этот аудиоусилитель представляет собой стереоусилитель с двумя выходами мощностью до 3,3 Вт.
Усилитель для динамиковВторой основной компонент, расположенный в этой секции, — это операционный усилитель NJRC NJM14558M.
Операционный усилитель 4558Bluetooth
Схема Bluetooth
Последняя основная секция этой печатной платы — это секция Bluetooth.Основным компонентом раздела Bluetooth является система Bluetooth CSR8635. Эта микросхема, изготовленная CSR, предназначена для использования в беспроводных аудиоустройствах, точно так же, как эта Bluetooth-колонка. Этот модуль Bluetooth поддерживает Bluetooth 4.0, беспроводное стереозвук, проводное аудио и т. Д. Микросхема Bluetooth даже содержит регуляторы напряжения и зарядное устройство для литиевой батареи, которое не используется в этом продукте. В паре с модулем Bluetooth используется 128-битная EEPROM FM24C128 производства Fairchild. Он связывается с модулем Bluetooth по двухпроводному последовательному интерфейсу и, скорее всего, содержит информацию о настройках и конфигурации модуля Bluetooth.Последний важный компонент секции Bluetooth — антенна. Этот модуль использует трассирующую антенну, которая использовалась для отправки и получения данных. Есть два индуктора, L6 и L4, которые используются для точной настройки импеданса антенны для достижения оптимальных характеристик.
Трассовая антеннаВторичная печатная плата
Вторичная печатная плата
Внутри динамика находится еще одна небольшая печатная плата.Маленькая плата используется для подключения линейного аудиовхода, питания USB и кнопки питания к основной плате. Интересно отметить, что на вторичной плате есть два светодиода, которые не видны снаружи динамика.
Завершение!
Подобные Bluetooth-колонкине стоят больших денег, обычно от 20 до 30 долларов, но все же в них помещается ряд компонентов, и они обладают достаточно высоким качеством сборки. (Динамик даже прошел сертификацию FCC!)
Все части динамика BluetoothСпасибо, что заглянули во вторник Teardown на этой неделе.Заходите в следующий вторник, чтобы провести еще одну разборку!
Мы всегда ищем новые вещи для разборки. Если у вас есть какие-либо предложения или вы хотите внести что-то в будущий разборный вторник, щелкните здесь, чтобы увидеть мой адрес электронной почты.
Следующий разбор: что внутри радар-детектора?
Глубокое секвенирование в масштабе всего генома выявляет уникальный состав и паттерны экспрессии микроРНК в развивающейся пыльце Oryza sativa | Геномная биология
Воиннет О: Происхождение, биогенез и активность растительных микроРНК. Клетка. 2009, 136: 669–687. 10.1016 / j.cell.2009.01.046.
CAS PubMed Статья PubMed Central Google Scholar
Schwach F, Moxon S, Moulton V, Dalmay T: Расшифровка разнообразия малых РНК в растениях: длинное и короткое. Краткая функция геномной протеомики. 2009, 8: 472–481. 10.1093 / bfgp / elp024.
CAS PubMed Статья PubMed Central Google Scholar
Jones-Rhoades MW, Bartel DP, Bartel B: MicroRNAS и их регулирующая роль в растениях. Annu Rev Plant Biol. 2006, 57: 19–53. 10.1146 / annurev.arplant.57.032905.105218.
CAS PubMed Статья PubMed Central Google Scholar
Борг М., Браунфилд Л., Твелл Д.: Развитие мужских гаметофитов: молекулярная перспектива. J Exp Bot. 2009, 60: 1465–1478. 10.1093 / jxb / ern355.
CAS PubMed Статья PubMed Central Google Scholar
Wei LQ, Xu WY, Deng ZY, Su Z, Xue Y, Wang T: анализ в масштабе генома и сравнение профилей экспрессии генов в развивающейся и проросшей пыльце Oryza sativa . BMC Genomics. 2010, 11: 338-10.1186 / 1471-2164-11-338.
PubMed PubMed Central Статья CAS Google Scholar
Пина С., Пинто Ф., Фейо Дж. А., Беккер Д.: Анализ семейства генов транскриптома пыльцы Arabidopsis показывает биологические последствия для роста клеток, контроля деления и регуляции экспрессии генов.Plant Physiol. 2005, 138: 744–756. 10.1104 / pp.104.057935.
CAS PubMed PubMed Central Статья Google Scholar
Slotkin RK, Vaughn M, Borges F, Tanurdzic M, Becker JD, Feijo JA, Martienssen RA: Эпигенетическое репрограммирование и подавление малых РНК мобильных элементов в пыльце. Клетка. 2009, 136: 461–472. 10.1016 / j.cell.2008.12.038.
CAS PubMed PubMed Central Статья Google Scholar
Borges F, Gomes G, Gardner R, Moreno N, McCormick S, Feijo JA, Becker JD: Сравнительная транскриптомика сперматозоидов Arabidopsis . Plant Physiol. 2008, 148: 1168–1181. 10.1104 / pp.108.125229.
CAS PubMed PubMed Central Статья Google Scholar
Грант-Даунтон Р., Хафид С., Твелл Д., Дикинсон Х.Г. Пути малых РНК присутствуют и функционируют в мужском гаметофите покрытосеменных. Завод Мол.2009, 2: 500–512. 10.1093 / mp / ssp003.
CAS PubMed Статья PubMed Central Google Scholar
Грант-Даунтон Р., Трионнер Г.Л., Шмид Р., Родригес-Энрикес Дж., Саид Хафидх3 С.М., Твелл Д., Дикинсон Х. Разнообразие микроРНК и тасиРНК в зрелой пыльце Arabidopsis thaliana . BMC Genomics. 2009, 10: 643-10.1186 / 1471-2164-10-643.
PubMed PubMed Central Статья CAS Google Scholar
Chambers C, Shuai B: Профилирование экспрессии микроРНК в пыльце Arabidopsis с использованием массива микроРНК и ПЦР в реальном времени. BMC Plant Biol. 2009, 9: 87-
PubMed PubMed Central Статья CAS Google Scholar
Henderson IR, Zhang XY, Lu C, Johnson L, Meyers BC, Green PJ, Jacobsen SE: Dissecting Arabidopsis thaliana Функция DICER в процессинге малых РНК, сайленсинге генов и формировании паттерна метилирования ДНК.Нат Жене. 2006, 38: 721–725. 10.1038 / ng1804.
CAS PubMed Статья PubMed Central Google Scholar
Като М., де Ленкастр А., Пинкус З., Slack FJ: Динамическая экспрессия малых некодирующих РНК, включая новые микроРНК и piРНК / 21U-РНК, во время развития Caenorhabditis elegans . Genome Biol. 2009, 10: R54-10.1186 / gb-2009-10-5-r54.
PubMed PubMed Central Статья CAS Google Scholar
Reddy AM, Zheng Y, Jagadeeswaran G, Macmil SL, Graham WB, Roe BA, Desilva U, Zhang WX, Sunkar R: Клонирование, характеристика и анализ экспрессии микроРНК свиней. BMC Genomics. 2008, 10: 65-
Статья CAS Google Scholar
Li R, Li Y, Kristiansen K, Wang J: SOAP: Short Oligonucleotide Alignment Program. Биоинформатика (Оксфорд). 2008, 24: 713–714. 10.1093 / биоинформатика / btn025.
CAS Статья Google Scholar
Гриффитс-Джонс С., Грокок Р. Дж., Ван Донген С., Бейтман А., Энрайт А. Дж.: MiRBase: последовательности микроРНК, мишени и номенклатура генов. Nucleic Acids Res. 2006, 34: D140–144. 10.1093 / нар / gkj112.
CAS PubMed Статья PubMed Central Google Scholar
Гриффитс-Джонс С: Реестр микроРНК. Nucleic Acids Res. 2004, 32: D109-111. 10.1093 / нар / гх023.
CAS PubMed PubMed Central Статья Google Scholar
Гриффитс-Джонс С., Сайни Г.К., ван Донген С., Энрайт А.Дж.: miRBase: инструменты для геномики микроРНК. Nucleic Acids Res. 2008, 36: D154–158. 10.1093 / нар / gkn221.
CAS PubMed Статья PubMed Central Google Scholar
Girard A, Hannon GJ: Консервированные темы в контроле транспозонов, опосредованных малой РНК. Trends Cell Biol. 2008, 18: 136–148. 10.1016 / j.tcb.2008.01.004.
CAS PubMed PubMed Central Статья Google Scholar
Аравин А.А., Лагос-Кинтана М., Ялчин А., Заволан М., Маркс Д., Снайдер Б., Гаастерланд Т., Мейер Дж., Тушл Т. Профиль малых РНК во время развития Drosophila melanogaster . Dev Cell. 2003, 5: 337–350. 10.1016 / S1534-5807 (03) 00228-4.
CAS PubMed Статья PubMed Central Google Scholar
Kurihara Y, Watanabe Y: Arabidopsis биогенез микро-РНК посредством функций белка Dicer-like 1.Proc Natl Acad Sci USA. 2004, 101: 12753–12758. 10.1073 / pnas.0403115101.
CAS PubMed Статья PubMed Central Google Scholar
Рейнхарт Б.Дж., Вайнштейн Э.Г., Родес М.В., Бартель Б., Бартель Д.П.: МикроРНК в растениях. Genes Dev. 2002, 16: 1616–1626. 10.1101 / гад.1004402.
CAS PubMed PubMed Central Статья Google Scholar
Мейерс BC, Axtell MJ, Бартель B, Бартел Д. П., Баулкомб Д., Боуман JL, Cao X, Кэррингтон JC, Чен XM, Грин PJ, Гриффитс-Джонс S, Jacobsen SE, Мэллори AC, Martienssen RA, Poethig RS, Qi Y , Vaucheret H, Voinnet O, Watanabe Y, Weigel D, Zhu JK: Критерии аннотации растительных микроРНК. Растительная клетка. 2008, 20: 3186–3190. 10.1105 / tpc.108.064311.
CAS PubMed PubMed Central Статья Google Scholar
Fahlgren N, Howell MD, Kasschau KD, Chapman EJ, Sullivan CM, Cumbie JS, Givan SA, Law TF, Grant SR, Dangl JL, Carrington JC: высокопроизводительное секвенирование микроРНК арабидопсиса : доказательства для частого рождения и смерти генов MIRNA.PLoS One. 2007, 2: e219-10.1371 / journal.pone.0000219.
PubMed PubMed Central Статья CAS Google Scholar
Sunkar R, Zhou X, Zheng Y, Zhang W, Zhu JK: Идентификация новых и потенциальных miRNAs в рисе с помощью высокопроизводительного секвенирования. BMC Plant Biol. 2008, 8: 25-10.1186 / 1471-2229-8-25.
PubMed PubMed Central Статья CAS Google Scholar
Rajagopalan R, Vaucheret H, Trejo J, Bartel DP: разнообразный и эволюционно подвижный набор микроРНК в Arabidopsis thaliana . Genes Dev. 2006, 20: 3407–3425. 10.1101 / гад.1476406.
CAS PubMed PubMed Central Статья Google Scholar
Zhang G, Guo G, Hu X, Zhang Y, Li Q, Li R, Zhuang R, Lu Z, He Z, Fang X, Chen L, Tian W, Tao Y, Kristiansen K, Zhang X , Li S, Yang H, Wang J, Wang J: Глубокое секвенирование РНК при разрешении одной пары оснований показывает высокую сложность транскриптома риса.Genome Res. 2010, 20: 646–654. 10.1101 / gr.100677.109.
CAS PubMed PubMed Central Статья Google Scholar
Ouyang S, Buell CR: TIGR Plant Repeat Databases: коллективный ресурс для идентификации повторяющихся последовательностей в растениях. Nucleic Acids Res. 2004, 32: D360 – D363. 10.1093 / нар / гх099.
CAS PubMed PubMed Central Статья Google Scholar
Zhang YJ: miRU: автоматизированный сервер прогнозирования мишеней miRNA. Nucleic Acids Res. 2005, 33: W701 – W704. 10.1093 / нар / gki383.
CAS PubMed PubMed Central Статья Google Scholar
Мэллори AC, Vaucheret H: Функции микроРНК и родственных малых РНК в растениях. Нат Жене. 2006, 38: S31 – S36. 10.1038 / ng1791.
CAS PubMed Статья PubMed Central Google Scholar
Lu C, Jeong DH, Kulkarni K, Pillay M, Nobuta K, German R, Thatcher SR, Maher C, Zhang L, Ware D, Liu B, Cao X, Meyers BC, Green PJ: анализ генома для обнаружения микроРНК риса выявляют природные антисмысловые микроРНК (nat-miRNA). Proc Natl Acad Sci USA. 2008, 105: 4951–4956. 10.1073 / pnas.0708743105.
CAS PubMed Статья PubMed Central Google Scholar
Luo YC, Zhou H, Li Y, Chen JY, Yang JH, Chen YQ, Qu LH: Эмбриогенные каллусы риса экспрессируют уникальный набор микроРНК, что указывает на регуляторную роль микроРНК в постэмбриогенном развитии растений.FEBS Lett. 2006, 580: 5111–5116. 10.1016 / j.febslet.2006.08.046.
CAS PubMed Статья PubMed Central Google Scholar
German MA, Pillay M, Jeong DH, Hetawal A, Luo SJ, Janardhanan P, Kannan V, Rymarquis LA, Nobuta K, German R, German R, De Paoli E, Lu C, Schroth G, Meyers BC, Green PJ: Глобальная идентификация пар микроРНК-мишень РНК путем параллельного анализа концов РНК. Nat Biotechnol. 2008, 26: 941–946.10.1038 / nbt1417.
CAS PubMed Статья PubMed Central Google Scholar
Аддо-Куэй С., Снайдер Дж. А., Парк Й.Б., Ли Й.Ф., Сункар Р., Акстелл М.Дж .: Нарезанные мишени микроРНК и точная обработка шпилек MIR319 с первой петлей, выявленная при анализе деградома Physcomitrella patens . РНК. 2009, 15: 2112–2121. 10.1261 / rna.1774909.
CAS PubMed PubMed Central Статья Google Scholar
Addo-Quaye C, Eshoo TW, Bartel DP, Axtell MJ: Эндогенные миРНК и миРНК-мишени, идентифицированные путем секвенирования деградома Arabidopsis . Curr Biol. 2008, 18: 758–762. 10.1016 / j.cub.2008.04.042.
CAS PubMed PubMed Central Статья Google Scholar
Wu L, Zhang QQ, Zhou HY, Ni FR, Wu XY, Qi YJ: эффекторные комплексы и мишени микроРНК риса. Растительная клетка. 2009, 21: 3421–3435. 10.1105 / тпк.109.070938.
CAS PubMed PubMed Central Статья Google Scholar
Li YF, Zheng Y, Addo-Quaye C, Zhang L, Saini A, Jagadeeswaran G, Axtell MJ, Zhang WX, Sunkar R: Идентификация всех микроРНК в рисе по транскриптому. Плант Дж. 2010, 62: 742–759. 10.1111 / j.1365-313X.2010.04187.x.
CAS PubMed Статья PubMed Central Google Scholar
Yang JH, Li JH, Shao P, Zhou H, Chen YQ, Qu LH: starBase: база данных для изучения карт взаимодействия микроРНК-мРНК из данных Argonaute CLIP-Seq и Degradome-Seq. Nucleic Acids Res. 2010, 39: D202–209.
PubMed PubMed Central Статья CAS Google Scholar
Carthew RW, Sontheimer EJ: Происхождение и механизмы миРНК и миРНК. Клетка. 2009, 136: 642–655. 10.1016 / j.cell.2009.01.035.
CAS PubMed PubMed Central Статья Google Scholar
Гамильтон А, Воиннет О., Чаппелл Л., Баулкомб Д. Два класса коротких интерферирующих РНК в молчании РНК. EMBO J. 2002, 21: 4671–4679. 10.1093 / emboj / cdf464.
CAS PubMed PubMed Central Статья Google Scholar
Chen X: Малые РНК и их роль в развитии растений. Annu Rev Cell Dev Biol. 2009, 25: 21–44. 10.1146 / annurev.cellbio.042308.113417.
PubMed PubMed Central Статья CAS Google Scholar
Zilberman D, Cao X, Jacobsen SE: ARGONAUTE4 контроль локус-специфичного накопления миРНК и метилирования ДНК и гистонов. Наука. 2003, 299: 716–719. 10.1126 / science.1079695.
CAS PubMed Статья PubMed Central Google Scholar
Чан С.В., Зильберман Д., Се З., Йохансен Л.К., Кэррингтон Дж.С., Якобсен С.Е.: гены подавления РНК контролируют метилирование ДНК de novo. Наука. 2004, 303: 1336-10.1126 / science.1095989.
CAS PubMed Статья PubMed Central Google Scholar
Wu L, Zhou HY, Zhang QQ, Zhang JG, Ni FR, Liu C, Qi YJ: метилирование ДНК, опосредованное путем микроРНК. Mol Cell. 2010, 38: 465–475. 10.1016 / j.molcel.2010.03.008.
CAS PubMed Статья PubMed Central Google Scholar
Ван Х, Эллинг А.А., Ли Х, Ли Н, Пэн З, Хе Г, Сан Х, Ци И, Лю XS, Дэн ХУ: общегеномные и органоспецифичные ландшафты эпигенетических модификаций и их взаимосвязи. к транскриптомам мРНК и малых РНК кукурузы.Растительная клетка. 2009, 21: 1053–1069. 10.1105 / tpc.109.065714.
CAS PubMed PubMed Central Статья Google Scholar
Valoczi A, Varallyay E, Kauppinen S, Burgyan J, Havelda Z: Пространственно-временное накопление микроРНК очень скоординировано в развивающихся тканях растений. Плант J. 2006, 47: 140–151. 10.1111 / j.1365-313X.2006.02766.x.
CAS PubMed Статья PubMed Central Google Scholar
Sieber P, Wellmer F, Gheyselinck J, Riechmann JL, Meyerowitz EM: Избыточность и специализация среди растительных микроРНК: роль семейства MIR164 в устойчивости развития. Разработка. 2007, 134: 1051–1060. 10.1242 / dev.02817.
CAS PubMed Статья PubMed Central Google Scholar
Чжан Б.Х., Пан XP, Кэннон С.Х., Кобб Г.П., Андерсон Т.А.: Сохранение и дивергенция генов микроРНК растений. Плант Дж. 2006, 46: 243–259.10.1111 / j.1365-313X.2006.02697.x.
CAS PubMed Статья PubMed Central Google Scholar
Zhang BH, Pan XP, Anderson TA: Идентификация 188 консервативных микроРНК кукурузы и их мишеней. FEBS Lett. 2006, 580: 3753–3762. 10.1016 / j.febslet.2006.05.063.
CAS PubMed Статья PubMed Central Google Scholar
Морин Р.Д., Аксай Г., Долгошина Е., Эбхардт Х.А., Магрини В., Мардис Э.Р., Сахиналп С.К., Унрау П.Дж.: Сравнительный анализ транскриптомов малых РНК Pinus contorta и Oryza sativa.Genome Res. 2008, 18: 571–584. 10.1101 / гр.6897308.
CAS PubMed PubMed Central Статья Google Scholar
Грэм Л.К., Уилкокс Л.В.: Происхождение смены поколений у наземных растений: акцент на матротрофии и транспорте гексозы. Philos Trans R Soc Lond B Biol Sci. 2000, 355: 757–766. 10.1098 / rstb.2000.0614. обсуждение 766–757
CAS PubMed PubMed Central Статья Google Scholar
Tang F: Малые РНК в зародышевой линии млекопитающих: Крошечные для бессмертных. Дифференциация. 2010, 79: 141–146. 10.1016 / j.diff.2009.11.002.
CAS PubMed Статья PubMed Central Google Scholar
Honys D, Twell D: Транскриптомный анализ развития гаплоидных мужских гаметофитов у арабидопсиса . Genome Biol. 2004, 5: R85-10.1186 / GB-2004-5-11-r85.
PubMed PubMed Central Статья Google Scholar
Achard P, Herr A, Baulcombe DC, Harberd NP: Модуляция развития цветков с помощью регулируемой гиббереллином микроРНК. Разработка. 2004, 131: 3357–3365. 10.1242 / dev.01206.
CAS PubMed Статья PubMed Central Google Scholar
Миллар А.А., Габлер F: GAMYB-подобные гены Arabidopsis , MYB33 и MYB65, представляют собой регулируемые микроРНК гены, которые избыточно способствуют развитию пыльников. Растительная клетка. 2005, 17: 705–721.10.1105 / tpc.104.027920.
CAS PubMed PubMed Central Статья Google Scholar
Wu MF, Tian Q, Reed JW: Arabidopsis microRNA167 контролирует паттерны экспрессии ARF6 и ARF8 и регулирует воспроизводство как самок, так и самцов. Разработка. 2006, 133: 4211–4218. 10.1242 / dev.02602.
CAS PubMed Статья PubMed Central Google Scholar
Guo HS, Xie Q, Fei JF, Chua NH: МикроРНК направляет расщепление мРНК фактора транскрипции NAC1 для подавления сигналов ауксина для развития боковых корней арабидопсиса . Растительная клетка. 2005, 17: 1376–1386. 10.1105 / tpc.105.030841.
CAS PubMed PubMed Central Статья Google Scholar
Jones-Rhoades MW, Bartel DP: Вычислительная идентификация растительных микроРНК и их мишеней, включая индуцированную стрессом миРНК.Mol Cell. 2004, 14: 787–799. 10.1016 / j.molcel.2004.05.027.
CAS PubMed Статья PubMed Central Google Scholar
Cecchetti V, Altamura MM, Falasca G, Costantino P, Cardarelli M: Arabidopsis ауксин регулирует расхождение пыльников, созревание пыльцы и удлинение волокон. Растительная клетка. 2008, 20: 1760–1774. 10.1105 / tpc.107.057570.
CAS PubMed PubMed Central Статья Google Scholar
de Martinis D, Cotti G, Hekker ST, Harren FJM, Mariani C: Этиленовая реакция на рост пыльцевых трубок в цветках Nicotiana tabacum . Planta. 2002, 214: 806–812. 10.1007 / s00425-001-0684-2.
CAS PubMed Статья PubMed Central Google Scholar
Huang S, Cerny RE, Qi YL, Bhat D, Aydt CM, Hanson DD, Malloy KP, Ness LA: Трансгенные исследования участия цитокинина и гиббереллина в развитии мужчин.Plant Physiol. 2003, 131: 1270–1282. 10.1104 / pp.102.018598.
CAS PubMed PubMed Central Статья Google Scholar
Park JH, Halitschke R, Kim HB, Baldwin IT, Feldmann KA, Feyereisen R: нокаут-мутация в синтазе алленоксида приводит к мужской стерильности и дефектной трансдукции раневого сигнала у Arabidopsis из-за блока в биосинтезе жасмоновой кислоты. Плант J. 2002, 31: 1–12. 10.1046 / j.1365-313X.2002.01328.x.
PubMed Статья PubMed Central Google Scholar
Cheng H, Song SS, Xiao LT, Soo HM, Cheng ZW, Xie DX, Peng JR: Гиббереллин действует через жасмонат, чтобы контролировать экспрессию MYB21, MYB24 и MYB57, способствуя росту тычинок у арабидопсиса . PLoS Genet. 2009, 5: e1000440-10.1371 / journal.pgen.1000440.
PubMed PubMed Central Статья CAS Google Scholar
Baker SM, Bronner CE, Zhang L, Plug AW, Robatzek M, Warren G, Elliott EA, Yu JA, Ashley T., Arnheim N, Flavell RA, Liskay RM: самцы мышей с дефектом гена восстановления несоответствия ДНК Pms2 демонстрируют аномальные хромосомный синапс в мейозе. Клетка. 1995, 82: 309–319. 10.1016 / 0092-8674 (95) -6.
CAS PubMed Статья PubMed Central Google Scholar
Komori T, Ohta S, Murai N, Takakura Y, Kuraya Y, Suzuki S, Hiei Y, Imaseki H, Nitta N: клонирование на основе карты гена восстановления фертильности, Rf-1, в рисе ( Oryza sativa L.). Плант Дж. 2004, 37: 315–325.
CAS PubMed Статья PubMed Central Google Scholar
Saze H, Kakutani T: наследственная эпигенетическая мутация фланкированного транспозоном гена Arabidopsis из-за отсутствия фактора ремоделирования хроматина DDM1. EMBO J. 2007, 26: 3641–3652. 10.1038 / sj.emboj.7601788.
CAS PubMed PubMed Central Статья Google Scholar
O’Toole N, Hattori M, Andres C, Iida K, Lurin C, Schmitz-Linneweber C, Sugita M, Small I: О расширении семейства генов пентатрикопептидных повторов в растениях. Mol Biol Evol. 2008, 25: 1120–1128. 10.1093 / molbev / msn057.
CAS PubMed Статья PubMed Central Google Scholar
Le Trionnaire G, Twell D: Малые РНК в гаметофитах покрытосеменных: от эпигенетики до развития гамет. Genes Dev. 2010, 24: 1081–1085.10.1101 / gad.1936110.
CAS PubMed PubMed Central Статья Google Scholar
Cartagena JA, Matsunaga S, Seki M, Kurihara D, Yokoyama M, Shinozaki K, Fujimoto S, Azumi Y, Uchiyama S, Fukui K: Arabidopsis SDG4 способствует регулированию роста пыльцевых трубок путем метилирование лизинов 4 и 36 гистона h4 в зрелой пыльце. Dev Biol. 2008, 315: 355–368. 10.1016 / j.ydbio.2007.12.016.
CAS PubMed Статья PubMed Central Google Scholar
Ито М., Коике А., Коидзуми Н., Сано Н: Метилированные ДНК-связывающие белки из арабидопсиса . Plant Physiol. 2003, 133: 1747–1754. 10.1104 / стр.103.026708.
CAS PubMed PubMed Central Статья Google Scholar
Земах А., МакДэниел И.Е., Сильва П., Зильберман Д.: Полногеномный эволюционный анализ метилирования эукариотической ДНК. Наука. 2010, 328: 916–919. 10.1126 / science.1186366.
CAS PubMed Статья PubMed Central Google Scholar
Ли Т.Ф., Чжай Дж. Х., Мейерс BC: Сохранение и расхождение в метилировании эукариотической ДНК. Proc Natl Acad Sci USA. 2010, 107: 9027–9028. 10.1073 / pnas.1005440107.
CAS PubMed Статья PubMed Central Google Scholar
Zemach A, Grafi G: Характеристика белков Arabidopsis thaliana метил-CpG-связывающего домена (MBD). Плант Дж. 2003, 34: 565–572. 10.1046 / j.1365-313X.2003.01756.x.
CAS PubMed Статья PubMed Central Google Scholar
Rousseaux S, Caron C, Govin K, Lestrat C, Faure AK, Khochbin S: Установление специфической для мужчин эпигенетической информации. Ген. 2005, 345: 139–153. 10.1016 / j.gene.2004.12.004.
CAS PubMed Статья PubMed Central Google Scholar
Trasler JM: Эпигенетика в сперматогенезе. Mol Cell Endocrinol. 2009, 306: 33–36. 10.1016 / j.mce.2008.12.018.
CAS PubMed Статья PubMed Central Google Scholar
Liu SP, Li D, Li QB, Zhao P, Xiang ZH, Xia QY: микроРНК Bombyx mori , идентифицированные с помощью секвенирования Solexa. BMC Genomics. 2010, 11: 148-10.1186 / 1471-2164-11-148.
PubMed PubMed Central Статья CAS Google Scholar
ArrayExpress. [https://doi.org/www.ebi.ac.uk/arrayexpress/]
Qi X, Bao FS, Xie Z: Глубокое секвенирование малых РНК выявляет роль РНК-зависимой РНК талианы Arabidopsis полимеразы в биогенезе вирусных миРНК.PLoS One. 2009, 4: e4971-10.1371 / journal.pone.0004971.
PubMed PubMed Central Статья CAS Google Scholar
Gardner PP, Daub J, Tate JG, Nawrocki EP, Kolbe DL, Lindgreen S, Wilkinson AC, Finn RD, Griffiths-Jones S, Eddy SR, Bateman A: Rfam: обновления базы данных семейств RNA. Nucleic Acids Res. 2009, 37: D136 – D140. 10.1093 / нар / gkn766.
CAS PubMed Статья PubMed Central Google Scholar
Гриффитс-Джонс С., Бейтман А., Маршалл М., Ханна А., Эдди С.Р .: Rfam: база данных семейства РНК. Nucleic Acids Res. 2003, 31: 439–441. 10.1093 / нар / gkg006.
CAS PubMed PubMed Central Статья Google Scholar
Griffiths-Jones S, Moxon S, Marshall M, Khanna A, Sr E, Bateman A: Rfam: аннотирование некодирующих РНК в полных геномах. Nucleic Acids Res. 2005, 33: D121 – D124.
CAS PubMed Статья PubMed Central Google Scholar
GenBank. [https://doi.org/www.ncbi.nlm.nih.gov/genbank/]
Chen C, Ridzon DA, Broomer AJ, Zhou Z, Lee DH, Nguyen JT, Barbisin M, Xu Н.Л., Махувакар В.Р., Андерсен М.Р., Лао К.К., Ливак К.Дж., Геглер К.Дж.: Количественная оценка микроРНК в реальном времени с помощью ОТ-ПЦР «стебель-петля». Nucleic Acids Res. 2005, 33: e179-10.1093 / nar / gni178.
PubMed PubMed Central Статья CAS Google Scholar
Ван К., Хуанг З., Сюэ HL, Джин С.К., Джу XL, Хан JDJ, Чен Ю.Г .: МикроРНК miR-24 ингибирует эритропоэз, воздействуя на рецептор активина I типа ALK4.Кровь. 2008, 111: 588–595. 10.1182 / кровь-2007-05-092718.
CAS PubMed Статья PubMed Central Google Scholar
Bu Y, Lu CH, Bian CJ, Wang JH, Li J, Zhang B, Li ZY, Brewer G, Zhao RC: нокдаун Dicer в клетках карциномы молочной железы человека MCF-7 приводит к остановке G1 и увеличению чувствительность к цисплатину. Oncol Rep. 2009, 21: 13–17.
CAS PubMed PubMed Central Google Scholar
Varkonyi-Gasic E, Wu RM, Wood M, Walton EF, Hellens RP: Протокол: высокочувствительный метод RT-PCR для обнаружения и количественной оценки микроРНК. Растительные методы. 2007, 3: 12-10.1186 / 1746-4811-3-12.
PubMed PubMed Central Статья CAS Google Scholar
Lacombe S, Nagasaki H, Santi C, Duval D, Piegu B, Bangratz M, Breitler JC, Guiderdoni E, Brugidou C, Hirsch J, Cao X, Brice C, Panaud O, Karlowski WM, Sato Y , Echeverria M: Идентификация транскриптов-предшественников для 6 новых miRNA расширяет разнообразие геномной организации и экспрессии генов miRNA в рисе.BMC Plant Biol. 2008, 8: 123-10.1186 / 1471-2229-8-123.
PubMed PubMed Central Статья CAS Google Scholar
Урожай самолетов: как получить прибыль от вывода самолетов из эксплуатации
По мере того, как эксплуатанты ищут более экономичные и экологически чистые самолеты, использующие одни из самых дешевых вариантов финансирования крупногабаритного оборудования за последние десятилетия, вывод из эксплуатации старых моделей ускоряется. Эксперты в области авиации прогнозируют, что в ближайшие 10 лет во всем мире будет заменено от 6 000 до 7 000 коммерческих самолетов.Исторически низкие процентные ставки, более высокие затраты на топливо и новые технологии — все это способствует увеличению выбытия самолетов.
В настоящее время мировой парк коммерческого воздушного транспорта составляет около 23 000 самолетов, как реактивных, так и турбовинтовых. При ежегодном росте воздушного движения примерно на 5 процентов и более 3 миллиардах пассажиров в год поставки самолетов превысят 16 000 в следующие 10 лет. После вывода из эксплуатации самолетов к 2024 году мировой парк составит более 32000 самолетов.
По прогнозам, в ближайшие 10 лет состав выбывающих самолетов будет включать примерно 43 процента узкофюзеляжных, 21 процент широкофюзеляжных, 28 процентов региональных реактивных самолетов и 8 процентов турбовинтовых самолетов. Учитывая исторически высокий уровень использования арендованных самолетов операторами, простота замены старых моделей, безусловно, сохранится. Итак, что произойдет со всеми этими стареющими самолетами, которые будут выведены из эксплуатации и поставлены на стоянку?
Ряд самолетов, примерно от 2 000 до 3 000, были просто брошены в основном в развивающихся странах.Многие из них списаны из-за большого количества разобранных деталей и общего износа самолетов. Остальные припаркованы в различных хранилищах, преимущественно на юго-западе США, где климат является оптимальным, чтобы свести к минимуму коррозию.
Для большинства этих самолетов двигатели несут большую часть остаточной стоимости и, если они останутся на крыле, в какой-то момент будут возвращены в эксплуатацию. Большинство устаревших самолетов, таких как DC-8, DC-9, DC-10; B707, B727, старые B737 и B747 больше не будут летать.Эти самолеты первого поколения предлагают очень мало с точки зрения частичного использования. Однако неклассические модели Boeing и Airbus представляют собой привлекательные перспективы для демонтажа и могут принести пользу операторам благодаря поддержке компонентов.
Решения по окончании срока службы
Newcastle Aviation специализируется на решениях по окончании срока службы для платформ самолетов, оценивая определенные типы планеров, двигателей и компонентов, которые, по нашему мнению, принесут максимальную пользу рынку, поскольку динамика спроса / предложения продолжает развиваться, а мировой парк меняется.При рассмотрении и последующей обработке конкретного летательного аппарата для разборки в игру вступает ряд факторов. Помимо геополитических, нормативных и правительственных последствий, связанных с местонахождением актива, необходимо тщательно спланировать и выполнить выполнение предпродажной проверки документации и физического осмотра. Но прежде чем это произойдет, необходимо сделать один важный шаг. Мы проводим тщательный анализ профиля спроса на актив, чтобы получить четкое представление о стоимости демонтажа и оптимальной стратегии выхода для этого актива.
Типичный коммерческий узкофюзеляжный самолет, например, при разборке от носа до хвоста, будет производить около 2000 деталей (только планер) в зависимости от того, насколько обширна эта деталь. Используя иллюстрированные каталоги запчастей (IPC), разработанные Newcastle Aviation для оценки стоимости авиационных запчастей и моделирование прогноза спроса, мы можем определить, какие компоненты принесут наибольшую пользу рынку при наилучшей окупаемости инвестиций. Правило 80/20 обычно применяется к любому частичному проекту: 20 процентов деталей обычно несут 80 процентов стоимости, и именно этим компонентам уделяется наибольшее внимание.
Классификация деталей
Помимо сложной оценки и моделирования спроса, еще одним ключевым моментом при рассмотрении вопроса о монетизации самолета является критичность диспетчеризации каждого компонента. Детали, которые критичны для полета и не подлежат использованию, помечаются как элементы Категории 1. Гидравлические компоненты, колеса и тормоза, приборы кабины экипажа и навигационное оборудование — вот некоторые примеры таких деталей. Даже если анализ спроса несколько слаб, а насыщенность рынка высока, эти детали по-прежнему являются хорошими кандидатами на склад для поддержки клиентов.
Детали категории 2 имеют решающее значение, но они перечислены в Главном перечне минимального оборудования (MMEL), поэтому они имеют определенный уровень облегчения при отправке. Детали категории 3 охватывают широкий спектр разделов ATA, включая аксессуары и средства управления полетом.
Наконец, компонентами категории 4, как правило, являются внутренние детали и структурные элементы. Категоризация компонентов помогает расставить приоритеты в рабочем процессе во время разгрузки и добавляет дополнительный уровень интеллекта при рассмотрении того, что нужно удалить с самолета.
Логистика и контроль качества
Логистика и контроль качества являются важными элементами любого проекта по демонтажу. Где физически расположен актив и его летная годность определяет первые шаги; должен ли самолет быть перемещен на частичный объект или механиков можно отправить туда, где он в настоящее время припаркован. Безопасность актива превыше всего.
После перехода права собственности важно, чтобы самолет находился в надежном и безопасном месте и был в целости и сохранности, как описано в договоре купли-продажи.Если самолет необходимо разобрать в удаленном месте, необходимо также решить экологические проблемы. Наряду с этим должна быть организована окончательная утилизация корпуса.
Любые компоненты, снятые с самолета, должны быть выполнены в соответствии с руководством по техническому обслуживанию и идентифицированы с помощью IPC самолета. Для каждой детали требуется бирка снятия деталей, на которой указан точный номер детали, описание, состояние, а также серийный номер самолета, дата, имя механика и номер лицензии. Основная таблица удаления обновляется ежедневно.
Затраты на инвентаризацию и ремонт
Еще одним важным аспектом любого проекта разборки является то, что после того, как детали сняты и теперь находятся на складе в снятом состоянии, какой уровень ремонтных долларов следует вложить в инвентарь на передней части? В какие детали нужно вкладывать деньги в ремонт? Какие из них не должны? Какова стратегия выхода для обоих?
Операторы часто не могут освоить время выполнения заказа на деталь из-за критичности требований. Таким образом, требуется определенная сумма инвестиций для того, чтобы запасные части были на складе в кратчайшие сроки.Использование вышеупомянутого анализа спроса снова дает надежный инструмент, который определит конкретные компоненты, которые необходимо проверить, протестировать, отремонтировать или отремонтировать. Кроме того, детали категории 1, которые могут помешать отправке рейса, тщательно проверяются на предмет возможности ремонта. Даже если общий спрос невелик, эти закрытые единицы являются главными кандидатами для инвестиций. Конечно, даже при таком типе анализа тенденции рыночного спроса и возможное насыщение предложения любого компонента по-прежнему необходимо тщательно оценивать и контролировать на постоянной основе. основание.
Пенсионные стратегии и управление активами
Пер Стив Хендриксон, генеральный директор Newcastle Aviation: «Наша способность предлагать экономичные решения по поддержке материалов и управлению активами зависит от нашей способности разрабатывать и использовать самые передовые инструменты рыночной информации и оценки активов, доступные сегодня. Это критически важно для помощь нашим конечным клиентам в сокращении и управлении их эксплуатационными расходами и расходами на владение на протяжении всего жизненного цикла их автопарков. Это включает в себя первоначальное резервирование запасов, экономию на масштабе, управление ремонтом и объединение запасных частей, лизинг активов, обратную аренду и консигнацию товарных запасов, а также работу в тесном сотрудничестве с ними, чтобы разработать и реализовать стратегии вывода из эксплуатации парка самолетов, которые позволят максимально увеличить отдачу от устаревших самолетов, инвентаря и соответствующего вспомогательного оборудования.«
Поскольку мировой коммерческий флот продолжает трансформироваться как по размеру, так и по составу, количество списываемых самолетов увеличивается. Средний возраст самолетов упал ниже 24 лет, и, учитывая, что в 2008 году он составлял почти 30 лет, это значительное снижение. Кроме того, с появлением новых самолетов, таких как A320NEO и B737MAX, дальнейшее давление будет оказываться на размещение мирового парка существующих A320 и B737NG.
Примерно одна треть мирового флота находится в Северной Америке, в Западной Европе — 20 процентов, в Восточной Европе — 5 процентов, а в Китае, Азиатско-Тихоокеанском регионе и Индии вместе взятых — около 25 процентов.