Порядок разбора слова по составу Выделить окончание. Выделить основу. Найти корень. Выделить приставки и суффиксы.
Нажмите для полного просмотра!
Содержание ▲
- Порядок разбора слова по составу Выделить…
- Разбор по составу некоторых частей речи: …
- Попробуйте найти корень в…
- Проверьте себя: Разуть Заснуть…
- Скачать
- Похожие презентации
Вы можете ознакомиться и скачать
Порядок разбора слова по составу Выделить окончание. Выделить основу. Найти корень. Выделить приставки и суффиксы..
Презентация содержит 4 слайдов.
Презентации для любого
класса можно скачать бесплатно. Если материал и наш сайт презентаций Вам
понравились
–
поделитесь им с друзьями с помощью социальных кнопок и добавьте в закладки в
своем
браузере.
Слайд 1
Описание слайда:
Порядок разбора слова по составу Выделить окончание. Выделить основу. Найти корень. Выделить приставки и суффиксы.
Слайд 2
Описание слайда:
Разбор по составу некоторых частей речи:
Слайд 3
Описание слайда:
Попробуйте найти корень в словах: Разуть Заснуть Воткнутый Начнут Ушла Переел Сожжение Навсегда Вынуть Доить Драчливый
Слайд 4
Описание слайда:
Проверьте себя: Разуть Заснуть Воткнутый Начнут Ушла Переел Сожжение Навсегда Вынуть Доить Драчливый
Теги
Порядок разбора слова по составу Выделить окончание.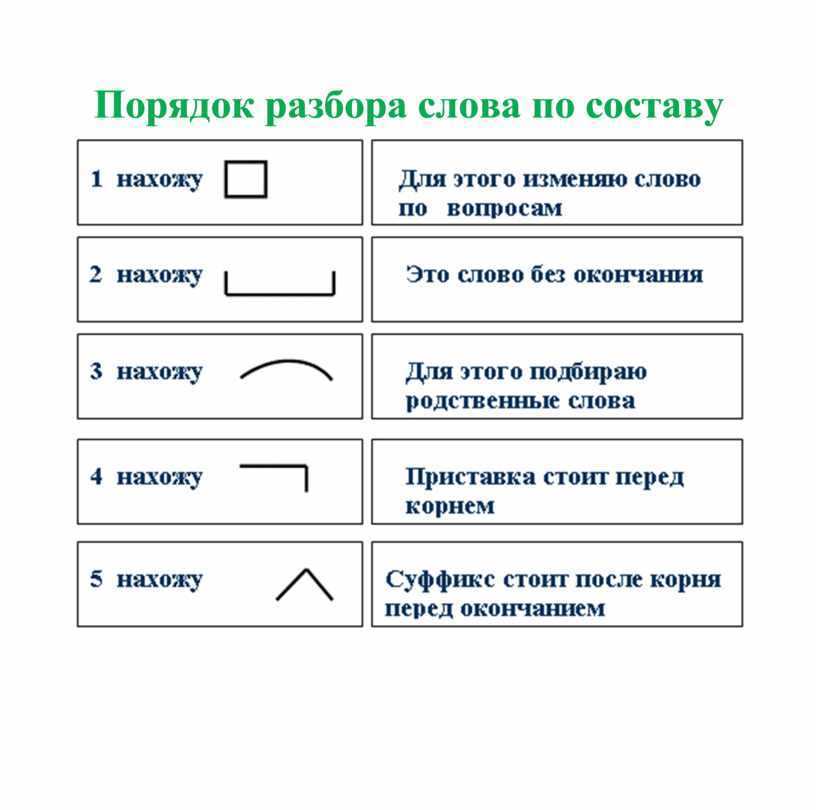
Похожие презентации
Частые ошибки мастера при кератиновом выпрямлении волос
Дорогие мастера восстановления волос, начинающие и уже практикующие! В интернете сегодня огромная куча информации о кератиновом выпрямлении – плюсы/минусы, польза/вред, мифы о кератине, отзывы клиентов, однако очень мало информации реально полезной как мастеру, так и клиенту. Зачастую мастера учатся на бесплатных семинарах, при условии приобретения продукции, и начинают работу на свой собственный страх и риск, на одном составе для всех, с кучей ошибок, разобраться в которых самому очень сложно, ведь опыт и уверенность в руках приходит со временем.
Поэтому хотим разобрать такую тему, как «Частые ошибки мастера при кератиновом выпрямлении волос». Насколько это конечно возможно уложиться в короткой статье, попробую максимально кратко и информативно разобрать наиболее частые ошибки мастеров с самого начала рабочего процесса и до самого конца.
1. Самая серьезная ошибка в работе мастера — это неправильно произведенная диагностика волос клиента. Ведь исходя из базы мы выстраиваем нашу дальнейшую работу. В процессе диагностики вы должны определить степень повреждения волоса от 1 до 5 максимально исходя из всех показателей:
- степень повреждения дисульфидной структуры волоса (тест на эластичность) – насколько волос тянется в мокром виде;
- пористость волоса – насколько волос впитывает и теряет влагу;
- цвет – окрашен волос, осветлен или натуральный;
- этническая принадлежность волоса (да, для новичков это может показать удивительно, но составы для процедур кератинового выпрямления так же подбираются и под этно направление. Для вас ведь не секрет, что азиатские, славянские и афро волосы отличаются по структуре и, соответственно требуют разного подхода).
Правильно произведенная диагностика самый первый шаг к успеху.
2. Необходимо учесть все параметры диагностики! А также пожелания клиента и уже исходя из полученной в процессе диагностики информации вы сможете правильно выбрать состав, который вы будете применять в данном случае. Неверный выбор состава это уже пол пути к недовольному клиенту, не получению желаемого эффекта, короткому периоду носки процедуры и, к сожалению, сожженным волосам. Ведь абсолютно логично, что состав для афро-завитка не подходит для блонда. Включаем голову и думаем. Все ли клиенты хотят кератин? Кому-то достаточно ботокса для питания, кому-то необходима нанопластика, а волосы с высокой степенью повреждения могут не выдержать классический состав.
Необходимо учесть все параметры диагностики! А также пожелания клиента и уже исходя из полученной в процессе диагностики информации вы сможете правильно выбрать состав, который вы будете применять в данном случае. Неверный выбор состава это уже пол пути к недовольному клиенту, не получению желаемого эффекта, короткому периоду носки процедуры и, к сожалению, сожженным волосам. Ведь абсолютно логично, что состав для афро-завитка не подходит для блонда. Включаем голову и думаем. Все ли клиенты хотят кератин? Кому-то достаточно ботокса для питания, кому-то необходима нанопластика, а волосы с высокой степенью повреждения могут не выдержать классический состав.
3. Ошибки при нанесении состава достаточно часто совершают начинающие мастера и мастера самоучки. Не пренебрегайте советом – хорошее обучение вам в помощь. Правильно, качественно, без излишков и самое главное без пробелов нанести состав тоже важно уметь. Внимательно прорабатывайте каждую прядь, следите за тем, чтобы состав был нанесен равномерно, если состав нанесен в недостаточном количестве, то необработанные пряди пострадают от высоких температур при выпаривании состава. Также тщательно прочесывайте пряди для избежания излишков, так как это усложнит выпаривание и приведет к раздражению слизистых глаз и носа и дискомфорту во время работы. Секретный совет новичкам – если после полного нанесения состава вы обратили внимание, что кончики волос, в отличии от основной длины, уже высохли – стоит дополнительно нанести состав на кончики, так как они нуждаются в большем внимании и качественной проработке.
Также тщательно прочесывайте пряди для избежания излишков, так как это усложнит выпаривание и приведет к раздражению слизистых глаз и носа и дискомфорту во время работы. Секретный совет новичкам – если после полного нанесения состава вы обратили внимание, что кончики волос, в отличии от основной длины, уже высохли – стоит дополнительно нанести состав на кончики, так как они нуждаются в большем внимании и качественной проработке.
4. Время выдержки состава является немаловажным фактором для качественного выполнения процедуры. Предположим производитель пишет в инструкции, что время выдержки состава 20 минут. Так ли это будет верно? Если несколько моментов, учитывая которые мастеру просто необходимо увеличить или уменьшить время выдержки состава. Если в помещении холодно, если у клиента очень тугой завиток, если волос жесткий как леска – время выдержки состава стоит увеличить на 10 минут. Если же волос сильно поврежден, очень тонкий или в случае если каратиновое выпрямление выполняется на прямой волос для улучшения качества, уменьшения пушистости волос, то время выдержки можно уменьшить на 10 минут.
5. Сушка волос после выдержки состава
неотъемлемая и очень важная часть процедуры. Что бы вам не говорили на обучении – не принципиально каким воздухом вы будете сушить волосы горячим или холодным, от этого зависит только интенсивность запаха состава. А вот что принципиально – это на 100% высушить волос без использования расчески. Если волос не досушен и в нем осталась влага — это отразиться на сроке носки и качестве результата.6. Использование некачественного инструмента усложнит вашу работу и сведет к минимуму качество и стойкость результата. Для кератинового выпрямления нельзя использовать обычный утюжок для домашнего использования! Качественный утюжок — это скорость, качество и удобство в работе для мастера, шикарный результат и длительная носка для клиента. В арсенале мастера должно быть два утюжка – с широкими и с узкими пластинами. Широкие пластины позволяют быстро прорабатывать длинные волосы, а узкие пластины помогут идеально сформировать челку и короткие волосы.
7. В инструкции к составу написано «Выставить температуру утюжка 230 градусов и протянуть каждую прядь 10 раз». Как так? У нас что, все волосы одинаковые? Верный выбор температурного режима и количества протяжек – это самая важная часть нашей процедуры! Стоит ошибиться, и клиент уйдет от вас с прямым, но сухим волосом и результатом на 2 недели. Прежде всего температуру мы будем выставлять, ориентируясь на качество волос клиента и термозащиту состава. Чем выше качество волос и термозащита, тем выше температуру мы можем выставить. Но начинать всегда стоит с наименьшей температуры, — ставим 210 и смотрим как реагирует прядь, если все идет по плану, то оставляем температуру, если прядь долго остается жирной, то увеличиваем температуру шагом 5 градусов и останавливаемся на оптимальной температуре.
8. Смывать или не смывать состав? Хороший вопрос. Уверены ли вы в том, что у клиента не случится индивидуальной аллергической реакции на состав? Уверены ли вы, что клиент правильно смоет состав сам в домашних условиях с соблюдением всех правил? Ответ очевиден. Если вы хотите убедиться в качестве произведенной процедуры и увидеть крутой результат, то смывайте состав сами! Волос необходимо тщательно промыть большим количеством теплой воды, нанести ухаживающую маску на 10 минут для восстановления уровня РН.
9. Сушке волос отдают несправедливо одно из последних мест в ошибках мастеров. Неправильная сушка волос после процедуры кератина не позволит вам увидеть и оценить качество выполненной вами работы и показать клиенту какой прекрасный результат получился. Сушите волосы направив поток воздуха вниз по направлению роста волос без расчески, мягко прочесывая волосы пальцами и на комфортной для клиента температуре и скорости. По мере сушки клиент будет видеть, как его волосы преображаются, становятся гладкими, блестящими и как они здорово рассыпаются у вас в руках. Правильная сушка позволит вам и клиенту увидеть максимальный результат от процедуры и даст возможность сделать крутые фотки и видео.
Сушите волосы направив поток воздуха вниз по направлению роста волос без расчески, мягко прочесывая волосы пальцами и на комфортной для клиента температуре и скорости. По мере сушки клиент будет видеть, как его волосы преображаются, становятся гладкими, блестящими и как они здорово рассыпаются у вас в руках. Правильная сушка позволит вам и клиенту увидеть максимальный результат от процедуры и даст возможность сделать крутые фотки и видео.
10. Неправильно подобранный домашний уход максимально быстро может свести на нет все ваши старания. Как бы круто вы не отработали, если клиент моет голову дешевеньким составом из массмаркета, не использует бальзамы, маски, дополнительный уход для кончиков и неправильно сушит волосы, то результат процедуры уйдет в несколько раз быстрее. Не жалейте своего времени и сил и посоветуйте клиенту хороший комплексный уход для поддержания эффекта после процедуры кератинового выпрямления. Хороший уход — это не обязательно бессульфатный шампунь и точка. Существуют шикарные профессиональные шампуни с пониженным содержанием сульфатов, с содержание допустимых сульфатных соединений, с низким уровнем PH разработанные специально для химически обработанных, выпрямленных и поврежденных волос. Научите клиента как правильно ухаживать за волосами и тем самым вы окажете ему полный сервис.
Существуют шикарные профессиональные шампуни с пониженным содержанием сульфатов, с содержание допустимых сульфатных соединений, с низким уровнем PH разработанные специально для химически обработанных, выпрямленных и поврежденных волос. Научите клиента как правильно ухаживать за волосами и тем самым вы окажете ему полный сервис.
Подведем итоги? Перестаньте думать о том, как больше заработать и думайте о том, как максимально удовлетворить ожидания клиента от процедуры кератинового выпрямления волос. Подумайте над своими ошибками и проработайте каждую из них. Используйте качественные материалы, заботьтесь о клиенте и будет вам счастье!
Читайте также:
- Нужны ли волосам процедуры по восстановлению?
- Как общаться с трудными клиентами
- Гидрализация волос — Vitaker SOS Hydramax Therapy
Научная статья или глава книги | Регуляция состава и разборки фокальных комплексов с помощью кальцийзависимой протеазы кальпаин | ID: ff365f400
Общественный Депонировано
Аналитика
У вас нет доступа к существующим коллекциям. Вы можете создать новую коллекцию.
Вы можете создать новую коллекцию.
Скачать PDF
Запросить версию для программы чтения с экрана
MLA
Регуляция состава и разборки фокального комплекса с помощью кальций-зависимой протеазы Calpain. 2002. https://doi.org/10.17615/20wg-rr49APA
(2002). Регуляция состава очаговых комплексов и их разборка кальций-зависимой протеазой кальпаином. https://doi.org/10.17615/20wg-rr49Чикаго
2002. Регуляция состава и разборки фокального комплекса с помощью кальций-зависимой протеазы Calpain. https://doi.org/10.17615/20wg-rr49- EndNote
- Аннотация
- Миграция клеток требует регулируемого и динамичного оборота адгезивных комплексов. Ранее мы показали, что кальций-зависимая протеаза, кальпаин, регулирует организацию адгезивных комплексов и отделение клеток во время миграции клеток.
 В настоящее время предоставлены доказательства того, что ингибирование кальпаина за счет сверхэкспрессии эндогенного ингибитора кальпаина, кальпастатина и фармакологических ингибиторов приводит к ингибированию разборки адгезивного комплекса со стабилизацией GFP-винкулина и GFP/RFP-зиксина на периферии клетки. Кальпаин также был необходим для опосредованного микротрубочками оборота участков адгезивных комплексов после вымывания нокодазола, что указывает на то, что кальпаин может опосредовать фокальную разборку комплекса ниже по течению от микротрубочек. Используя двойную визуализацию RFP-зиксин и GFP-альфа-актинин, мы наблюдали временную и пространственную связь между локализацией альфа-актинина в фокальных контактах и последующей разборкой или транслокацией комплексов, содержащих RFP-зиксин, в области ретракции клеток. Ингибирование кальпаина нарушало локализацию альфа-актинина в фокальных контактах, содержащих зиксин, и разборку фокального комплекса или транслокацию в центр клетки. Кроме того, нарушение локализации альфа-актинина в фокальных комплексах посредством экспрессии палочковидного домена альфа-актинина, но не головного домена, приводило к ингибированию разборки фокальной адгезии, аналогичному ингибированию кальпаином. Наши исследования предполагают новый механизм действия, посредством которого кальпаин может модулировать локализацию альфа-актинина в фокальных комплексах и их последующую разборку или транслокацию.
В настоящее время предоставлены доказательства того, что ингибирование кальпаина за счет сверхэкспрессии эндогенного ингибитора кальпаина, кальпастатина и фармакологических ингибиторов приводит к ингибированию разборки адгезивного комплекса со стабилизацией GFP-винкулина и GFP/RFP-зиксина на периферии клетки. Кальпаин также был необходим для опосредованного микротрубочками оборота участков адгезивных комплексов после вымывания нокодазола, что указывает на то, что кальпаин может опосредовать фокальную разборку комплекса ниже по течению от микротрубочек. Используя двойную визуализацию RFP-зиксин и GFP-альфа-актинин, мы наблюдали временную и пространственную связь между локализацией альфа-актинина в фокальных контактах и последующей разборкой или транслокацией комплексов, содержащих RFP-зиксин, в области ретракции клеток. Ингибирование кальпаина нарушало локализацию альфа-актинина в фокальных контактах, содержащих зиксин, и разборку фокального комплекса или транслокацию в центр клетки. Кроме того, нарушение локализации альфа-актинина в фокальных комплексах посредством экспрессии палочковидного домена альфа-актинина, но не головного домена, приводило к ингибированию разборки фокальной адгезии, аналогичному ингибированию кальпаином. Наши исследования предполагают новый механизм действия, посредством которого кальпаин может модулировать локализацию альфа-актинина в фокальных комплексах и их последующую разборку или транслокацию.
- Миграция клеток требует регулируемого и динамичного оборота адгезивных комплексов. Ранее мы показали, что кальций-зависимая протеаза, кальпаин, регулирует организацию адгезивных комплексов и отделение клеток во время миграции клеток.
- Дата публикации
- 2002
- Ключевое слово
- Fibroblasts
- Animals
- Cricetinae
- CHO Cells
- Focal Adhesions
- Glycoproteins
- Calcium-Binding Proteins
- Cell Adhesion
- Cell Movement
- Cells, Cultured
- Actinin
- Cysteine Proteinase Inhibitors
- Цитоскелетные белки
- Кальций
- Калпейн
- ДОИ
- https://doi.org/10.17615/20wg-rr49
- Идентификатор
- Идентификатор Onescience: d5165be2f9b9af227368ff6b068a266d8e4559d0
- Тип ресурса
- Артикул
- Заявление о правах
- В авторском праве
- Название журнала
- Журнал клеточных наук
- Том журнала
- 115
- Выпуск журнала
- Часть 17
- Начало страницы
- 3415
- Конец страницы
- 34125
- Язык
- Английский
- ISSN
- 1477-9137
- 0021-9533
 В настоящее время предоставлены доказательства того, что ингибирование кальпаина за счет сверхэкспрессии эндогенного ингибитора кальпаина, кальпастатина и фармакологических ингибиторов приводит к ингибированию разборки адгезивного комплекса со стабилизацией GFP-винкулина и GFP/RFP-зиксина на периферии клетки. Кальпаин также был необходим для опосредованного микротрубочками оборота участков адгезивных комплексов после вымывания нокодазола, что указывает на то, что кальпаин может опосредовать фокальную разборку комплекса ниже по течению от микротрубочек. Используя двойную визуализацию RFP-зиксин и GFP-альфа-актинин, мы наблюдали временную и пространственную связь между локализацией альфа-актинина в фокальных контактах и последующей разборкой или транслокацией комплексов, содержащих RFP-зиксин, в области ретракции клеток. Ингибирование кальпаина нарушало локализацию альфа-актинина в фокальных контактах, содержащих зиксин, и разборку фокального комплекса или транслокацию в центр клетки. Кроме того, нарушение локализации альфа-актинина в фокальных комплексах посредством экспрессии палочковидного домена альфа-актинина, но не головного домена, приводило к ингибированию разборки фокальной адгезии, аналогичному ингибированию кальпаином.
В настоящее время предоставлены доказательства того, что ингибирование кальпаина за счет сверхэкспрессии эндогенного ингибитора кальпаина, кальпастатина и фармакологических ингибиторов приводит к ингибированию разборки адгезивного комплекса со стабилизацией GFP-винкулина и GFP/RFP-зиксина на периферии клетки. Кальпаин также был необходим для опосредованного микротрубочками оборота участков адгезивных комплексов после вымывания нокодазола, что указывает на то, что кальпаин может опосредовать фокальную разборку комплекса ниже по течению от микротрубочек. Используя двойную визуализацию RFP-зиксин и GFP-альфа-актинин, мы наблюдали временную и пространственную связь между локализацией альфа-актинина в фокальных контактах и последующей разборкой или транслокацией комплексов, содержащих RFP-зиксин, в области ретракции клеток. Ингибирование кальпаина нарушало локализацию альфа-актинина в фокальных контактах, содержащих зиксин, и разборку фокального комплекса или транслокацию в центр клетки. Кроме того, нарушение локализации альфа-актинина в фокальных комплексах посредством экспрессии палочковидного домена альфа-актинина, но не головного домена, приводило к ингибированию разборки фокальной адгезии, аналогичному ингибированию кальпаином. Наши исследования предполагают новый механизм действия, посредством которого кальпаин может модулировать локализацию альфа-актинина в фокальных комплексах и их последующую разборку или транслокацию.
Наши исследования предполагают новый механизм действия, посредством которого кальпаин может модулировать локализацию альфа-актинина в фокальных комплексах и их последующую разборку или транслокацию.- Родители:
У этой работы нет родителей.
Новые способы уничтожения старых боеприпасов | Статья
1 / 5 Показать заголовок + Скрыть заголовок – После извлечения красного фосфора он проходит тот же процесс преобразования, что и белый фосфор, в фосфорную кислоту. Затем фосфорная кислота продается в качестве сельскохозяйственного удобрения, а металл отправляется на местную переработку. Нарисовал ее… (Фото предоставлено армией США) ПОСМОТРЕТЬ ОРИГИНАЛ 2 / 5 Показать заголовок + Скрыть заголовок – (Фото предоставлено армией США) ПОСМОТРЕТЬ ОРИГИНАЛ 3 / 5 Показать заголовок + Скрыть заголовок – (Фото предоставлено армией США) ПОСМОТРЕТЬ ОРИГИНАЛ 4 / 5 Показать заголовок + Скрыть заголовок – Ранее не существовало способа демилитаризации гранаты L8A3 безопасным и безвредным для окружающей среды способом. Благодаря этому процессу теперь существует эффективный и рентабельный способ уничтожения этого устаревшего боеприпаса и обеспечения готовности к будущему…
(Фото предоставлено армией США)
ПОСМОТРЕТЬ ОРИГИНАЛ
5 / 5
Показать заголовок +
Скрыть заголовок –
После извлечения красного фосфора он проходит тот же процесс преобразования, что и белый фосфор, в фосфорную кислоту. Затем фосфорная кислота продается в качестве сельскохозяйственного удобрения, а металл отправляется на местную переработку. Нарисовал ее…
(Фото предоставлено армией США)
ПОСМОТРЕТЬ ОРИГИНАЛ
Благодаря этому процессу теперь существует эффективный и рентабельный способ уничтожения этого устаревшего боеприпаса и обеспечения готовности к будущему…
(Фото предоставлено армией США)
ПОСМОТРЕТЬ ОРИГИНАЛ
5 / 5
Показать заголовок +
Скрыть заголовок –
После извлечения красного фосфора он проходит тот же процесс преобразования, что и белый фосфор, в фосфорную кислоту. Затем фосфорная кислота продается в качестве сельскохозяйственного удобрения, а металл отправляется на местную переработку. Нарисовал ее…
(Фото предоставлено армией США)
ПОСМОТРЕТЬ ОРИГИНАЛ Расположенный на территории в 100 квадратных миль в Южной Индиане, Crane Army Ammunition Activity поддерживает высокотехнологичную миссию по уничтожению устаревших боеприпасов для Министерства обороны. На протяжении более двух десятилетий снаряды с белым фосфором успешно демилитаризировались в рамках закрытой системы и превращались в сельскохозяйственные удобрения, что одновременно обеспечивало готовность к хранению все более модернизируемых боеприпасов и предоставляло продукт, поддерживающий американских фермеров.
На протяжении более двух десятилетий снаряды с белым фосфором успешно демилитаризировались в рамках закрытой системы и превращались в сельскохозяйственные удобрения, что одновременно обеспечивало готовность к хранению все более модернизируемых боеприпасов и предоставляло продукт, поддерживающий американских фермеров.
Препятствие возникло во всем остальном гладкой операции, когда необходимость демилитаризации снарядов с красным фосфором стала настоятельной необходимостью для обеспечения будущей готовности для Joint Warfighter. Из-за небольших молекулярных различий, которые вызывали разные точки возгорания, инженерам Crane Army пришлось разработать процесс, который обеспечил бы безопасное извлечение красного фосфора, чтобы миссия могла продолжаться в течение десятилетий.
История
В 1989 году миссия по демилитаризации белого фосфора в закрытой системе и превращению его в фосфорную кислоту была поставлена перед Crane Army Ammunition Activity. С тех пор установка по конверсии белого фосфора в фосфорную кислоту считается успешной миссией. Он не только демилитаризирует устаревшие боеприпасы безопасным и экологически чистым способом, но и производит два материала, приносящих доход: сельскохозяйственные удобрения и металл для переработки.
Он не только демилитаризирует устаревшие боеприпасы безопасным и экологически чистым способом, но и производит два материала, приносящих доход: сельскохозяйственные удобрения и металл для переработки.
За один день предприятие производит 48 000 фунтов фосфорной кислоты. Для этого в боеприпасе, содержащем белый фосфор, пробивается отверстие, и он вставляется в модифицированную печь с вращающейся печью через систему гравитационной подачи. Два воздуходувки мощностью 75 л.с. используются для поддержания отрицательного давления на материал, который втягивает нагретые пары через систему гидратации потока/противотока для получения концентрации фосфорной кислоты. Дизайн объекта, отвечающий экологическим стандартам с прибылью, сделал его флагманом в поиске процессов демилитаризации закрытых систем.
Около двадцати лет спустя возникла новая проблема. Растет запас различных боеприпасов, содержащих красный фосфор, которые занимают квадратные метры на складах, которые можно использовать для размещения более современных боеприпасов. В то время не существовало метода или процесса для безопасной утилизации красного фосфора путем открытого сжигания или открытой детонации. Состав действует аналогично белому фосфору, однако разница в двух молекулах означает, что необходимо будет создать новый процесс.
В то время не существовало метода или процесса для безопасной утилизации красного фосфора путем открытого сжигания или открытой детонации. Состав действует аналогично белому фосфору, однако разница в двух молекулах означает, что необходимо будет создать новый процесс.
«Раньше у нас не было возможности избавиться от красного фосфора, — сказал руководитель программы демилитаризации CAAA Пол Олсведе. «Мы не могли открыто сжечь его из-за негативного воздействия на окружающую среду, поэтому были предприняты шаги по разработке, чтобы устранить дефицит для Министерства обороны. появился завод по переработке фосфорной кислоты».
Партнерство
После того, как потребность была установлена, SciTech Services выиграла контракт с Консорциумом технологии артиллерийского вооружения Министерства обороны на разработку экономичного и экологически чистого процесса преобразования красного фосфора, содержащегося в дымовых шашках, в фосфорную кислоту.
В частности, они хотели использовать существующий потенциал демилитаризации, завод по конверсии белого фосфора в фосфорную кислоту в Crane Army. Затем SciTech Services заключила субподряд с Gradient Technology для проектирования, изготовления и интеграции прототипа машины, которая могла разбирать дымовые шашки и удалять материал. С 1999 года Gradient Technology и Crane Army совместно работали над несколькими различными проектами по демилитаризации.
Затем SciTech Services заключила субподряд с Gradient Technology для проектирования, изготовления и интеграции прототипа машины, которая могла разбирать дымовые шашки и удалять материал. С 1999 года Gradient Technology и Crane Army совместно работали над несколькими различными проектами по демилитаризации.
«Правительству требовалось решение по демилитаризации, которое не только обеспечивало бы метод демилитаризации самой гранаты с красным фосфором, но также высвобождало бы материал и упаковывало его таким образом, чтобы его можно было подавать в завод CAAA по превращению белого фосфора в фосфорную кислоту. объекта», — сказал руководитель программы Gradient Technology Джо Кэрролл. «Вся инициатива была основана на желании иметь экологически безопасный процесс».
В отличие от боеприпасов с белым фосфором, которые можно разрезать или проделать отверстия в снаряде, чтобы немедленно начать процесс преобразования, у красного фосфора чрезвычайно низкая температура воспламенения. Это означало, что процесс извлечения красного фосфора из некоторых боеприпасов, содержащих красный фосфор, будет намного сложнее, чем из боеприпасов, содержащих белый фосфор. Компания Gradient Technology смогла разработать процесс, который безопасно удаляет красный фосфор из снаряда с наименьшей вероятностью воспламенения.
Это означало, что процесс извлечения красного фосфора из некоторых боеприпасов, содержащих красный фосфор, будет намного сложнее, чем из боеприпасов, содержащих белый фосфор. Компания Gradient Technology смогла разработать процесс, который безопасно удаляет красный фосфор из снаряда с наименьшей вероятностью воспламенения.
В прошлом году были проведены ограниченные испытания, доказавшие жизнеспособность системы, и был начат процесс демилитаризации дымовых гранат L8A3.
Процесс
Дымовая граната L8A3 — это обскурант, который при использовании создает дымовую завесу для укрытия солдата. Содержит красный фосфор. Предметы, содержащие красный фосфор, требуют наличия протоколов для обеспечения безопасного хранения боеприпасов и обращения с ними из-за опасных побочных продуктов, которые может образовывать красный фосфор.
«Красный фосфор выделяет газы из фосфина, а это означает, что когда состав находится на хранении, он будет выделять газ», — сказал Олсведе. «При хранении состава в баллончиках с боеприпасами имеются газопоглощающие модули, которые поглощают фосфин, который не имеет запаха, легко воспламеняется и очень опасен. поглощаться газопоглощающими модулями».
В то время как хранение является вариантом для гранаты, концентрированное количество фосфина может быть очень опасным для тех, кто имеет дело с ним в непосредственной близости. Он легко воспламеняется и токсичен, поэтому очень важно иметь способ своевременной утилизации, чтобы избежать хранения.
Существует несколько этапов демилитаризации дымовой гранаты L8A3. Оператор помещает гранату в модульный блок, созданный Gradient Technology, где цилиндрический резиновый элемент, содержащий состав, отделен от энергоносителя. В этот момент энергетик вывозится оператором из блока и отвозится на демилитаризацию в другое время в другом месте другим способом.
Затем кусок резины, содержащий композицию, просверливают, чтобы высверлить крошки красного фосфора. Крошки падают в металлическую канистру внизу, а теперь пустой кусок резины вычищается и становится отходами, предназначенными для захоронения отходов.
«В этот момент красный фосфор наиболее уязвим для тления или возгорания», — сказал Алсведе. «Когда вы работаете с красным фосфором, это похоже на сверление коробки от спичечных головок. Материал по своей природе хочет гореть, и мы пытаемся просверлить его, не горя».
Для защиты операторов в случае тления компания Gradient Technology разработала модульную установку, внутри которой разбирается дымовая шашка и удаляется красный фосфор, с различными уровнями защиты.
«Материал, стальной лист толщиной в четверть дюйма, был выбран для обеспечения защиты от самой большой угрозы воспламенения — разрывной трубы», — сказал Кэрролл. «Стены модульного блока спроектированы так, чтобы выдерживать детонацию разрывной трубы и должным образом снижать связанные с этим опасности, включая осколки, тепловой поток и тепло».
Помимо детонации, еще одной проблемой безопасности на протяжении всего процесса является дегазация фосфина. Чтобы защитить операторов от скопления газа, была установлена система вентиляции, чтобы гарантировать, что оператор не подвергается воздействию опасного газа.
«Система вентиляции, встроенная в машину, правильно выпускает воздух изнутри машины», — сказал Кэрролл. «В логику компьютера встроены предохранительные блокировки, которые даже не позволят машине работать, если вентиляция не будет достаточной».
Кэрролл также упомянул, что в случае воспламенения или тления материала вентиляционная система устройства будет улавливать и отводить дым или газ, образовавшиеся в результате горения, и предотвращать воздействие вредных паров на оператора.
Во время ограниченного тестового запуска были отдельные случаи тления, поэтому инженеры CAAA разработали и провели эксперимент, чтобы определить, что необходимо сделать для снижения риска тления. Чтобы решить эту проблему, в процессе предусмотрены две отдельные точки выдержки, которые позволяют материалу охлаждаться перед его упаковкой на поддон с другими канистрами, готовыми к следующему этапу процесса.
«Чем холоднее становится материал, тем более стабильным он становится», — сказал Олсведе.
Сразу после того, как крошки падают в металлическую канистру, канистра стоит на одном месте примерно восемь минут. После этого его перемещают в другое место для охлаждения. Только после этого подойдет оператор, чтобы переместить канистру на поддон с другими канистрами, готовыми к транспортировке на завод по конверсии белого фосфора в фосфорную кислоту для демилитаризации.
Будущее
После завершения этого процесса красный фосфор обрабатывается так же, как и белый фосфор. Он поступает на тот же конверсионный завод и превращается в фосфорную кислоту, которая в конечном итоге будет использоваться для удобрения.
Во всем есть преимущества этого типа закрытой системы демилитаризации. CAAA не только избавляется от боеприпасов, которые могут стать опасными по мере их износа, но и получает доход, который компенсирует затраты на производство, что в конечном итоге сэкономит деньги налогоплательщиков.
«Доход — это просто вишенка на торте», — сказал Алсведе. «Настоящая задача — сократить запасы небезопасных устаревших боеприпасов».
Gradient Technology также имеет возможность продвигать свою отраслевую миссию.
«Этот тип совместного выполнения программы позволяет отрасли использовать свой опыт в области проектирования, проектирования оборудования и изготовления на заказ для поддержки миссии правительства, высвобождая ресурсы правительства для поддержки основной миссии, которая заключается в поддержке Warfighter», — Кэрролл. сказал.
Расширение возможностей по демилитаризации красного фосфора было непростой задачей, но важной по многим причинам. Эта способность важна не только для уничтожения устаревших боеприпасов таким образом, чтобы не нанести вред окружающей среде, но и теперь является ключевым элементом в обеспечении боевой готовности нашего национального истребителя.
«Возможность перехода от разоружения боеприпасов с использованием белого фосфора к использованию красного фосфора обеспечит безопасный и безвредный для окружающей среды метод утилизации предметов без других средств разоружения», — сказал заместитель командующего CAAA Норман Томас.